Lutein is a lipid-soluble pigment that can be found naturally in algae and plants. There, it is a component of the light-collecting complexes in the chloroplasts.
For example, kale contains a relatively high concentration of up to 0.25mg lutein per g wet weight. For industrial purposes, however, lutein is extracted from the petals of marigold; they contain up to 8.5mg/g wet weight.
In the animal organism, lutein occurs in the egg yolk, in milk, or the macula lutea (“yellow spot”) of the animal/human eye. However, animals and humans cannot synthesize it.
Lutein belongs to the group of carotenoids, which is divided into carotenes and xanthophylls. Lutein, chemically expressed as “3,3’-dihydroxy-α-carotene”, is a xanthophyll always accompanied by its isomer zeaxanthin. It is synthesized out of two α-carotenes through hydroxylation.
Lutein provides benefits for animals and humans
Due to its beneficial characteristics, lutein is an essential ingredient of plants and is used in animal nutrition as well as in human medicine.
Lutein has antioxidant protective properties
Under normal conditions, the cells in the animal and human organism control ROS (reactive oxygen species) levels. Usually, there is a balance between the generation of ROS and their elimination by scavenging systems. However, the high performance levels in modern animal production can easily lead to high ROS levels, translated into oxidative stress and leading to cell damage. Cell damage contributes to the generation of cancer and early aging in humans. In animals, the negative impact of oxidative stress can be responsible for lower performance and inferiority of meat and eggs.
Antioxidants stop ROS by taking up their energy
Through the uptake of energy, molecules can get into an excited state. One example is singlet excited oxygen, a highly reactive form of oxygen able to destroy proteins, lipids, and DNA. Carotenoids can intervene in this process: by exchanging electrons, the singlet excited oxygen gets neutralized, and the carotenoid gets into this excited state with higher energy. Once able to release this energy as heat into the environment, the carotenoid gets back to its normal state and can once again start acting as an antioxidant.
In this way, carotenoids, including lutein, ‘quench’ the energy of excited molecules and prevent the adverse effects of ROS (reactive oxidative substances).
Antioxidant properties profitably used
The antioxidant character of lutein plays an important role in the treatment or prophylaxis of macular degeneration in humans (Landrum & Bone, 2001). There is also evidence that lutein can be used to improve the visual and retinal function in dogs (Wang et al., 2016). In the eye, lutein and zeaxanthin, occurring in the retina and the macula, neutralize free radicals produced due to the ultraviolet light and thereby prevent damage to the macula.
Further possible applications are against cardiovascular diseases (Dwyer et al., 2001) and various types of cancer (e.g., breast cancer, Gong et al., 2018).
Lutein is important in infant nutrition
Lutein and its isomer zeaxanthin are the two primary carotenoids found in human milk (Giordano and Quadro, 2018). Stringham and co-workers (2019) postulate that lutein plays an important role in children’s visual and cognitive development/optimization. They report that a lutein supplementation of the mother can lead to a higher concentration of this substance in the milk and, consequently, in the child’s plasma (Sherry et al., 2014). In dairy cows, an increased level of lutein in the milk can also be observed (Xu et al., 2014), suggesting that lutein could also be essential in calf development.
Lutein stimulates the immune system
Another benefit of lutein is its positive influence on the immune system.
On the one hand, lutein stimulates the production of antibodies. In dogs, Guimarães Alarça et al. (2016) could show an increase of CD4+ and CD8+ T-lymphocyte subtypes. Kim et al. (2000) demonstrated the increase of lymphocytes and cells expressing CD5, CD4, CD8, and major histocompatibility complex class II (MHC II) molecules. Bédécarrats and Leeson (2006) provoked a higher secondary antibody response to infectious bronchitis vaccination in laying hens.
Besides, lutein acts as an anti-inflammatory agent, as shown in vitro by Chao et al. (2015) and in broiler chickens by Moraes and team (2016).
Lutein improves the attractivity of poultry products
In the marketing of poultry products, appearance and color are of central importance for evaluating quality. Egg yolk coloration is to a large extent a matter of regional preferences, however it is clear that an egg with a yolk that does not have the typical color is classified as inferior by the consumer. In areas with traditional corn growing, a white-skinned chicken is not commercially viable. Even when pullets are bought, the shanks and beaks should be yellow.
The use of xanthophylls like lutein and zeaxanthin enables producers to safely control the color of the egg yolk and of the broiler skin. It also leads to a healthy color of the shanks and beaks of the birds.
Lutein in a nutshell
Lutein is a true all-rounder: a substance that delivers benefits across the board. In plants, it helps fruits and petals become attractive for insects and other animals. It positively influences the animal, acting as an antioxidant, promoting infant development, and stimulating the immune system. As a pigment, it makes poultry and poultry products look more attractive to the consumer. Through its presence in eggs and milk, lutein provides clear and clean benefits to both animals and humans.
References
Bédécarrats, G.Y. and S. Leeson. “Dietary lutein influences immune response in laying hens.” J. Appl. Poult. Res. 15 (2006): 183–189.
Chao, Shih-Chun, Tommaso Vagaggini, Chan-Wei Nien, Sheng-Chieh Huang, and Hung-Yu Lin. “Effects of Lutein and Zeaxanthin on LPS-Induced Secretion of IL-8 by Uveal Melanocytes and Relevant Signal Pathways.” Journal of Ophtalmology, vol. 2015 Article ID 152854 (2015): 7 pages. https://doi.org/10.1155/2015/152854
Dwyer, James H., Mohamad Navab, Kathleen M. Dwyer, Kholood Hassan, Ping Sun, Anne Shircore, Susan Hama-Levy, Greg Hough, Xuping Wang, Thomas Drake, C. Noel Bairey Merz, and Alan M. Fogelman. “Oxygenated Carotenoid Lutein and Progression of Early Atherosclerosis.” Circulation (American Heart Association) 103, no. 24 (2001): 2922-2927.
Gong, Xiaoming, Joshua R. Smith, Haley M. Swanson, and Lewis P. Rubin. “Carotenoid Lutein Selectively Inhibits Breast Cancer Cell Growth and Potentiates the Effect of Chemotherapeutic Agents through ROS-Mediated Mechanisms.” Molecules 23 no. 4(2018): 905.
Kim, Hong Wook, Boon Chew, Teri Ann S Wong, Jean Soon Park, Bor-Chun Weng, Katherine M Byrne, Michael G Hayek, and Gregory A. Reinhart. “Dietary lutein stimulates immune response in the canine.” Veterinary Immunology and Immunopathology 74 no. 3-4 (2000): 315-327.
Moraes, M. L., A. M. L. Ribeiro, E. Santin, and K. C. Klasing. “Immunology, health, and disease: effects of conjugated linoleic acid and lutein on the growth performance and immune response of broiler chickens.” Poultry Science 95 (2016): 237–246.
Ochoa Becerra, Mario, Luis Mojica Contrerasa, Ming Hsieh Loa, Juan Mateos Díaz, Gustavo Castillo Herrera. “Lutein as a functional food ingredient: Stability and bioavailability.” Journal of Functional Foods 66 (2020): 103771.
Sherry, Christina L., Jeffery S. Oliver, Lisa M. Renzi, and Barbara J. Marriage. “Lutein supplementation increases breast milk and plasma lutein concentrations in lactating women and infant plasma concentrations but does not affect other carotenoids.” J. Nutr. 144 (2014): 1256–1263.
Stringham, James M., Elizabeth J Johnson, and B Randy Hammond. “Lutein across the lifespan: From childhood cognitive performance to the aging eye and brain.” Curr Dev Nutr 3 (2019): nzz066.
Xu, C.Z., H. F. Wang, J. Y. Yang, J. H. Wang, Z. Y. Duan, C. Wang, J. X. Liu , and Y. Lao. “ Effects of feeding lutein on production performance, antioxidative status, and milk quality of high-yielding dairy cows.” J. Dairy Sci. 97; American Dairy Science Association (2014):7144–7150
Digital EuroTier 2021: Get to know EW Nutrition like never before
Visitors to EW Nutrition’s innovative online booth at this year’s EuroTier (9-12 February) will get exclusive access to our program and product presentations, delivered by our technical experts. Look forward to exploring our newly acquired Feed Quality and Pigment offers, as well as a live stream session on passive immunity in dairy calves.
The EW Nutrition team will share our expertise on how to support gut health, mitigate the impact of mycotoxins and bacterial toxins, and reduce the incidence of diarrhea in young animals. We will also introduce visitors to our recently-acquired new product ranges, including the SANTOQUIN® feed preservative range, the feed mill processing aid SURF●ACE®, and the feed ingredient Agrado® Plus. Our technical experts will be available for audio and video meetings, which can be scheduled via the EuroTier 2021 online platform.
Not to be missed is EW Nutrition’s “spotlight” event on cattle, presented by our technical manager for Farm Solutions, Lea Poppe. Lea will explore the topic of “Passive Immunity in Dairy Calves: Recent Insights and Innovative Solutions” during a public event for all EuroTier visitors. Join us at 9 am, CET, on February 10, for an in-depth presentation and lively discussion.
As the 2021 EuroTier will take place fully online, visitors to the world’s leading trade fair for animal production will be able to enjoy attractive digital networking opportunities. We look forward to meeting you there!
About EW Nutrition
EW Nutrition is a global animal nutrition company that offers integrators, feed producers, and self-mixing farmers comprehensive, customer-focused solutions for feed quality, gut health management, antibiotic reduction, young animal nutrition and more.
EW Nutrition acquires Feed Quality and Pigment business from Novus International
VISBEK (February 1, 2021) – EW Nutrition has today completed the acquisition of the Feed Quality and Pigments business from Novus International, Inc. Under the terms of the agreement, EW Nutrition becomes the owner of world-renowned brands such as Santoquin® feed preservative, SURF●ACE®, a feed mill processing aid, and feed ingredient Agrado®. The acquisition also gives EW Nutrition ownership of a state-of-the-art production facility in Constantí, Spain.
More challenges? More solutions.
“This transaction will bring additional value to our customers, further reinforcing EW Nutrition’s global market position, and increasing its product portfolio and geographical reach,” says Michael Gerrits, Managing Director of EW Nutrition. “The products acquired will further support EW Nutrition’s mission to mitigate the impact of antimicrobial resistance by providing comprehensive animal nutrition solutions.”
Dan Meagher, President and CEO of Novus International, Inc., explained that the sale is part of Novus’s Project Destiny, a multi-year plan to focus the company’s resources on core platforms and emerging technologies, with a focus on gut health.
“We are pleased to have found a committed owner for these platforms so that they may continue bringing value to the industry,” said Meagher. “This event is a significant milestone in our Project Destiny journey. Now that our Feed Quality and Pigments platforms are in good hands with EW Nutrition, we are excited to focus our energies on developing new, innovative technologies into meaningful nutrition solutions for our customers.”
A robust services agreement between the companies is governing critical activities to ensure customers are supported through the transition.
The range of products is immediately available to our customers. You can find out more details on our Products page or by going directly to the relevant web page.
From sub-acute ruminal acidosis to endotoxins: Prevention for lactating cows
by Technical Team, EW Nutrition
Sub-acute acidosis (SARA) is linked to high levels of ruminal LPS. The LPS cause inflammation and contribute to different metabolic conditions and diseases. Various strategies and solutions can be applied to modulate the rumen microbiota and prevent this risk.
In sub-acute rumen acidosis (SARA), the quantity of free lipopolysaccharides (LPS) coming from Gram- bacteria increases considerably. These LPS cross the ruminal wall and intestine, passing into the bloodstream. The negative consequences on the health of the animal are then reflected in decreased productive and reproductive performance.
The LPS are released during the lysis of GRAM- bacteria which die due to the low pH, and these bacteria are mainly responsible for the production of propionic acid for the energy yield that is obtained. It is essential to preserve ruminal balance between Gram+ and Gram- such that there is no excess of LPS.
Nutritional needs of lactating cows with SARA
In the first phase of lactation (from 1 week after calving to 80 – 100 days of lactation), the cow needs a high energy level to meet the large demand for milk production. This energy demand is often not fully satisfied and feed intake falls short. This deficit leads to the need to provide as much energy as possible per feed ration.
Imagine a 650 kg live weight cow, producing about 35 kg of milk per day with a fat percentage of 3.7 and a protein percentage of 3.2. To achieve this production level and fulfill its maintenance requirements, this animal needs a feed intake of 22 kg of dry matter (DM) per day, with an energy level of 21 UFL equal to 36,000 Kcal/day of NE l (Net Energy Lactation)).
To obtain an energy supply of this type, it is necessary to provide rations with a high content of cereals rich in nonstructured carbohydrates (NSC). This will allow the animals to obtain the maximum efficacy in getting the NE I from the metabolizable energy (ME) expressed as kl*.
*kl expresses the effectiveness in passing from EM to EN l net of the heat dissipated by the animal, therefore kl = ENl/EM (Van Es 1978).
Compared to a diet rich in NDF (Neutral Detergent Fiber), this type of diet promotes and stimulates certain strains of bacteria to the detriment of others, shifting the balance towards a greater population of bacteria that produce propionic acid instead those which produce acetic acid. This change also determines a greater share of Gram- compared to Gram+.
What is rumen acidosis?
Rumen acidosis is that “pathology” whereby the volume of SCFA (Short Chain Fatty Acids) produced by the rumen bacteria is greater than the ability of the rumen itself to absorb and neutralize them. Rumen acidosis is mainly caused by the amylolytic and saccharolytic bacteria (Streptococcus bovis; Selenomonas ruminantium, Bacteroides amylophilus, Bacteroides ruminicola and others) responsible for the production of lactic acid. Unlike the other most representative volatile fatty acids (acetic, butyric and propionic), lactic acid has a lower pKa: 7 (3.9 versus 4.7). This means that for the same amount of molecules produced, lactic acid releases a number of ions H+ in the fluid ten times greater than other VFAs, with evident effects on the pH.
