Organic acids: How the mode of action delivers benefits | INFOGRAPHIC

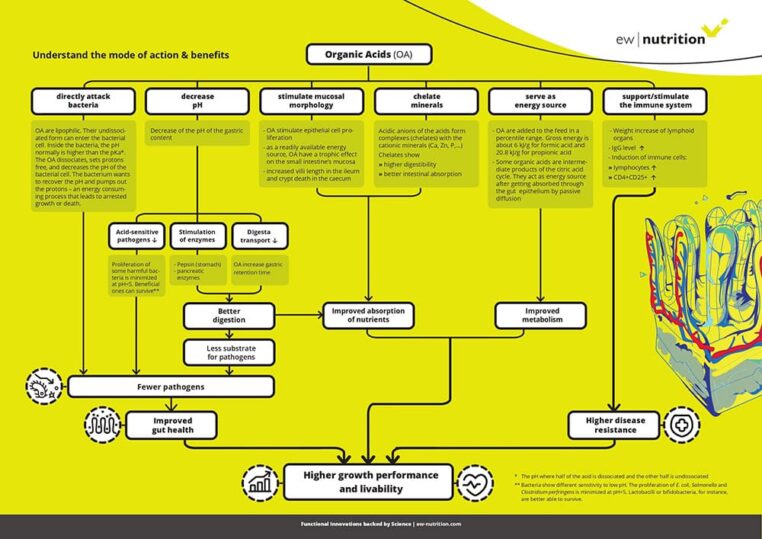
Download this comprehensive A3 infographic to understand how organic acids can benefit your operation. For further questions or feedback, we’ll be happy to hear from you.


Download this comprehensive A3 infographic to understand how organic acids can benefit your operation. For further questions or feedback, we’ll be happy to hear from you.

By Technical Team, EW Nutrition
A storm has been brewing.
Even before the invasion of Ukraine in late February, global growth was