Ruminal acidosis can be characterized as acute or subacute. During acute ruminal acidosis, the pH in the rumen drops below 4.8 and remains low for an extended period of time. Acute acidosis leads to complete anorexia, abdominal pain, diarrhea, lethargy, and eventually death. However, the prevalence of acute acidosis in dairy is very low.
Consequences of rumen acidosis
In such situations, a series of negative consequences can be triggered in the lactating cow. Investigations (for instance, using fistulated cows) can reveal, among others, the following alteration in the rumen:
Shift in total microbiome rumen profile (density; diversity; community structure)
Shift in protozoa population (increase in ciliates protozoa after 3 weeks of SARA; increase in the GNB population)
Shift in fungi population (decreasing the fungi population with high fibrolytic enzymes, which are sensitive to low pH)
Rise in LPS rumen concentration (increasing the GNB strain and their lysis)
Influence on the third layer of Stratified Squamous Epithelium (SSE) (desmosomes and tight junctions)
Lower ruminal fiber degradation (reduction in the number of cellulolytic bacteria which are less resistant to acid pH)
Reduction of the total production of fatty acids (propionic, acetic, butyric), therefore less available energy
Lower rumen motility (also as a consequence of the smaller number of protozoa)
The increased acid load damages the ruminal epithelium
Acid accumulation increases the osmotic pressure of the rumen inducing an higher flux of water from the blood circulation into the rumen, causing swelling and rupture of rumen papilla as well as a greater hemoconcentration
The last points are extremely important, as it enables an easier passage of fluids from the blood to the pre-stomachs, greatly influencing the fermentation processes.
Furthermore, with diets low in NDF, the level of chewing and salivation is certainly lower, with a consequent lower level of salivary buffers that enter the rumen and which would maintain an appropriate pH under normal conditions.
Rumen sub-acute and acute acidosis: a fertile ground for LPS
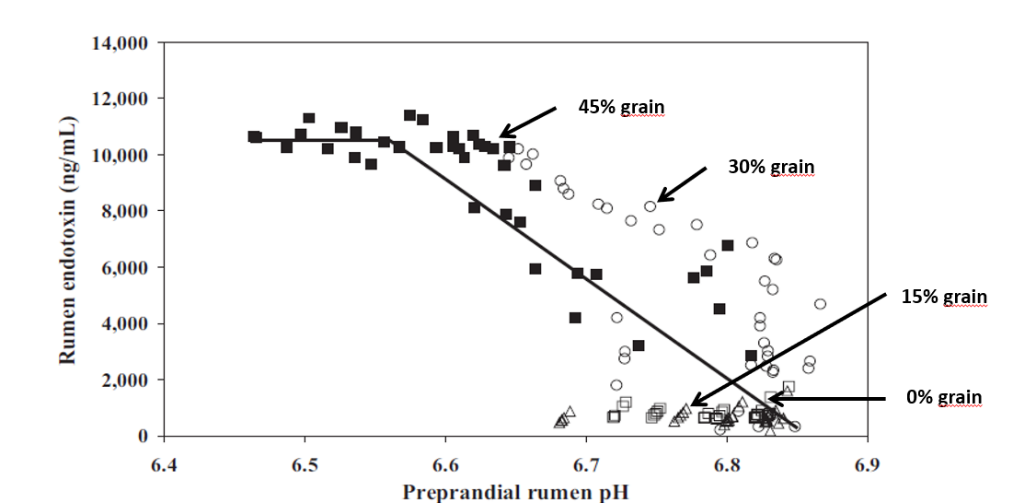
Studies inducing SARA in dairy cows have shown that feeding high levels of grain causes the death and cell lysis of Gram- bacteria, resulting in higher concentration of free LPS in the rumen. In a trial conducted by Ametaj et al., in 2010 (Figure 1), a lower ruminal pH and an increase in the concentration of LPS in the rumen fluid -measured as ng / ml (nanograms / milliliter)-, was the result of increasing of NSC present in the diet (% of grains).
Figure 1. The increase in the level of endotoxins in the rumen is directly correlated with an increase in ration concentrates
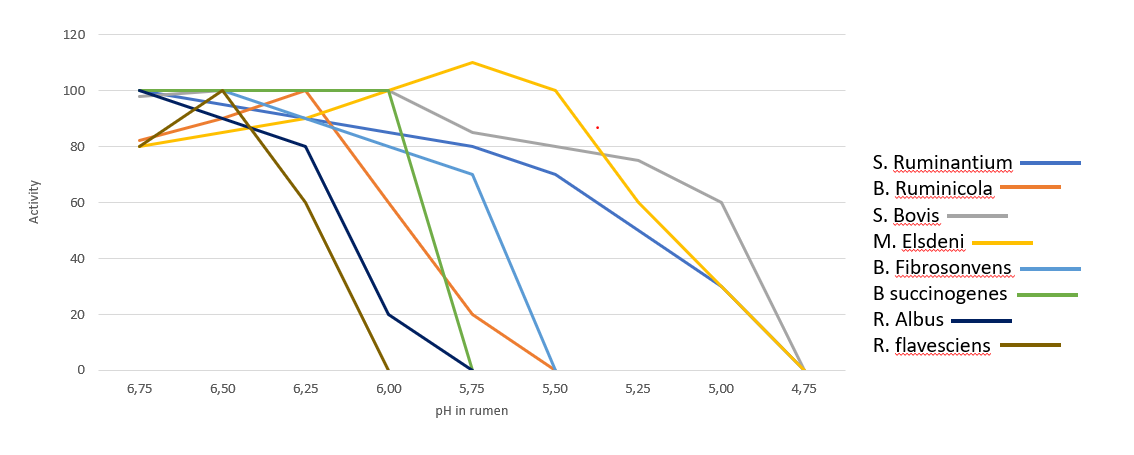
In the rumen, the presence of Gram- is very significant, however the dietary changes towards high energy concentrates, reduce the substates necessary for them to thrive, leading to their lysis and favoring gram-positive bacteria (Gram+). Gram+ also produce bacteriocins against a wide variety of bacteria, including many Gram-. Figure 2 shows the influence of ruminal pH in the population of different bacteria, many of which are are crucial for the production of SCFA and therefore of energy.
Figure 2. Activity of main bacteria in the rumen in function of pH (Daniele Cevolani Edizioni Agricole di New Business Media srl 2020)
It is therefore necessary to pay close attention to the energy level of the ration as an energy input (generally around 1500 – 1700 Kcal/kg of DM intake). At the same time, we need to ensure that the animal does receive and ingest that daily amount of DM. If ingestion is negatively influenced by acidosis (clinical or sub-clinical), this can lead to endotoxemia, with harmful consequences for the animal’s health and production performance.
We can therefore note that the level of LPS (endotoxins) present in the rumen is directly correlated with the pH of the rumen itself and with a symptomatologic picture dating back to SARA. This occurs when the mortality and lysis of Gram- bacteria (GNB) is high and through the consequent imbalance created with diets containing excess fermentable starches, compared to diets with higher fiber content.
In fact, it was shown that the transition from a concentrated fodder ratio of 60:40 to a more stringent ratio of 40:60 caused the level of free LPS in the rumen to go from 410 to 4.310 EU / ml.
Endotoxemia: Pathological consequences in dairy cows
Once the LPS enter the bloodstream, they are transported to the liver (or other organs) for the detoxification. However, sometimes this is not enough to neutralize all the endotoxins present in blood. The remaining excess can cause issues such as the modification of the body’s homeostasis or cause that cascade of inflammatory cytokines responsible for the most common pathologies typical in cows in the first phase of lactation. The most common symptoms are the increase of somatic cells in milk or claws inflammation.
Pro-inflammatory cytokines as TNF, IL6 and IL8 induced by LPS-related inflammation are able to stimulate the production of ACTH (adrenocorticotropic hormone).
ACTH, together with cortisol and the interleukins, inhibit the production of GnRH and LH, with serious effects on milk production. The productivity and the fertility of the animal are thus compromised.
Moreover, prostaglandins are as well stimulated by LPS, and are linked with fever, anorexia and ruminal stasis. This not only limits the amount of energy available for production and maintenance functions, but also induces a higher susceptibility to disease and adds-up to the emergence of other metabolic conditions, such as laminitis and mastitis.
Preventing rumen acidosis
The solution to these massive risks is a prudent and proactive approach by the nutritionist towards all situations that can cause a rapid increase of Gram- in the rumen.It is therefore necessary to avoid cases of clinical and sub-clinical acidosis (SARA) in order to avoid the issues listed above. This would also help avoid stressful conditions for the animal that would lead to decreased performance and health.
To maintain balance and a healthy status of the animal, the use of additives such as phytomolecules and binders is suggested in the first phase of lactation, starting from 15 days before giving birth.
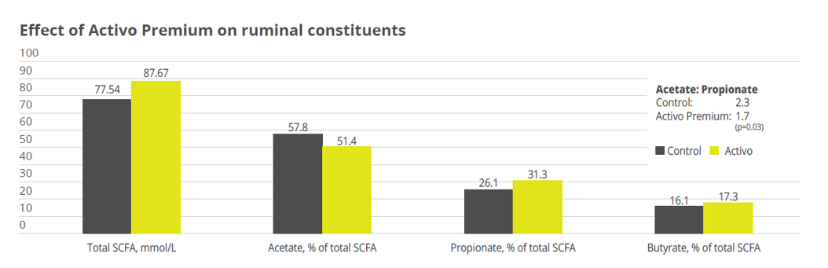
Activo Premium (a mix of phytogenic substances) has given excellent results in decreasing the acetic/propionic acid ratio, while safeguarding the population of Gram+ bacteria. This is in contrast to treatments with ionophores, which, as is well known, interfere with the Gram+ population.
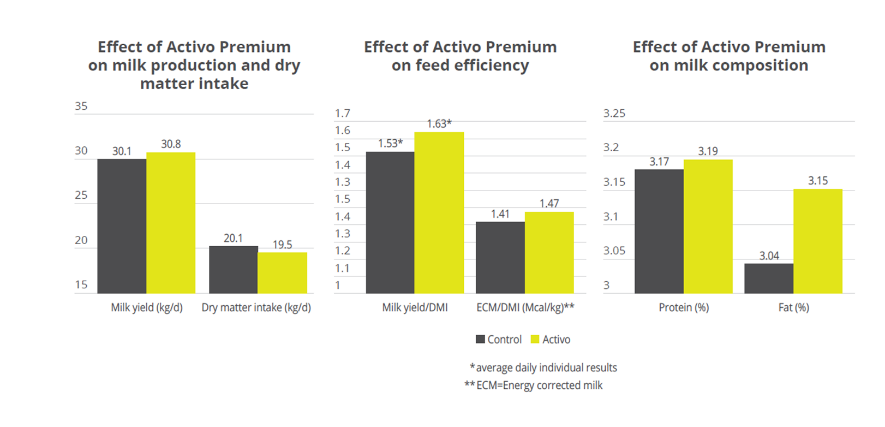
Case study. Acetic acid:propionic acid ratio with Activo Premium
In a study conducted at the the University of Lavras and the Agr. Res. Comp. of Minas Gerais (both Brazil), 30 Holstein cows were allocated to two groups considering parity and milk production. One group was fed the standard feed (control), the other group received standard feed containing 150mg of Activo Premium/kg of dietary dry mass (DM). The following parameters were measured or calculated: intake of DM and milk production, milk ingredients such as fat, protein, lactose every week, body weight and body condition score every two weeks, and ruminal constituents (ph and SCFAs) through oesophaeal samples at day 56.
Activo Premium was able to decrease the ratio between acetic acid and propionic acid, and at the same time maintain the level of Gram+ bacteria in the rumen, thus reducing the risk of endotoxins.The same trial carried out at the University of Lavras demonstrated how the performance of the animals was superior in the group fed with Activo Premium compared to the control group (see below).
Figure 3. Effect of Activo Premium on ruminal constituents
Figure 4. Effect of Activo Premium on animal performance
Solution: Preserve Gram+ bacteria levels while decreasing free LPS
We have therefore seen how important it is to decrease the acetic:propionic ratio in the rumen to obtain a greater share of available energy. However, the level of endotoxins in the rumen must remain low in order to avoid those problems of endotoxemia linked to very specific pathologies typical of “super productive cows”. These pathologies (always linked to inflammatory manifestations) can be prevented by decreasing the level of free LPS in the rumen with a product that can irreversibly bind the LPS and thus make them inactive.
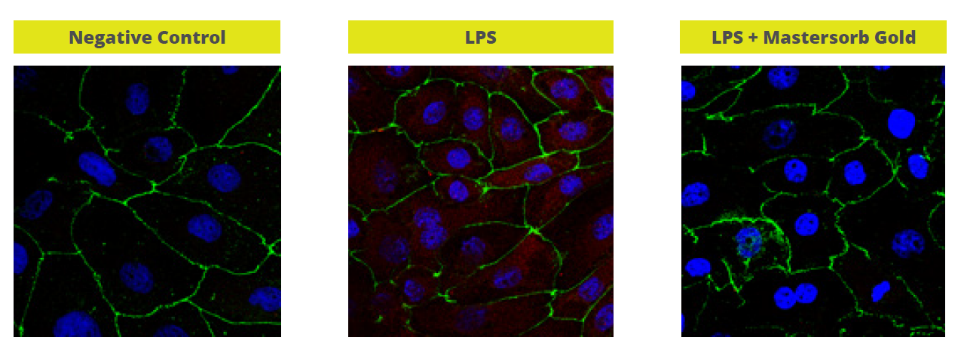
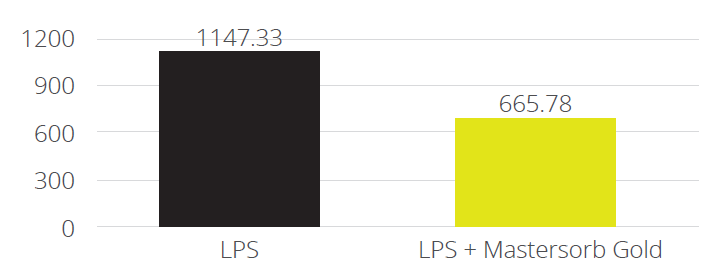
In a trial with porcine intestinal cells (IPEC-J2) challenged by E. coli LPS, a decrease in the intensity of inflammation was observed when Mastersorb Gold was added. This decrease could be shown through a lower amount of phosphorylated NF-kB in an immunofluorescence trial, as well as through the reduced production of Interleukin (IL)-8 in the cells measured by ELISA.
The fact that pig intestine tissue was used does not affect the adsorption concept. In this case, these intestinal cells are only a vehicle to demonstrate that in an aqueous solution containing 50 ŋg of LPS / ml and in the same solution with the addition of Mastersorb Gold, the level of LPS actually active is decreased, as a part of the LPS was tied up by Mastersorb. The solution with a lower level of LPS gave minor “inflammatory” reactions to intestinal cells, and this can be statistically reported in dairy cows.
Figure 5. Immunofluorescence in PEG-J2: Challenge with LPS without (in the middle) and with Mastersorb Gold (right)
Figure 6. IL-8 AP secretion after incubation with LPS 0111:B4 for 24h without and with Mastersorb Gold
Conclusions
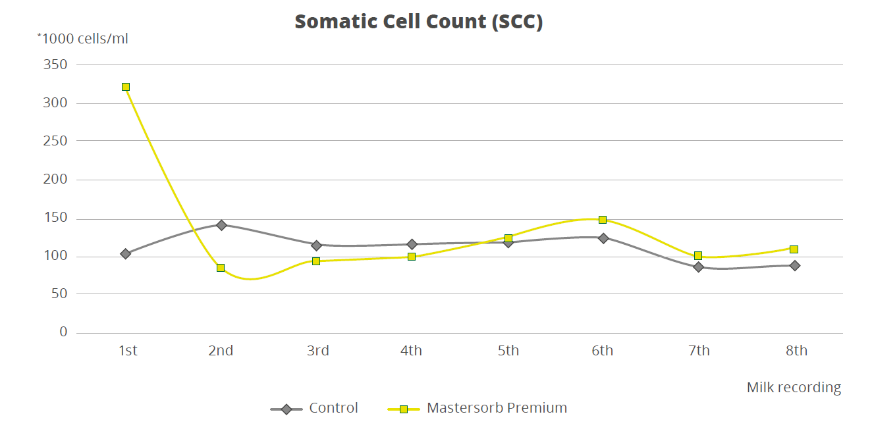
To demonstrate how the decrease in the level of LPS in the rumen is directly correlated with inflammatory states in general, a trial with a total of 60 dairy cows shows that the inclusion of 25g of Mastersorb Premium/animal/day increases milk yield and improves milk quality by decreasing somatic cell count. Adsorbing substances contained in Mastersorb Premium tie up the LPS produced in the rumen in different cow lactation phases.
Normally, the rise in the level of somatic cells in milk depends on etiological agents such as Streptococcus spp, Staphylococcus spp, mycoplasma and more. LPS stress is not the sole agent responsible for raising somatic cell counts, but also other factors among which:
Lactation stage and age of the animal
Season of the year (in summer the problem is increased)
Milking plant (proper maintenance)
General management and nutrition
However, by reducing the level of LPS, Mastersorb provides an important aid to decrease somatic cell count.
Figure 7. Effect of Mastersorb Premium on somatic cell count
Prevent escalation with rumen balance
In the end, ruminant producers are, like all livestock operations, interested in producing healthy animals that can easily cope with various stressors. Ensuring a proper diet, adjusted to the energy requirements of various production stages, is a first step. Providing the animal with the ingredients that modulate the microbiota and reduce the negative impact of stress in the rumen is the next essential step in efficient production.
EW Nutrition launches new xylanase enzyme in Malaysia
Singapore – December 28, 2020 – In its continued efforts to reduce antibiotic use in the region while also improving farm profitability, EW Nutrition launched Axxess® XY in Malaysia on December 17, 2020.
Axxess® XY is a next-generation intrinsically thermostable xylanase, with unparalleled stability under high temperatures and especially longer conditioning time. With its ability to break down both the soluble and insoluble fiber fraction from feed ingredients, the enzyme offers high flexibility in feed formulation. The newly launched product thus gives feed producers peace of mind when working with various feed processing conditions, as well as significant feed cost savings.
The launch took place in the context of a customer online event titled “Revolutions in Poultry Nutrition: The Future of Enzymes” for customers in Malaysia. During the information-laden event, Dr. Howard Simmins, an accomplished global communicator and independent key opinion leader, highlighted the need for improved functionality from xylanases. With his vast experience on feed additive development for animal nutrition and health, including cutting edge biotech research into enzymes, probiotics, prebiotics and novel additives, Dr. Simmins also discussed ways to formulate feed accurately to optimize xylanase value, and optimize nutrition with enzyme products.
During the event, other presentations were delivered by Daniel Tepe, Managing Director, Dr. Andreas Michels, Head of Biotechnology, and Dr. Ajay Awati, Global Category Manager Gut Health and Nutrition, EW Nutrition. The webinar was hosted by Jurek Grapentin, Regional Director, EW Nutrition South East Asia/Pacific.
The webinar was rounded off with an interactive Question and Answer session, where the audience had the opportunity to have their questions answered live by the panel of speakers.
“This revolutionary enzyme is a testament of our dedication to provide a holistic suite of animal nutrition solutions to our valued customers in this region. Axxess® XY will provide a competitive edge with regard to feed formulation, which translates to cost savings to our customer. With the expertise of our R&D researchers, we are constantly striving to bring more innovative solutions to meet the challenging needs of this industry,” said Jurek Grapentin.
Strong demand by consumers; restaurant chains and wholesalers for antibiotic-free (ABF) meat; the threat of antimicrobial resistance; and stringent regulations on the use of antibiotics – there are many good reasons for poultry producers to strive for antibiotic-free production systems. Crucially, to successfully produce poultry meat without antibiotics requires a paradigm shift that starts right at the parent stock level, with the antibiotic-free production of hatching eggs.
Broiler breeders’ gut health is linked to progeny’s performance
Broiler breeders’ performance is measured in terms of how many saleable day old chicks (DOCs) per hen they produce. However, within a sustainable ABF production system (also known as No Antibiotics Ever or NAE), this parameter is not seen in isolation. Breeder hens’ nutritional and health status not only affect the number of DOCs they can produce, but also the transfer of nutrients, antibodies, microbiota and even contaminants, e.g. mycotoxins, to the egg – and therefore, their progeny’s long-term health and performance.
This starts with egg formation, which requires several metabolic processes in the hen to function perfectly. If the hen’s intestinal integrity is compromised, for example due to mycotoxins, she will absorb fewer nutrients, which in turn affects egg formation. Mycotoxicosis has particularly insidious effects for egg formation as it can damage the liver whose biosynthetic activities strongly impact on the egg’s internal (yolk) and external (eggshell) quality.
Chick embryos depend on the maternal antibodies and nutrients deposited in the yolk, including vitamin D3, carotenoids, and fatty acids, to develop normally. Eggshell quality, among other things, affects the embryo’s access to oxygen, which is especially important when it develops body tissues.
Hens’ ability to form healthy eggs depends on their diet and health. Research indicates that, via the impact on egg formation, broiler breeders’ feeding program quantifiably influences their progeny’s immune system and intestinal health. There is indeed a direct relationship between parent and offspring’s gut health because the chick’s microbiome is in part also inherited from the hen. The impact on DOC quality is thus one of many dimensions to consider when calibrating one’s broiler breeders feeding approach.
The challenge of feeding an ABF broiler breeder
Just as their offspring, breeder hens are genetically predisposed for rapid growth and muscle development. From rearing right through to the laying period, poultry nutritionists need to carefully balance their diets and moderate weight gain in order for hens to reach their reproductive potential.
Different stages of a breeder’s life cycle come with different objectives – for example, good flock uniformity in the rearing period versus egg size and hatchability in the laying phase – and thus different requirements in terms of calories, amino acids, vitamins, and minerals. What remains constant is that the actual nutrient intake depends on intestinal health, determining both the breeders’ performance and, via the impact on egg characteristics, its progeny’s performance.
Can phytomolecules improve broiler breeders’ performance?
Among the plethora of feed additives, phytomolecules, or secondary plant compounds, stand out as a class of active ingredients that may help to improve gut health and thereby reduce the use of antibiotics. Synthesized by plants as a defense mechanism against pathogens, phytomolecules combine digestive, antimicrobial and antioxidant properties.
Some studies have shown that phytomolecules-based products can increase broilers’ body weight gain and improve laying hens’ laying rate, egg mass and egg weight. Both broilers and laying hens responded to the inclusion of phytomolecules in their diet with inclusion rate-dependent improvements in feed conversion. To evaluate if phytomolecules could similarly improve broiler breeders’ performance, two trials were conducted.
Study I: Effect of phytomolecules on laying performance during peak production
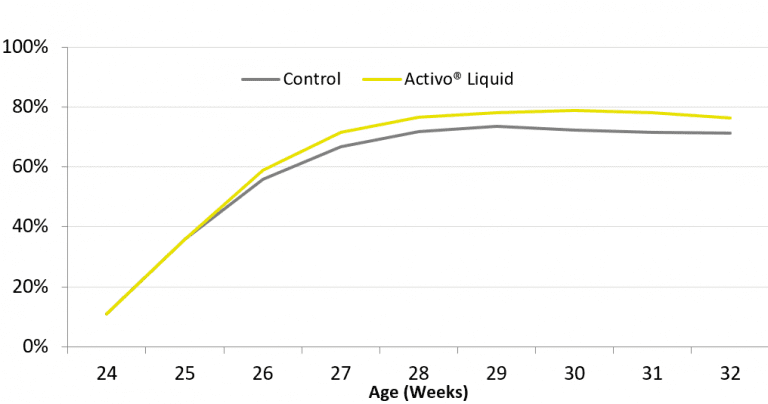
The first study was set up on a farm in Thailand. In total, 40000 Cobb broiler breeders (85% female, 15% male) were divided into two groups with 8500 hens (one house) in the control and 25500 (three houses) in the trial group. Both groups were fed standard feed. The trial group additionally received a phytomolecules-based liquid complementary feed (Activo® Liquid, EW Nutrition GmbH) via the waterline from week 24 to week 32 at a rate of 200ml/1000L during 5 days per week.
Activo® Liquid was found to have a positive influence on laying performance (Figure 1). The average laying rate increased by 7.2% during the trial period, resulting in almost 3 additional hatching eggs per hen housed. A further indication of the beneficial influence that this particular combination of phytomolecules had on gut health was a 0.2% lower mortality.
Figure 1: Laying rate (%) of breeder hens during first 9 weeks of production
Study II: Effect of phytomolecules on laying performance after peak production
For a second study, conducted in the Czech Republic, 800 female and 80 male Hubbard breeders (JA57 and M77, respectively) were divided into 2 groups with 5 replicate pens and 80 female and 8 male breeders per pen. The experiment started after the peak-production period, at 34 weeks of age and ended at 62 weeks of age. All animals received a standard mash diet. For one group a phytogenic premix (Activo®, EW Nutrition GmbH) was added to the diet at a rate of 100g/MT.
The results indicate that Activo® helped maintain the breeder hens’ egg laying performance close to the breed’s genetic potential (Figure 2). In the course of the experiment, Activo® supplemented birds produced 3.6 more eggs than control birds, while consuming a similar amount of feed. As a result, feed consumption per egg produced was lower for birds receiving phytomolecules than for the control birds (169.9 versus 173.6g/d, respectively).
As hatchability was not influenced by the dietary treatment in this study (P>0.5), the 3.6 extra eggs resulted in 2.9 extra day old chicks per hen produced, during the post-peak period alone.
The microencapsulated, selected phytomolecules contained in Activo® are likely to have improved gut health and feed digestibility, and thereby enhanced the animals’ feed efficiency.
Figure 2: Laying rate (%) of breeder hens week 35 till 62
Chicken or egg? Antibiotic-free poultry production looks at the bigger picture
To successfully produce antibiotic-free poultry meat requires a systematic re-think of each component of the production process. Broiler breeders’ lay the foundation for their progeny’s health and performance via the egg. Breeder hens need to be in optimal health to consistently deliver optimal eggs. Without recourse to antibiotics for maintaining or recovering intestinal functionality, an effective ABF production needs to make gut health central to its feeding approach.
The trials reviewed demonstrate that selected phytomolecules quantifiably boost breeders’ laying performance, increasing the number of hatching eggs and DOCs, while reducing mortality and feed consumption per egg produced. As part of an intelligent antibiotic reduction strategy, the right phytogenic products can be potent tools to help poultry producers achieve their NAE objectives.
by S. Regragui Mazili, T. van Gerwe and M. Caballero
References
Calini, F., and F. Sirri. “Breeder Nutrition and Offspring Performance.” Revista Brasileira De Ciência Avícola 9, no. 2 (2007): 77-83. doi:10.1590/s1516-635×2007000200001.
Understanding the dangers of mycotoxins for breeder hens
As the producers of hatching eggs and day-old chicks, breeding hens are the backbone of the poultry industry. Hence it is common practice to pay particular attention to this valuable asset’s feed, selecting raw materials of high nutritional quality and safety. However, in any feed formulated for animals in production and reproduction, studies show that it is almost inevitable to find a certain level of mycotoxin contamination.
Mycotoxins exert toxic effects mainly on the gastrointestinal tract, liver, and kidneys and can accumulate in some tissues but also in the eggs. Mycotoxin contamination in breeder hens rations does not always lead to visible symptoms, such as when trichothecenes cause oral lesions. However, it may influence productivity, egg quality, hatchery performance, as well as chick quality and immunity. Mycotoxin risk management is thus an essential part of managing breeder hens. Mycotoxins can negatively affect eggshell quality and, as a consequence, embryonic mortality.
Type of mycotoxin and exposure time determine effect on egg production
Mycotoxicosis in hens can cause reduced egg production, most likely because it causes a decrease in protein synthesis. A lower synthesis of albumin results from a degeneration of the liver tissue due to aflatoxin, ochratoxin, T2 and DON exposure. The liver then may look pale, friable and occasionally shows superficial hemorrhages.
The contamination levels at which these effects can be observed are as low as 100ppb in feed, for example, during a 21-day exposure to ochratoxin (Figure 1). With increasing levels of the toxin, production further decreases. A similar effect is observed when breeder hens are exposed to aflatoxins.
Figure 1 – Effect of mycotoxins on egg production, compared to non-contaminated control (=100 %)
Egg production, however, is not the only parameter that is affected when breeding hens are exposed to mycotoxins. Earlier on in the reproductive cycle, they already impact on embryonic mortality and hatchability. These effects are potentially more severe and may even occur without any noticeable change in the number of eggs produced.
Mycotoxins’ insidious consequences for eggshell quality and embryonic mortality
The eggshell is important to protect the progeny: thin and fragile shells can increase embryonic mortality, lower embryonic weight gain and decrease hatchability. Eggshell quality is a function of the hen’s calcium and vitamin D3 metabolism. The bioavailability of calcium and of vitamin D3 depends on intestinal integrity and on the production of enzymes and transporters that aid in feed metabolism. These processes can be adversely affected by aflatoxins, DON, T2, and Fumonisins.
The gastrointestinal tract is not the only site of mycotoxin action, however. Mycotoxins such as aflatoxins and ochratoxins have nephrotoxic effects, affecting calcium metabolism and increasing its excretion via the urine, while lowering its levels in blood serum.
Moreover, mycotoxins damage the liver, which plays a central role in egg production, being responsible for vitamin D3 metabolism and the synthesis of the lipids that make up the yolk. Moreover, the synthesis of transporters for lipids, calcium, and carotenoids ̶ important components of the egg ̶ also takes place in the liver. When liver function is impaired, the internal and external quality of the egg declines, which, in the end, affects the production of day-old chicks.
Figure 2 – Effects of mycotoxins on eggshell quality and embryonic mortality
Figure 2 summarises the possible ways in which mycotoxins can negatively affect eggshell quality and, as a consequence, increase embryonic mortality. If a hen’s intestinal integrity is compromised, the utilization of nutrients decreases. Liver and kidney damage leads to a diminished availability of calcium and other nutrients necessary for egg formation. The birds’ calcium (and phosphorus) levels in the plasma are then lower and may lead to a greater mobilization of calcium from the bones. However, this response cannot be maintained and the eggs get a thinner shell.
The thickness of the eggshell influences the egg’s moisture loss and exchange with the environment during the incubation period. An eggshell of optimal quality does not allow the loss of nutrients and prevents bacterial contamination. Thinner eggshells are less able to fulfill these functions, leading to higher embryo mortality.
Figure 3 – Effects of mycotoxins on embryonic mortality
Figure 3 shows the effect of different mycotoxins on embryonic mortality. Incremental levels of ochratoxin or aflatoxin heighten embryonic mortality in a range from 1.5 to 7.5 times the embryonic mortality of the control group. In some cases, embryos are affected even when the hens received feed contaminated with mycotoxin levels that are within the guidelines suggested by the EFSA.
For example, an exposure to 4900ppb of DON for ten weeks increases the number of embryos with abnormalities. The causes are not entirely clear, as only traces of DON can be found in the egg. However, we do know that this mycotoxin can affect the protein synthesis at the level of the hen’s liver and therefore compromise the deposition of nutrients into the egg.
Mycotoxins’ effects on the progeny may cause long-term damage
Ochratoxin and aflatoxin can be transferred into the egg, where they exert toxicity on the embryos. This does not necessarily result in mortality. However, the chicks can suffer from a compromised immune function due to two reasons: lower transmission of antibodies from the hen and lower viability of the chickens’ immune cells, accompanied by a lower relative weight of the bursa of Fabricio and the thymus.
When both aflatoxin and ochratoxin are present in the feed, the effect on these parameters is synergistic. As a consequence of mycotoxin contamination, the animals’ immune response is impaired, which makes them more susceptible to infection. The final result could be increased early chick mortality due to a higher incidence of bacterial and viral infections.
The transmission of other mycotoxins into the egg is minimal. While this means that a direct effect on the progeny is unlikely to occur, mycotoxin contamination still has a snowball effect: we have to consider the indirect effect of a lower deposition of nutrients on chick quality.
Prevention is key: mycotoxin risk management for breeder hens
The best approach to manage mycotoxin risk is to implement an integrated strategy that includes good crop and grain storing practices, regular raw material sampling and mycotoxin evaluation and analysis. Management tools (such as MasterRisk) can help to evaluate mycotoxin interactions and to choose the best strategy for dealing with specific mycotoxin challenges.
The results of mycotoxin analyses can be used to take decisions regarding the inclusion levels of raw materials and in choosing feed additives that counteract mycotoxins. Products based on plant extracts, yeast cell walls, and clay minerals can help to stabilize a digestive system challenged by mycotoxins. They support the barrier function in the intestine, preventing the passage of mycotoxins into the bloodstream.
Phytomolecules are another piece of the puzzle: thanks to their antimicrobial, anti-inflammatory and antioxidant properties, they support liver function. This is particularly important for long-living animals prone to accumulating mycotoxins in their body tissues.
For a long time the “deleterious effects” of mycotoxins on breeder hens and “their repercussions on progeny health status and performance have not received from a scientific point of view as much attention”(Calini and Sirri, 2007) as they ought to have. However, now that the dangers of mycotoxins for breeder hens’ welfare, health and performance are better understood, it is clear that mycotoxin risk evaluation and management is central to successful poultry production.
*This article first appeared in All About Feed on 31 October 2018
Nowadays, dairy cows are real top athletes. This comes with additional challenges for their health and for on-farm management. Many of these problems can be traced back to supply deficits and can be easily managed with appropriate feed supplements.
Milk fever is a disease that occurs mainly in cows around calving. It is caused by an insufficient amount of calcium in the blood and particularly affects cows with a very high milk yield.
The link between calcium and milk fever
Calcium performs essential functions in the body. It is particularly important for the nervous system and muscle cells, and plays a central role in muscle contraction. If the calcium content in the blood is too low, the muscles can no longer contract. When this happens, the cows cannot move or stand up.
While mild cases may not be easily detectable, they still trigger productivity loss. If undetected, long-term calcium deficiency can even lead to cardiac arrest and thus to the death of the animal.
The development of milk fever
The cause of milk fever is a lack of sufficient calcium in the blood serum (hypocalcemia). The dairy cow has to abruptly change its metabolism at the end of the dry period, going from the resting phase to a high performance phase. During the dry period, cows have a relatively low need for calcium.
When lactation starts, the need for calcium suddenly almost doubles, as large amounts of calcium are required for the production of colostrum (2.3 g/l). The calcium is generally drawn from feed or from the bones. In older cows, the mobilization mechanism often does not start quickly enough. The supply from the bones and feed is insufficient and the body draws the missing calcium from the muscles. This ultimately leads to symptoms of paralysis and overstimulation of the nervous system.
Phases of milk fever
Stage One
In the initial phase of milk fever, the initial signs are
muscle tremors
restlessness
stiff gait
slightly elevated temperature
Stage Two
At this point, the cows lie on the stomach with an extended neck or the head is lying on the flank. Early symptoms of paralysis appear:
fast, flat pulse
cold body surface
dilated pupils
flatulence
Stage Three
In the last phase of milk fever, the cow lies on its side, loses consciousness and falls into a coma. The third phase often leads to death (the mortality rate averages 2 – 5%).
While the second phase of milk fever is easy to recognize due to the clear symptoms, the consequences of a “slight” calcium deficiency (Stage One) are often underestimated. Feed intake diminishes, the negative energy and protein balance is increased, and the cows barely move. The impairment of the muscles can cause problems in the udder (mastitis) or in the gastrointestinal tract.
Prevention and solutions
As cases of hypocalcemia immediately after calving may be as high as 50% among second- or third-lactation cows, it is important to act preventively to keep potential milk fever from developing. The dairy farmer´s aim is to support the dairy cows that are at higher risk of milk fever, especially around the critical time of calving. The cows must be enabled to quickly release calcium from the bones after calving, or they must be supplied with calcium that can be easily metabolized.
Upfront prophylaxis
An energy and protein oversupply during the dry period should be avoided. In addition, an application of Vitamin D3 at the end of the pregnancy makes sense.
To stimulate the active regulatory mechanisms of calcium metabolism, the calcium content in the feed should be reduced three to four weeks before calving. In practice, however, this often is not properly observed and feed with a relatively high calcium content is still given out during this period.
There are, no doubt, farms where these above-mentioned preventive measures cannot be carried out due to operational reasons, just as there are animals that are particularly susceptible due to factors such as age, breed or healthy history.
To protect the cow from milk fever around calving, oral administration of calcium salts is widespread in practice. Vitamin D also plays a central role in calcium metabolism. It ensures that the absorption of calcium from the intestines and bones is increased.
When administering oral calcium supplements, there are three important points:
– The cow must have sufficient calcium available per dosage
– The calcium must be available immediately
– Administration must be appropriate for the animals and farmers
Methods of calcium supplementation
To support the cow, oral supplements such as pastes and gels are widely used. They are useful, however they are also relatively difficult to administer, as they require handling the animal in relatively difficult ways.
Liquids are another way of administering calcium supplements. When administering liquids, it is important to make sure the animal does not choke so that the liquids do not get into the lungs.
Boluses are probably the easiest and safest method of supplementation to prevent milk fever. The bolus must naturally be carefully inserted, however the process is easy and requires minimal handling of the animal.
EW Nutrition´s Calzogol Bolus is a dietetic mineral feed with a high level of calcium from of highly available calcium salts and vitamin D3. The Calzogol Bolus contains several calcium sources with different release rates. One major advantage is the very high mucous membrane compatibility, which helps avoid irritation of the mouth, esophagus and rumen. Furthermore, the Calzogol Bolus does not contain caustic calcium chloride. The application is simple and economical, as only one bolus per dose must be administered at the time of calving.
Conclusion
Milk fever is very common in dairy herds. When a cow has milk fever, the farm can incur costs of approx. €350. This is reflected in the loss of milk yield up to 600 kg, losses due to unusable milk, and veterinary and medication costs.
Time resources are also to be taken into account: The economic repercussions represent a significant factor, however they come on top of the extra workload due to the increased need for care of animals.
Cows that suffer from calcium deficiency are also much more susceptible to other diseases. For the farmer, the best strategy is to avoid losses through prophylaxis. Feeding plays a central role; to ensure the best possible production conditions, oral calcium administrations, such as Calzogol Bolus, have proven themselves in practice.
by Judith Schmidt, Product Manager, On Farm Solutions
References:
Rérat, M. (2005): Milchfieber bei der Milchkuh. ALP aktuell. Nr. 20.
Spiekers, H., Potthast, V. (2004): Erfolgreiche Milchviehfütterung. DLG-Verlag, Frankfurt a. M.
Kirchgeßner, M., Roth, F. X., Schwarz, F. J., Stangl, G. I. (2008): Tierernährung. 12. Auflage. DLG-Verlag, Frankfurt a. M.
Are endotoxins behind your low livestock productivity?
by Dr. Inge Heinzl, EW Nutrition
Impaired health status of the animals in stressful situations or an aggravation of the disease after antibiotic treatment? The culprit might be endotoxins.
What are endotoxins?
Origin
Endotoxins, together with exotoxins, are bacterial toxins. In contrast to exotoxins, which are actively secreted by living bacteria, endotoxins (name “endotoxin” greek; endo = inside; toxin = poison) are components of the outer cell membrane of gram-negative bacteria such as Escherichia coli, Salmonella, Shigella, and cyanobacteria (blue-green algae). They are only released in case of
bacterial death due to effective host defense mechanism or activities of certain antibiotics
Biochemically, endotoxins are lipopolysaccharides (LPS). They are composed of a relatively uniform lipid fraction (Lipid A) and a species-specific polysaccharides chain. Their toxicity is mainly due to the lipid A; the polysaccharide part modifies their activity. Unlike the bacteria, their endotoxins are very heat stable and resist sterilization. The names endotoxin and lipopolysaccharides are used synonymously with “endotoxin” emphasizing on the occurrence and biological activity and “lipopolysaccharide” on the chemical structure (Hurley, 1995).
General structure of Gram-negative lipopolysaccharides (according to Erridge et al., 2002)
Impact
Endotoxins belong to the so-called pyrogen-agents (they provoke fever), activating several immunocompetent cells’ signaling pathways. Early contact with endotoxins leads to activation and maturation of the acquired immune system. Braun-Fahrländer and co-workers (2002) found that children exposed to endotoxins had fewer problems with hay fever, atopic asthma, and atopic sensitization. This might be an explanation that in human populations, after the elevation of the hygiene standards, an increase of allergies could be observed.
Different animal species show different sensibilities to endotoxin infusions, e.g. (healthy) dogs, rats, mice, hens tolerate concentrations ≥1mg / kg body weight, whereas (healthy) ruminants, pigs, horses react very sensible already at concentrations <5μg / kg body weight (Olson et al., 1995 cited in Wilken, 2003).
Reasons for increased exposure of the organism to endotoxins
Endotoxins usually occur in the gut, as the microflora also contains gram-negative bacteria. The precondition for endotoxins to be harmful is their presence in the bloodstream. In the bloodstream, low levels of endotoxins can still be handled by the immune defense, higher levels can get critical. An increase of endotoxins in the organism results from higher input and/or lower clearance or detoxification rate.
Higher input of endotoxins into the organism
The “normal” small amounts of endotoxins arising in the gut due to regular bacterial activity and translocated to the organism have no negative impact as long as the liver performs its clearance function. Also, the endotoxins stored in the adipose tissue are not problematic. However, some factors can lead to a release of the endotoxins or translocation of endotoxins into the organism:
1. Stress
Stress situations such as parturition, surgeries, injuries can lead to ischemia in the intestinal tract and translocation of endotoxins into the organism (Krüger, 1997). Other stress situations in animal production, such as high temperatures and high stocking densities, contribute to higher endotoxin levels in the bloodstream. Stress leads to a higher metabolic demand for water, sodium, and energy-rich substances. For a higher availability of these substances, the intestinal barrier’s permeability is increased, possibly leading to a higher translocation of bacteria and their toxins into the bloodstream.
Examples:
Higher levels of endotoxins in pigs in an experimental study suffering from stress due to loading and transport, elevated temperatures (Seidler (1998) cited in Wilken (2003)).
Marathon runners (Brock-Utne et al., 1988) and racing horses (Baker et al., 1988) also showed higher endotoxin concentrations in the blood proportional to the running stress; thus, trained horses showed lower concentrations than untrained.
2. Lipolysis for energy mobilization
If endotoxins, due to continuous stress, consistently get into the bloodstream, they can be stored in the adipose tissue. The SR-B1 (Scavenger receptor B1, a membrane receptor belonging to the group of pattern recognition receptors) binds to lipids and the lipopolysaccharides, probably promoting the incorporation of LPS in chylomicrons. Transferred from the chylomicrons to other lipoproteins, the LPS finally arrives in the adipose tissue (Hersoug et al., 2016). The mobilization of energy by lipolysis e.g., during the beginning of lactation, for example, leads to a re-input of endotoxins into the bloodstream.
3. Damage of the gut barrier
In normal conditions, due to bacterial activity, endotoxins are present in the gut. Damage of the gut barrier allows translocation of these endotoxins (and bacteria) into the bloodstream.
4. Destruction of Gram-negative bacteria
Another “source” for endotoxins is the destruction of the bacteria. This can be done on the one hand by the organism’s immune system or by treatment with bactericidal substances targeting gram- bacteria (Kastner, 2002). To prevent an increased release of endotoxins, in the case of Gram-negative bacteria, a treatment with bacteriostatic substances only inhibiting the growth and not destroying the bacteria, or with bactericidal in combination with LPS-binding agents, would be a better alternative (Brandenburg, 2014).
5. Proliferation of gram-negative bacteria
As gram-negative bacteria also release small amounts of endotoxins when they grow, everything promoting their proliferation also leads to an increase of endotoxins:
Imbalanced feeding
High yielder cows e.g., are fed diets rich in starch, fat, and protein. Increased feeding of fat leads to a higher concentration of endotoxins in the organism, as the same “transporter” (scavenger receptor class B type 1, SR-BI) can be used (Hersoug et al., 2016) for the absorption of fat as well as for the absorption of endotoxins.
In a study with humans as representors of the monogastric species, Deopurkar and co-workers gave three different drinks (glucose – 100% carbohydrate, orange juice – 92% carbohydrate, and cream – 100% fat) to healthy participants. Only the cream drink increased the level of lipopolysaccharides in the plasma.
Infectious diseases
Infectious diseases like mastitis, metritis, and other infections caused by gram-bacteria such as E. coli, Salmonella, etc. can be regarded as sources of endotoxin release.
Decreased detoxification or degradation
Main responsible organ: the liver
Task: detoxification and degradation of translocated endotoxin. The liver produces substances such as lipopolysaccharide binding proteins (LBP) which are necessary for binding and neutralizing lipopolysaccharide structures.
During the post-partum period, the organism is in a catabolic phase, and lipolysis is remarkably increased for energy generation due to milk production. Increased lipolysis leads, as mentioned before, to a release of endotoxins out of the adipose tissue but also fatty degeneration of the liver. A fatty degenerated liver cannot bring the same performance in endotoxin clearance than a normal liver (Andersen, 2003; Andersen et al., 1996; Harte et al., 2010; Wilken, 2003). In a study conducted by Andersen and co-workers (1996), they couldn’t achieve complete clearance of endotoxins in cows with fatty livers. The occurrence of hepatic lipidoses increases after parturition (Reid and Roberts, 1993; Wilken, 2003).
Also, other diseases of the liver influence endotoxin clearance in the liver. Hanslin and co-workers (2019) found an impaired endotoxin elimination in pigs with pre-existing systemic inflammatory response syndrome.
Relation between lipid metabolism and endotoxin metabolism (according to Fürll, 2000, cited in Wilken, 2003)
Issues caused by endotoxins
Endotoxins, on the one hand, can positively stimulate the immune system when occurring in small amounts (Sampath, 2018). According to McAleer and Vella (2008), lipopolysaccharides are used as natural adjuvants to strengthen immune reaction in case of vaccination by influencing CD4 T cell responses. On the other hand, they are involved in the development of severe issues like MMA-Complex (Pig Progress) or a septic shock (Sampath, 2018).
MMA Complex in sows
MMA in sows is a multi-factorial disease appearing shortly after farrowing (12 hours to three days), which is caused by different factors (pathogens such as E. coli, Klebsiella spps., Staph. spps. and Mycoplasma spps., but also stress, diet). MMA is also known as puerperal syndrome, puerperal septicemia, milk fever, or toxemia. The last name suggests that one of the factors intervening in the disease is bacterial endotoxins. During the perinatal phase, massive catabolism of fat takes place to support lactation. The sows often suffer from obstipation leading to higher permeability of the intestinal wall, with bacteria, respectively endotoxins being transferred into the bloodstream. Another “source” of endotoxins can be the udder, as the prevalence of gram-negative bacteria in the mammary glands is remarkable (Morkoc et al., 1983).
The endotoxins can lead to an endocrine dysfunction: ↑ Cortisol, ↓ PGF2α, ↓Prolactin, ↓ Oxytocin. MMA stands for:
– Mastitis, a bacterial infection of the udder.
Mastitis can be provoked from two sides: on the one hand, endotoxemia leads to an elevation of cytokines (IL1, 6, TNFα). Lower Ca- and K-levels cause teat sphincter to be less functional, facilitating the entry of environmental pathogens into the udder and resulting in mastitis. On the other hand, due to farrowing stress, Cortisol concentrations get higher. The resulting immunosuppression enables E. coli to proliferate in the udder.
– Metritis, an infection of the uterus with vulvar discharges:
It leads to reduced contractions and, therefore, to prolonged and/or complicated farrowing or dead piglets. Metritis can be promoted by stress causing a decrease in oxytocin and prostaglandin F2α secretion. Morkoc and co-workers (1983) didn’t find a relation between metritis and endotoxins.
– Agalactia, a reduction or total loss of milk production:
In many cases, agalactia is not detected until the nursing litter shows signs of hunger and/or weight loss. At worst, the mortality rate in piglets increases. Probably, milk deficiency is caused by lower levels of the hormones involved in lactation. Prolactin levels e.g., may be dramatically reduced by small volumes of endotoxin (Smith and Wagner, 1984). The levels of oxytocin are often half those in normal sows (Pig Progress, 2020).
Endotoxin shock
A septic shock can be the response to the release of a high amount of endotoxins.
In the case of an infection with gram-negative bacteria, the animals are treated with (often bactericidal) antibiotics. Also, the immune system is eliminating the bacteria. Due to bacterial death, endotoxins are massively released. When not bound, they activate the immune system including macrophages, monocytes, and endothelial cells. Consequently, high amounts of cellular mediators like TNFα, Interleukin 1 (IL-1), IL-6, and leukotrienes are released. High levels of pro-inflammatory cytokines activate the complement and coagulation cascade. In some animals, then the production of prostaglandins and leukotrienes is stimulated, implicating high fever, decreased blood pressure, generation of thrombi in the blood, collapse, damaging several organs, and lethal (endotoxic) shock.
Endotoxic shock only occurs to a few susceptible animals, although the whole herd may have been immune-stimulated. A more severe problem is the decrease in the normally strong piglets’ performance, deviating resources from production to the immune system because of the endotoxemia.
Amplified diarrhea
Lipopolysaccharides lead to an augmented release of prostaglandins, which influence gastrointestinal functions. Together with leukotrienes and pro-inflammatory mediators within the mucosa, they reduce intestinal absorption (Munck et al., 1988; Chiossone et al., 1990) but also initiate a pro-secretory state in the intestine. Liang and co-workers (2005) observed a dose-dependent accumulation of abundant fluid in the small intestine resulting in increased diarrheagenic activity and a decreased gastrointestinal motility in rats.
Conclusion
Acting against Gram- bacteria can result in an even more severe issue – endotoxemia. Endotoxins, besides having a direct negative impact on the organism, also contribute to some diseases. Supporting gut health by different approaches, including the binding of toxins, helps to keep animals healthy.
References
Andersen, P.H. “Bovine endotoxicosis – some aspects of relevance to production diseases. A review.” Acta vet. scand. Suppl. 98 (2003): 141-155. DOI: 10.1186/1751-0147-44-S1-P57
Andersen, P.H., N. Jarløv, M. Hesselholt, and L. Bæk. “Studies on in vivo Endotoxin Plasma Disappearance Times in Cattle.” Zentralblatt für Veterinärmedizin. Reihe A 43 no. 2(1996): 93-101. DOI: 10.1111/j.1439-0442.1996.tb00432.x
Baker, B., S.L. Gaffin, M. Wells, B.C. Wessels and J.G. Brock-Utne. “Endotoxaemia in racehorses following exertion.” Journal of the South African Veterinary Association June (1988): 63-66. https://journals.co.za/docserver/fulltext/savet/59/2/1341.pdf?expires=1598542211&id=id&accname=guest&checksum=E50C766D318776E09CA41DA912F14CAD
Beutler, B. and T. Rietschel. “Innate immune sensing and its roots: The story of endotoxin.” Nature Reviews / Immunology 3(2003): 169-176. DOI: 10.1038/nri1004
Braun-Fahrländer, C., J. Riedler, U. Herz, W. Eder, M. Waser, L. Grize, S. Maisch, D. Carr, F. Gerlach, A. Bufe, R.P. Lauener, R. Schierl, H. Renz, D. Nowak and E. von Mutius. „Environmental exposure to endotoxin and its relation to asthma in school-age children. ”The New England Journal of Medicine 347 (2002): 869-877. DOI: 10.1056/NEJMoa020057.
Chiossone, D. C., P.L. Simon, P.L. Smith. “Interleukin-1: effects on rabbit ileal mucosal ion transport in vitro.” European Journal of Pharmacology 180 no. 2-3 (1990): 217–228. DOI: 10.1016/0014-2999(90)90305-P.
Deopurkar R., H. Ghanim, J. Friedman, et al. “Differential effects of cream, glucose, and orange juice on inflammation, endotoxin, and the expression of Toll-like receptor-4 and suppressor of cytokine signaling-3.” Diabetes care 33 no. 5 (2010):991–997.
Erridge, C., E. Bennett-Guerrero, and I.R. Poxton. “Structure and function of lipopolysaccharides.” Microbes and Infection 4 no. 8 (2002): 837-851. DOI: 10.1016/s1286-4579(02)01604-0
Fritsche, D. “Endotoxinpromovierte bakterielle Translokationen und Besiedelung von Uterus und Euter beim Hochleistungsrind im peripartalen Zeitraum.“ Dissertation. Leipzig, Univ., Veterinärmed. Fak. (1998)
Hanslin, K., J. Sjölin, P. Skorup, F. Wilske, R. Frithiof, A. Larsson, M. Castegren, E. Tano, and M. Lipcsey. “The impact of the systemic inflammatory response on hepatic bacterial elimination in experimental abdominal sepsis.” Intensive Care Medicine Experimental 7 (2019): art. 52. https://doi.org/10.1186/s40635-019-0266-x
Harte, A.L., N.F. da Silva, S.J. Creely, K.C. McGee, T. Billyard, E.M. Youssef-Elabd, G. Tripathi, E. Ashour, M.S. Abdalla, H.M. Sharada, A.I. Amin, A.D. Burt, S. Kumar, C.P. Day and P.G. McTernan. “Research Elevated endotoxin levels in non-alcoholic fatty liver disease.” Journal of Inflammation 7 (2010): 15-24. DOI: 10.1186/1476-9255-7-15
Hersoug, L.-G., P. Møller, and S. Loft. “Gut microbiota-derived lipopolysaccharide uptake and trafficking to adipose tissue: implications for inflammation and obesity.” Obesity Reviews 17 (2016): 297–312. DOI: 10.1111/obr.12370
Hurley, J. C. “Endotoxemia: Methods of detection and clinical correlates.” Clin. Microbiol. Rev. 8 (1995): 268–292. DOI: 10.1128/CMR.8.2.268
Kastner, A. “Untersuchungen zum Fettstoffwechsel und Endotoxin-Metabolismus bei Milchkühen vor dem Auftreten der Dislocatio abomasi.“ Inaug. Diss. Universität Leipzig, Veterinärmed. Fak. (2002). https://d-nb.info/967451647/34
Krüger M. “Escherichia coli: Problemkeim in der Nutztierhaltung.“ Darmflora in Symbiose und Pathogenität. Ökologische, physiologische und therapeutische Aspekte von Escherichia coli. 3. Interdisziplinäres Symposium. Alfred-Nissle-Gesellschaft (Ed.). Ansbach, 28.-29. Nov. (1997): 109-115.
Liang, Y.-C., H.-J. Liu, S.-H. Chen, C.-C. Chen, L.-S. Chou, and L. H. Tsai. “ Effect of lipopolysaccharide on diarrhea and gastrointestinal transit in mice: Roles of nitric oxide and prostaglandin E2.” World J Gastroenterol. 11 no. 3 (2005): 357–361. DOI: 10.3748/wjg.v11.i3.357
McAleer, J.P. and Vella, A.T. “Understanding how lipopolysaccharide impacts CD4 T cell immunity.” Crit. Rev. Immunol. 28 no. 4 (2008): 281-299. DOI:10.1615/CRITREVIMMUNOL.V28.I4.20
Morkok, A., L. Backstrom, L. Lund, A.R.Smith. “Bacterial endotoxin in blood of dysgalactic sows in relation to microbial status of uterus, milk, and intestine.” JAVMA 183 (1983): 786-789. PMID: 6629987
Munck, L.K., A. Mertz-Nielsen, H. Westh, K. Buxhave, E. Beubler, J. Rask-Madsen. “Prostaglandin E2 is a mediator of 5-hydroxytryptamine induced water and electrolyte secretion in the human jejunum.” Gut 29 no. 10 (1988): 1337-1341
Sampath, V.P. “Bacterial endotoxin-lipopolysaccharide; structure, function and its role in immunity in vertebrates and invertebrates.” Agriculture and Natural Resources 52 no. 2 (2018): 115-120. https://doi.org/10.1016/j.anres.2018.08.002
Seidler, T. “Freies Endotoxin in der Blutzirkulation von Schlachtschweinen: eine Ursache für bakterielle Translokationen?“ Diss. Universität Leipzig, Veterinärmed. Fak. (1998).
Smith, B.B. and W.C. Wagner. “Suppression of prolactin in pigs by Escherichia coli endotoxin.“ Science 224 no. 4649 (1984): 605-607
Wilken, H. “Endotoxin-Status und antioxidative Kapazität sowie ausgewählte Stoffwechselparameter bei gesunden Milch- und Mutterkühen.“ Inaugural Diss. Universität Leipzig (2003).
Necrotic enteritis: The complete overview
by Inge Heinzl and Technical Team, EW Nutrition
Necrotic enteritis is a profit killer in poultry production
Necrotic enteritis is the cause of USD 6 billion losses every year in global poultry production, corresponding to USD 0.0625 per bird (Wade and Keyburn, 2015). This controllable disease is on the rise. One reason is the voluntary or legally required reduction of antibiotics in animal production due to the increasing occurrence of antimicrobial resistance but also consumer demand. Another reason is the administration of live Coccidiosis vaccines and partial reduction of ionophores, which also show efficacy against Gram-positive bacteria (Williams, 2005).
Necrotic enteritis and coccidiosis are the most significant health problem in broilers (Hofacre et al., 2018).
The disease generally occurs in broiler chickens of 2-6 weeks of age. It is caused by an overgrowth of Clostridium perfringens type A and, to a lesser extent, type C in the small intestine. The toxins produced by C. perfringens also damage the intestinal wall.
Clinical and subclinical forms of NE – which one causes more significant losses?
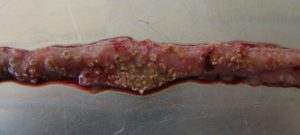
The clinical form is obvious
Intestine showing signs of NE
…is characterized by acute, dark diarrhea resulting in wet litter and suddenly increasing flock mortality of up to 1% per day after the first clinical signs appear (Ducatelle and Van Immerseel, 2010), sometimes summing up to mortality rates of 50% (Van der Sluis, 2013). The birds have ruffled feathers, lethargy, and inappetence.
Necropsy typically shows ballooned small intestines with a roughened appearing mucosal surface, lesions, and brownish (diphtheritic) pseudomembranes. There is a lot of watery brown, blood-tinged fluid and a foul odor during post-mortem examination. The liver is dark, swollen, and firm, and the gall bladder is distended (Hofacre et al., 2018).
In the case of peracute Necrotic Enteritis, birds may die without showing any preliminary signs.
The subclinical form often only can be noticed at the end of the cycle
When birds suffer from the subclinical form, chronic damage to the intestinal mucosa and an increased quantity of mucus in the small intestine lead to impaired digestion and absorption of nutrients resulting in poor growth performance. The deteriorated feed conversion and the resulting decreased performance become noticeable around day 35 of age. As feed contributes approximately 65-75% of the input cost to produce a broiler chicken, poor feed conversion increases production costs and significantly influences profitability. Often, due to a lack of clear symptoms, this subclinical disease remains untreated and permanently impacts the efficiency of production.
The pathogen causing NE – a ubiquitous bacterium
Responsible for Necrotic Enteritis are Gram-positive, anaerobic bacteria, specific strains of Clostridium perfringens type A and, to a lesser extent, type C (Keyburn et al., 2008).
Clostridia primarily occur in the soil where organic substances are degraded, in sewage, and in the gastrointestinal tract of animals and humans. These bacteria produce spores, which are extremely resistant to environmental impact (heat, irradiation, exsiccation), some disinfectants, and can survive for several years. Under suitable conditions, C. perfringens spores can even proliferate in feed or litter.
Clostridium perfringens is a “natural inhabitant” of the intestine of chickens. In healthy birds, it occurs in a mixture of diverse strains at 102-104 CFU/g of digesta (McDevitt et al., 2006). The disease starts when C. perfringens proliferates in the small intestine, usually due to a combination of factors such as high amount protein, low immunity, and an imbalance in the gut flora. Then, the number rises to 107-109 CFU/g of digesta (Dahiya et al., 2005).
Highly important: NetB, a pore-forming toxin is a key virulence factor for NE
To establish in the host, Clostridium Spp. and other pathogens depend on virulence factors (see infobox). These virulence factors include for example “tools” for attachment, evasion or suppression of the host’s immune system, “tools” for getting nutrients, and “tools” for entry into intestinal cells. Over the years, the α-toxin produced by C. perfringens was assumed to be involved in the development of the disease and a key virulence factor. In 2008, Keyburn and coworkers found another key virulence factor by using a C. perfringens mutant unable to produce α-toxin, while still causing Necrotic Enteritis.
Thus, another toxin was identified occurring only in chickens suffering from Necrotic Enteritis: C. perfringens necrotic enteritis B-like toxin (NetB). NetB is a pore-forming toxin. Pore-forming toxins are exotoxins usually produced by pathogenic bacteria but may also be produced by other microorganisms. These toxins destroy the integrity of gut wall cell membranes. The leaking cell contents serve as nutrients for the bacteria. If immune cells are destroyed, an immune reaction might be partially imparted.
Additionally, pathogenic strains of C. perfringens produce bacteriocins – the most important is Perfrin (Timbermont et al., 2014) – to inhibit the proliferation of harmless Clostridium Spp. strains and to replace the normal intestinal flora of chickens (Riaz et al., 2017).
Examples of virulence factors
Adhesins
Enable the pathogen to adhere or attach within the target host site, e.g. via fimbria. Pili enable the exchange of RNA or DNA between pathogens.
Invasion factors
Facilitate the penetration and the distribution of the pathogens in the host tissue (invasion and spreading enzymes). For example: hyaluronidase attacking the hyaluronic acid of the connective tissue or flagella enabling the pathogens to actively move.
Toxins
Damage the function of the host cells or destroy them (e.g. endotoxins – lipopolysaccharides, exotoxins)
Strategies of evasion
Enable the pathogen to undergo the strategies of defense of the immune system (e.g. antiphagocytosis factors provide protection against an attack by phagocytes; specific antibodies are inactivated by enzymes).
Predisposing factors favor the development of NE
A chicken with optimal gut health may be less susceptible to NE. Additional predisposing factors are necessary to allocate nutrients and make the gut environment suitable for the proliferation of these pathogens, enabling them to cause disease (Van Immerseel et al., 2008; Williams, 2005).
1. FEED: composition and particle size are critical
Feed plays a role in the development of Necrotic Enteritis that should not be underestimated. Here, substances creating an intestinal environment favorable for C. perfringens must be mentioned.
2. Mycotoxins create ideal conditions for NE
Mycotoxins harm gut integrity and create ideal conditions for the proliferation of Clostridium perfringens:
Mycotoxins do not have a direct effect on C. perfringens proliferation, toxin production, or NetB transcription. However, mycotoxins disrupt gut health integrity, creating a favorable environment for the pathogen. For example:
DON provides good conditions for proliferation of perfringens by disrupting the intestinal barrier and damaging the epithelium. The possibly resulting permeability of the epithelium and a decreased absorption of dietary proteins can lead to a higher amount of proteins in the small intestine. These proteins may serve as nutrients for the pathogen (Antonissen et al., 2014).
DON and other mycotoxins decrease the number of lactic acid producing bacteria indicating a shift in the microbial balance (Antonissen et al., 2016.)
3. Eimeria spp.: forming a perfect team with Clostridium perfringens
An intact intestinal epithelium is the best defense against potential pathogens such as C. perfringens. Here, coccidiosis comes into play. Moore (2016) showed that by damaging the gut epithelium, Eimeria species give C. perfringens access to the intestinal basal domains of the mucosal epithelium. Then, the first phase of the pathological process takes place and from there, C. perfringens invades the lamina propria. Damage to the epithelium follows (Olkowski et al., 2008). The plasma proteins leaking to the gut and the mucus produced are rich nutrient sources (Van Immerseel et al., 2004; Collier et al., 2008). A further impact of coccidiosis is shifting the microbial balance in the gut by decreasing the number of e.g., Candidatus savagella which activates the innate immune defense.
Figure 1
Figure 1:
Eimeria induce leakage of plasma proteins by killing epithelial cells
They enhance mucus production in the intestine
A+B lead to an increase in available nutrients and create an environment favorable for the proliferation of perfringens
Not only Eimeria Spp., also other pathogens (e.g. Salmonella Spp., Ascarid larvae, viruses) and agents, such as mycotoxins damaging the intestinal mucosa can pave the way for a C. perfringens infection. Predisposing factors like wet litter, the moisture of which is essential for the sporulation of Eimeria Spp. oocysts, must also be considered as promoting factors for Necrotic Enteritis (Williams, 2005).
4. Immunosuppressive Factors: Bacteria, viruses…, and stress
Any factor which induces stress in the animals disrupts the balance of the intestinal flora. The resulting suppression of the immune system contributes to the risk of Necrotic Enteritis (Tsiouris, 2016).
Bacteria
Shivaramaiah and coworkers (2011) investigated a neonatal Salmonella typhimurium infection as a predisposing factor for NE. The early infection causes significant damage to the gut (Porter et al., 1998) Additionally, Hassan et al. (1994) showed that the challenge with Salmonella typhimurium negatively impacted the development of lymphocytes which might also promote a colonization of Clostridium perfringens.
Viruses
Infectious Bursal Disease is known to increase the severity of infections with salmonella, staphylococci, but also clostridia. Another clostridia-promoting viral disease is Marek’s Disease.
Stress:
The intestinal tract is particularly sensitive to any type of stress. This stress can be caused by e.g. too high temperatures, high stocking densities, an abrupt change of feed.
Figure 2: Predisposing factors weakening the birds and enabling Clostridium to attack
Treatment is necessary in the case of acute disease
In this instance, the farmer is obligated to consult a veterinarian and treat his birds.
It must be mentioned that, as the treatment takes place via feed or water, only birds which still consume water or feed may be treated.
Antibiotics are effective but also take a risk
Antibioitics targeting Gram-positive bacteria are commonly used for the treatment of acute NE. The antibiotic choice shall be addressed by a veterinarian, taking into account the mode of action and the presence of resistance genes in the farm/flock.
Some bacteria are less sensitive to certain antibiotics due to genetic mutations. They are able to:
stimulate the production of enzymes, which break down or modify the antibiotics and inactivate them (1).
eliminate entrances for antibiotics or promote the development of pumps, which discharge the antibiotic before taking effect (2).
change or eliminate molecules to which the antibiotic would bind (targets for the antibiotics).
This means that, when the corresponding antibiotics are used, bacteria resistant against these antibiotics survive. Due to the fact that their competitors have been eliminated they are able to reproduce better. Additionally, this resistance may be transferred by means of “resistance genes”
to daughter cells
via their intake from dead bacteria (3)
through horizontal gene transfer (4)
through viruses (5)
Every application of antibiotics promotes the development of resistance (Robert Koch Institute, 2019). A short-term use, better biosecurity, or an application at low dosage give the bacteria a better chance to adapt.
Bacteriophages would be possible but are still disputed
Experimental use of phage treatments has shown to be effective in reducing disease progression and symptoms of Necrotic Enteritis (Miller et al., 2010). By oral application of a bacteriophage cocktail, Miller and coworkers could reduce mortality by 92% in C. perfringens-challenged broilers compared to the untreated control.
Mode of action: the endolysins, highly evolved enzymes produced by bacteriophages, are able to digest the bacterial cell wall for phage progeny release (Fischetti, 2010). However, phages are still not approved by the EFSA.
Excurs:
Antimicrobial Resistance (AMR)Some bacteria are less sensitive to certain antibiotics due to genetic mutations. They are able to:
stimulate the production of enzymes, which break down or modify the antibiotics and inactivate them (1).
eliminate entrances for antibiotics or promote the development of pumps, which discharge the antibiotic before taking effect (2).
change or eliminate molecules to which the antibiotic would bind (targets for the antibiotics) (figure 3)
Figure 3: Possibilities of a bacterial cell to defend itself against antibiotics
This means that, when the corresponding antibiotics are used, bacteria resistant against these antibiotics survive. Due to the fact that their competitors have been eliminated they are able to reproduce better.Additionally, this resistance may be transferred by means of “resistance genes”
to daughter cells
via their intake from dead bacteria (3)
through horizontal gene transfer (4)
through viruses (5) (figure 4)
Figure 4: Possibilities to transfer resistance to other bacterial cells
Every application of antibiotics promotes the development of resistance (Robert Koch Institute, 2019). A short-term use or an application at low dosage give the bacteria a better chance to adapt.
Preventing a disease is always better than its treatment!
But how to do it? Preventing the conditions that favor the proliferation of Clostridium perfringens and strengthening the host’s immune response lowers the probability of disease. Besides eliminating the predisposing factors, the main targets are:
Balance of the gut flora
Optimization of gut function and integrity
Maintenance of immunity
1. Biosecurity is of the highest importance!
There is evidence that most Clostridium strains isolated from birds suffering from Necrotic Enteritis could induce the disease experimentally, while strains isolated from healthy birds cannot. This confirms that only specific strains are problematic (Ducatelle and Van Immerseel, 2010).
So, it’s of highest importance to avoid introducing these pathogenic strains to the farm.
Strict biosecurity measures!
Separate clothing, boots, and hand washing/disinfecting facilities in each poultry house
More than 14 days of down time between flocks
2. Specific measures against coccidiosis
Vaccination
According to parasitologists, 7 to 9 Eimeria species are found in chickens, and they do not cross-protect against each other. An effective vaccination must contain sporulated oocysts of the most critical pathogenic Eimeria species (E. acervulina, E. maxima, E. tenella, E. necatrix, and E. brunetti). The more species contained in the vaccine, the better. However, if not applied the correct way, vaccines can be ineffective or cause reactions in the birds that might lead to NE (Mitchell, 2017).
Anticoccidials
Alternate use of chemicals (synthetic compounds) and ionophores (polyether antibiotics) with different modes of action is important to avoid development of resistance.
Ionophores have a specific mode of action and kill oocysts before they are able to infect birds. Being very small, ionophore molecules can be taken up and diffused into the outer membrane of the sporozoite. There, it decreases the concentration gradient leading to an accumulation of water within the sporozoite causing its bursting.
3. Diet – favorable for the birds but not for clostridium!
Minimizing non-starch polysaccharides (NSPs) in cereals
To prevent a “feeding” of Clostridium perfringens, high content of water-soluble but indigestible NSPs such as wheat, wheat by-products, and barley should be avoided or at least minimized. Additionally, xylanases should be included in the feed formulation to reduce the deleterious effects of NSPs and improve feed energy utilization. Instead of these cereals, maize could be included in the diet. It is considered a perfect ingredient in broiler diets due to its high energy content and high nutrient availability.
Formulating low protein diets/diets with highly digestible amino acids
Feeding low-protein diets supplemented with crystalline amino acids might be beneficial to reduce the risk of Necrotic Enteritis (Dahiya et al., 2007). To improve protein digestibility and therefore reduce the proliferation of C. perfringens, proteases may be added to the feed.
Avoiding/Minimizing animal fats in the diet
Animal fats tend to increase the counts of Clostridium perfringens; thus, they should be replaced by vegetable fat sources.
Feed form is decisive
In terms of feed form, Engberg et al. (2002) found that birds fed pellets showed a reduced number of Clostridium perfringens in the caeca and the rectum than mash-fed birds. Branton and coworkers (1987) reported a lower mortality by feeding roller-milled (coarsely ground) than hammer-milled feed.
4. Additives
Additives can be used either to prevent the proliferation of Clostridium perfringens or to change the environmental conditions in a way that proliferation of C. perfringens is prevented.
1. Probiotics directly support the balance of the microbiome
These live microbial supplements can be used to help to establish, maintain or re-establish the intestinal microflora.
Mode of action:
They compete with pathogenic bacteria for substrates and attachment sites and produce antimicrobial substances inhibiting the growth of pathogenic bacteria (Gillor et al., 2008). They bind and neutralize enterotoxins (Mathipa and Thantsha, 2017) andpromote immune function of the host (Yang et al., 2012)
2. Prebiotics indirectly promote the microbiome
These feed ingredients serve as substrates to promote beneficial bacteria in the intestine.
Mode of action:
D-mannose or fructose, starches non-digestible by birds, selectively stimulate the growth and the activity of the “good” gut flora. Fructooligosaccharides decrease C. perfringens and E. coli in the gut and increase the diversity of Lactobacillus Spp. (Kim et al., 2011). Galactooligosaccharides, in combination with a B. lactis-based probiotic, have been reported to selectively promote the proliferation of Bifidobacterium Spp. (Jung et al., 2008).
3. Organic acids support gut health
Organic acids are often used in animal diets to improve intestinal health.
Mode of action:
A decreased pH promotes beneficial bacteria. Caprylic acid suppresses C. perfringens but also Salmonella spp. by inhibiting their utilization of glucose (Skrivanova et al., 2006). Lauric, citric, oleic, and linoleic acid, as well as medium-chain fatty acids (C8-C14), impede the growth of C. perfringens.
A trial with different organic acid products showed high efficacy for Acidomix AFG and Acidomix AFL against Clostridium perfringens as well as against Salmonella enterica. For the test, 50 µl solution containing different microorganisms (reference strains of S. enterica and C. perfringens; conc. 105 CFU/ml) together with 50 µl of increasing concentrations of various organic acids/organic acid products (Acidomix) were pipetted into microdilution plates. After the respective incubation, the MICs of every organic acid/organic acid product were calculated.
Figure 5 shows the minimum inhibiting concentrations (MIC). For Acidomix AFL and AFG, lower concentrations than for fumaric, lactic, and propionic acid were needed to inhibit the growth of Salmonella enterica and Clostridium perfringens.
Figure 5: Minimal inhibiting concentrations of Acidomix AFL and Acidomix AFG against Salmonella enterica and Clostridium perfringens
Phytomolecules : different types are available against NE
Phytomolecules, also known as secondary plant compounds, have been used against pathogens for centuries. In general, two subgroups of these substances are known as effective against Clostridium perfringens: Tannins and Essential Oils.
Tannins
Many studies have shown the efficacy of tannins against different pathogens such as helminths, Eimeria spp., viruses, and bacteria. Extracts from the chestnut and quebracho trees are effective not only against C. perfringens but also its toxins (Elizando et al., 2010). Tannins act against Eimeria spp. (Cejas et al., 2011) and Salmonella Sp., two predisposing factors for NE.
A trial was conducted with Pretect D, a product based on tannins and saponins, to show its efficacy against coccidia, one of the predisposing factors of NE. For the 35-day study conducted at a commercial research facility in the US, 1800 one-day-old Cobb 500 broilers were divided into four groups of 450 birds each (with 9 replicates & 50 birds per replicate). They all received the standard feed of the farm (Starter D0-D21, Grower D22-D35).
The challenge was given in the form of a freshly prepared mixed inoculum with E. acervulina (100,000 oocysts/ bird), E. maxima (50,000 oocysts/ bird), and E. tenella (75,000 oocysts/ bird). The inoculum was mixed into the feed in the base of each pen’s tube feeder.
The oocyst count per gram of feces (OPG) was done on D21, D27 & D35. The cocci lesion scoring (CLS) was done on D27 following Johnson and Reid (1970) with 0=normal; 4=most
Group
Challenge
Additive
Non-challenged Control (NC)
No
No
Challenged Control (CC)
Yes
No
CC + Ionophore
Yes
Ionophore@60ppm
CC + Pretect D
Yes
Pretect D@500ppm
The trial showed that, due to Pretect D, the lesion score showed a lower value indicating that lesions could be reduced or were less severe, which can be seen in figure 6:
Figure 6: Average lesion score
Essential Oils
Their hydrophobic characteristic enables them to interact with the lipids of the membrane of C. perfringens. They can incorporate into the bacterial membrane and disrupt its integrity, increasing the permeability of the cell membrane for ions and other small molecules such as ATP and leading to the decrease of the electrochemical gradient above the cell membrane and the loss of the cell’s energy equivalents. Besides their direct effect on Clostridium spp., many phytomolecules improve gut health and help prevent the proliferation of Clostridium spp. And, therefore, Necrotic Enteritis.
An In vitro-trial shows Ventar D reducing Clostridia and sparing the beneficial lactobacilli. In this trial, the bacteria (Clostridium perfringens, Lactobacillus agilis S73, and Lactobacillus plantarum) were cultured under favorable conditions (RCM, 37°C, anaerobe for Clostridium perfringens, and MRS, 37°C, 5 % CO2 for Lactobacilli) and exposed to different concentrations of Ventar D (0 µg/ml – control, 500 µg/ml, 750 µg/ml, and 1000 µg/ml).
The results of the trial with Clostridium perfringens are shown in figure 7.
Figure 7: Different concentrations of Ventar D added to Clostridium perfringens cultures
Here, a significant reduction of colonies could already be observed at a concentration of 500 µg/ml of Ventar D. With 750 µg/ml, only a few colonies remained, and at a Ventar D concentration of 1000 µg/ml, Clostridium perfringens didn’t grow anymore.
In contrast, the Lactobacilli showed a different picture: only at the higher concentration (1250 µg/ml of Ventar D) did Lactobacillus plantarum and Lactobacillus agilis S73 show a slight growth reduction (figure 8).
Figure 8: Lactobacillus plantarum exposed to 0 (left) and 1250 µg/ml (right) of Ventar D
In vivo-trial in poultry shows that phytomolecules reduce gut lesions
The study was conducted at Southern Poultry Feed & Research, Athens, GA (USA), over 42 days. It included in total 880 Cobb 500 broilers across 2 trial groups, with 11 repetitions per trial set-up and 40 animals per replicate floor pen. All animals received routine vaccinations at the hatchery and were healthy when starting the trial.
Control group
Built-up litter (no additive)
Ventar D group
Built-up litter + Ventar D, 100 g/MT of feed
All birds received standard feed, fed as crumbles/pellets ad libitum. Feed intake by pen was recorded per feeding phase for starter (D21), grower (D35), and finisher feed (D42). Bird weights were recorded at study initiation, on D21, D35, and D42. On D21 and D35, three birds per pen were sacrificed. The GIT was scored for necrotic enteritis lesions; figures 9 and 10 show the results.
Figures 9 and 10: Lesion score on days 21 and 35
Already on day 21, the birds of the Ventar D group showed a less impacted gut mucosa, indicated by a lower lesion score. Lesions were reduced in both groups until day 35; however, the value of the Ventar D group was still better.
A less impacted gut has a higher digestion and absorption capacity, which results in better performance (FCR and weight gain) and lower mortality (figures 11-14).
Figures 11-14: Performance data of a control group compared with birds supplemented with Ventar D
The two trials show that Ventar D allows the poultry producer to proactively strengthen broilers’ gut health by controlling Clostridia perfringens and promoting/saving beneficial bacteria such as lactobacilli. The effects of the reduction of Clostridia can be seen in vivo in a lower lesion score and better performance.
They bind mycotoxins and, therefore, reduce or prevent damage to the intestinal wall so that the preconditions for Clostridium spp. proliferation are not generated.
Additionally, binding toxins produced by Clostridium perfringens can reduce the occurrence or severity of lesions: Alpha-toxin, phospholipase C, hydrolyses membrane phospholipids and damages erythrocytes, leucocytes, myocytes, and endothelial cells and causes their lipolysis (Songer, 1996), leading to necrosis and tissue damage.
Binding NetB toxin, the key virulence factor, could reduce the severity of Necrotic Enteritis.
A trial was conducted in a laboratory in Valladolid/Spain to show the high binding capacity of Solis Plus 2.0. All tests were carried out as duplicates and using a standard liquid chromatography/mass spectrometry (LC/MS/MS) quantification. Interpretation and data analysis were carried out with the corresponding software. Toxin concentrations, anti-mycotoxin agent application rates, and pH levels were set as follows:
Mycotoxin
Challenge Level
Challenge (ppb)
Solis Plus 2.0 inclusion rate
Assay time
Aflatoxin
Low
150
0.2%
30 min.
High
1500
Fumonisin
Low
500
High
5000
Ochratoxin
Low
150
High
1500
The results are shown in figure 15:
Figure 15: Adsorption capacity of Solis Plus for relevant mycotoxins
Under acidic conditions (pH 3), Solis Plus 2.0 effectively adsorbs the three tested mycotoxins at low and high contamination levels:
Aflatoxin: 150 ppb -100 %; 1500 ppb – 98 %
Fumonisin: 500 ppb – 87%; 5000 ppb – 86 %
Ochratoxin: 150 ppb – more than 43 %; 1500 ppb – 52 %.
By binding harmful toxins and preventing their negative impact on the gut, toxin binders can also be a tool to reduce necrotic enteritis.
NE can be controlled – even in an antibiotic-free era
The ever-growing trend of reduced antibiotic and ionophore use increases the incidence of Necrotic Enteritis in poultry production. Especially the subclinical form, which generally goes unnoticed, results in poor feed efficiency and is a major cause of financial losses to poultry producers.
Maintaining optimum gut health is key to preventing the occurrence of Necrotic Enteritis. In the era of antibiotic-free poultry production, alternatives acting against the pathogenic bacterium and also against its predisposing factors must be considered to control this devastating disease. The industry already provides solutions like phytomolecules-based products or toxin binders to support the animals.
References:
Annett, C.B., J. R. Viste, M. Chirino-Trejo, H. L. Classen, D. M. Middleton, and E. Simko. “Necrotic enteritis: effect of barley, wheat and corn diets on proliferation of Clostridium perfringens type A.” Avian Pathology 31 (2002): 599– 602. https://doi.org/10.1080/0307945021000024544
Antonissen G, F. Van Immerseel, F. Pasmans, R. Ducatelle, F. Haesebrouck, L. Timbermont, M. Verlinden, G.P.J. Janssens, V. Eeckhaut, M. Eeckhout, S. De Saeger, S. Hessenberger, A. Martel, and S. Croubels. “The mycotoxin deoxynivalenol predisposes for the development of Clostridium perfringens-Induced necrotic enteritis in broiler chickens. PLoS ONE 9 no. 9 (2014): e108775. https://doi.org/10.1371/journal.pone.0108775
Antonissen, G., V. Eeckhaut, K. Van Driessche, L. Onrust , F. Haesebrouck, R. Ducatelle, R.J. Moore, and F. Van Immerseel. “Microbial Shifts Associated With Necrotic Enteritis.” Avian Pathol. 45 no. 3 (2016): 308-312. https://doi.org/10.1080/03079457.2016.1152625
Branton, S.L., F.N. Reece, and W.M. Hagler. “Influence of a wheat diet on mortality of broiler chickens associated with necrotic enteritis.” Poultry Sci. 66 (1987): 1326-1330. https://doi.org/10.3382/ps.0661326
Cejas, E., S. Pinto, F. Prosdócimo, M. Batalle, H. Barrios, G. Tellez, and M. De Franceschi. “Evaluation of quebracho red wood (Schinopsis lorentzii) polyphenols vegetable extract for the reduction of coccidiosis in broiler chicks.” International Journal of Poultry Science 10 no. 5 (2011): 344–349. https://doi.org/10.3923/ijps.2011.344.349
Collier, C.T., C.L. Hofacre, A.M. Payne, D.B. Anderson, P. Kaiser, R.I. Mackie, and H.R. Gaskins. “Coccidia-induced mucogenesis promotes the onset of necrotic enteritis by supporting Clostridium perfringens growth.” Veterinary Immunology and Immunopathology 122 (2008):104–115.
Dahiya, J.P., D. Hoehler, A.G. Van Kessel, and M.D. Drew. “Effect of different dietary methionine sources on intestinal microbial populations in broiler chickens.” Poultry Science 86 (2007):2358–2366
Dahiya, J.P., D. Hoehler, D.C. Wilkie, A.G. van Kessel, and M.D. Drew. “Dietary glycine concentration affects intestinal Clostridium perfringens and Lactobacilli populations in broiler chickens.” Poultry Science 84 no.12 (2005):1875-85. https://doi.org/10.1093/ps/84.12.1875
Diaz Carrasco, J.M., L.M. Redondo, E.A. Redondo, J.E. Dominguez, A.P. Chacana, and M.E. Fernandez Miyakawa. “Use of plant extracts as an effective manner to control Clostridium perfringens induced necrotic enteritis in poultry.” BioMed Research International (2016): Article ID 3278359. https://dx.doi.org/10.1155/2016/3278359
Ducatelle, R. and F. van Immerseel. “Necrotic enteritis: emerging problem in broilers.” WATTAgNet.com – Poultry Health and Disease (April 9, 2010).
Elizondo, A.M., E.C. Mercado, B.C. Rabinovitz, and M.E. Fernandez-Miyakawa. “Effect of tannins on the in vitro growth of Clostridium perfringens.” Veterinary Microbiology 145 no. 3-4 (2010): 308–314. https://doi.org/10.1016/j.vetmic.2010.04.003
Engberg, R.M., M.S. Hedemann, and B.B. Jensen. “The influence of grinding and pelleting of feed on the microbial composition and activity in the digestive tract of broiler chickens.” · British Poultry Science 43 no. 4 (2002):569-579. https://doi.org/10.1080/0007166022000004480
Fischetti, V.A. “Bacteriophage endolysins: A novel anti-infective to control Gram-positive pathogens.” J Med Microbiol. 300 no. 6 (2010): 357–362. https://doi.org/10.1016/j.ijmm.2010.04.002
Gillor, O., A. Etzion and M.A. Riley. “The dual role of bacteriocins as anti- and probiotics.” Appl Microbiol Biotechnol. 81 no. 4 (2008): 591–606. https://doi.org/10.1007/s00253-008-1726-5
Hofacre, C.L., J.A. Smith, and G.F. Mathis. “Invited Review. An optimist’s view on limiting necrotic enteritis and maintaining broiler gut health and performance in today’s marketing, food safety, and regulatory climate.” Poultry Science 97 (2018):1929–1933. https://dx.doi.org/10.3382/ps/pey082
Jung, S.J., R. Houde, B. Baurhoo, X. Zhao, and B. H. Lee. “Effects of galacto-oligosaccharides and a bifidobacteria lactis-based probiotic strain on the growth performance and fecal microflora of broiler chickens.” Poultry Science 87 (2008):1694–1699. https://doi.org/10.3382/ps.2007-00489
Kaldhusdal and Skjerve. “Association between cereal contents in the diet and incidence of necrotic enteritis in broiler chickens in Norway.” Preventive Veterinary Medicine 28 (1996):1-16. https://doi.org/10.1016/0167-5877(96)01021-5
Keyburn, A. L., S. A. Sheedy, M. E. Ford, M. M. Williamson, M. M. Awad, J. I. Rood, and R. J. Moore. “Alpha-toxin of Clostridium perfringens is not an essential virulence factor in necrotic enteritis in chickens.” Infect. Immun. 74 (2006): 6496–6500. https://doi.org/10.1128/IAI.00806-06
Keyburn, A.L., J.D. Boyce, P. Vaz, T.L. Bannam, M.E. Ford, D. Parker, A. Di Rubbo, J.I. Rood, and R.J. Moore. “NetB, a new toxin that is associated with avian necrotic enteritis caused by Clostridium perfringens.” PLoS Pathog 4 no. 2, e26 (2008): 0001-0011. https://doi.org/10.1371/journal.ppat.0040026
Kim, G.-B., Y. M. Seo , C. H. Kim , and I. K. Paik. “Effect of dietary prebiotic supplementation on the performance, intestinal microflora, and immune response of broilers.” Poultry Science 90 (2011):75–82. https://doi.org/10.3382/ps.2010-00732
Knap, I., B. Lund, A. B. Kehlet, C. Hofacre, and G. Mathis. “Bacillus licheniformis prevents necrotic enteritis in broiler chickens.” Avian Diseases 54 no. 2 (2010):931-935. https://doi.org/10.1637/9106-101509-ResNote.1
Knarreborg, A., M.A. Simon, R.M. Engberg, B.B. Jensen, and G.W. Tannock. “Effects of Dietary Fat Source and Subtherapeutic Levels of Antibioticon the Bacterial Community in the Ileum of Broiler Chickensat Various Ages.” Applied and Environmental Microbiology 68 no. 12 (2002): 5918-5924. https://doi.org/0.1128/AEM.68.12.5918–5924.2002
Kocher, A. and M. Choct. “Improving broiler chicken performance. The efficacy of organic acids, prebiotics and enzymes in controlling necrotic enteritis.” Australian Government-Rural Industries Research and Development Corporation. Publ. no. 08/149 (2008).
Kondo, F. “In vitro lecithinase activity and sensitivity to 22 antimicrobial agents of Clostridium perfringens isolated from necrotic enteritis of broiler chickens.” Research in veterinary Science 45 (1988): 337-340. https://doi.org/10.1016/S0034-5288(18)30961-5
Kubena, L.F., J.A. Byrd, C.R. Young, and D.E. Corrier. “Effects of tannic acid on cecal volatile fatty acids and susceptibility to Salmonella typhimurium colonization in broiler chicks.” Poultry Science 80, no. 9, pp. 1293–1298, 2001. https://doi.org/10.1093/ps/80.9.1293
M’Sadeq S.A., Shubiao Wu, Robert A. Swick, Mingan Choct. “Towards the control of necrotic enteritis in broiler chickens with in-feed antibiotics phasing-out worldwide.” Animal Nutrition 1 (2015): 1-11. https://dx.doi.org/10.1016/j.aninu.2015.02.004
Mathipa, M.G. and M.S. Thantsha. “Probiotic engineering: towards development of robust probiotic strains with enhanced functional properties and for targeted control of enteric pathogens.” Gut Pathog. 9 no. 28 (2017). https://doi.org/10.1186/s13099-017-0178-9
McDevitt, R.M., J.D. Brooker, T. Acamovic, and N.H.C. Sparks. “Necrotic enteritis, a continuing challenge for the poultry industry.” World’s Poultry Science Journal 62; World’s Poultry Science Association (June 2006). https://doi.org/10.1079/WPS200593
Miller, R.W., J. Skinner, A. Sulakvelidze, G.F. Mathis, and C.L. Hofacre. “Bacteriophage therapy for control of Necrotic Enteritis of broiler chickens experimentally infected with Clostridium perfringens.” Avian Diseases 54 no. 1 (2010): 33-40. https://doi.org/10.1637/8953-060509-Reg.1
Mitsch, P., K. Zitterl-Eglseer, B. Köhler, C. Gabler, R. Losa, and I. Zimpernik. “The Effect of Two Different Blends of Essential Oil Components on the Proliferation of Clostridium perfringens in the Intestines of Broiler Chickens.” Poultry Science 83 (2004):669–675. https://doi.org/10.1093/ps/83.4.669
Olkowski, A.A., C. Wojnarowicz, M. Chirino-Trejo, B. Laarveld, and G. Sawicki. “Sub-clinical necrotic enteritis in broiler chickens: Novel etiological consideration based on ultra-structural and molecular changes in the intestinal tissue.” Veterinary Science 85 (2008): 543–553. https://doi.org/10.1016/j.rvsc.2008.02.007
Pan, D. and Z. Yu. “Intestinal microbiome of poultry and its interaction with host and diet.” Gut Microbes 5 no. 1 (2014): 108–119. https://dx.doi.org/10.4161/gmic.26945
Rougière, N. and B. Carré. “Comparison of gastrointestinal transit times between chickens from D + and D- genetic lines selected for divergent digestion efficiency.” Animal 4 no. 11 (2010): 1861-1872. https://doi.org/10.1017/S1751731110001266
Santos, F.B.O., B.W. Sheldon, A.A. Santos Jr., and P.R. Ferket.” Influence of housing system, grain type, and particle size on Salmonella colonization and shedding of broilers fed triticale or corn-soybean meal diets.” Poultry Science 87 (2008): 405-420. https://dx.doi.org/10.3382/ps.2006-00417
Schiavone, A. , K. Guo, S. Tassone, L .Gasco, E. Hernandez, R. Denti, and I. Zoccarato. “Effects of a Natural Extract of Chestnut Wood on Digestibility, Performance Traits, and Nitrogen Balance of Broiler Chicks.” Poult Sci. 87 no. 3 (2008): 521-527. https://doi.org/10.3382/ps.2007-00113
Singh, Y., V. Ravindran, T.J. Wester, A.L. Molan, and G. Ravindran. “Influence of feeding coarse corn on performance, nutrient utilization, digestive tract measurements, carcass characteristics, and cecal microflora counts of broilers.” Poultry Science 93 (2014): 607–616. https://dx.doi.org/10.3382/ps.2013-03542
Skrivanova, E., M. Marounek, V. Benda, and P. Brezina. “Susceptibility of Escherichia coli, Salmonella sp. and Clostridium perfringens to organic acids and monolaurin.” Veterinarni Medicina 51 no. 3 (2006): 81–88. https://doi.org/10.17221/5524-VETMED
Stanley D., Wu S.-B., Rodgers N., Swick R.A., and Moore R.J. “Differential Responses of Cecal Microbiota to Fishmeal, Eimeria and Clostridium perfringens in a Necrotic Enteritis Challenge Model in Chickens.” PLoS ONE 9 no. 8 (2014): e104739. https://doi.org/10.1371/journal.pone.0104739
Tan, L., D. Rong, Y. Yang, and B. Zhang. “Effect of Oxidized Soybean Oils on Oxidative Status and Intestinal Barrier Function in Broiler Chickens.” Brazilian Journal of Poultry Science 18 no. 2 (2018): 333-342. http://dx.doi.org/10.1590/1806-9061-2017-0610
Tan, L., D. Rong, Y. Yang, and B. Zhang. “The Effect of Oxidized Fish Oils on Growth Performance, Oxidative Status, and Intestinal Barrier Function in Broiler Chickens.” J. Appl. Poult. Res. 28 (2019): 31-41. http://dx.doi.org/10.3382/japr/pfy013
Timbermont L., A. Lanckriet, J. Dewulf, N. Nollet, K. Schwarzer, F. Haesebrouck, R. Ducatelle, and F. Van Immerseel. “Control of Clostridium perfringens-induced necrotic enteritis in broilers by target-released butyric acid, fatty acids and essential oils.” Avian Pathol. 39 no. 2 (2010): 117-21. https://doi.org/10.1080/03079451003610586
Tsiouris, V. “Poultry management: a useful tool for the control of necrotic enteritis in poultry.” Avian Pathol. 45 no. 3 (2016):323-325. https://doi.org/10.1080/03079457.2016.1154502
Van der Most, P.J., B. de Jong, H.K. Parmentier and S. Verhulst. “Trade-off between growth and immune function: a meta-analysis of selection experiments.” Functional Ecology 25 (2011): 74-80. https://doi.org/0.1111/j.1365-2435.2010.01800.x
Van der Sluis, W. “Clostridial enteritis is an often underestimated problem.” Worlds Poult. Sci. J. 16 (2000):42–43.
Van Immerseel, F., J. De Buck, F. Pasmans, G. Huyghebaert, F. Haesebrouck, and R. Ducatelle. “Clostridium perfringens in poultry: an emerging threat of animal and public health.” Avian Pathology 33 (2004): 537-549. https://doi.org/10.1080/03079450400013162
Van Immerseel, F., J.I. Rood, R.J. Moore, and R.W. Titball. “Rethinking our understanding of the pathogenesis of necrotic enteritis in chickens.” Trends in Microbiology 17 no. 1 (2008):32-36. https://doi.org/10.1016/j.tim.2008.09.005
Wade, B., A.L. Keyburn, T. Seemann, J.I. Rood, and R.J. Moore. “Binding of Clostridium perfringens to collagen correlates with the ability to cause necrotic enteritis in chickens.” Veterinary Microbiology 180 no. 3–4 (2015): 299-303. https://doi.org/10.1016/j.vetmic.2015.09.019
Williams, R.B. “Intercurrent coccidiosis and necrotic enteritis of chickens: rational, integrated disease management by maintenance of gut integrity.” Avian Pathology 34 no. 3 (2005):159-180. https://doi.org/10.1080/03079450500112195
Yang , C.M., G.T. Cao, P.R. Ferket, T.T. Liu, L. Zhou, L. Zhang, Y.P. Xiao, and A. G. Chen. “Effects of probiotic, Clostridium butyricum, on growth performance, immune function, and cecal microflora in broiler chickens.” Poultry Science 91 (2012): 2121–2129. https://dx.doi.org/10.3382/ps.2011-02131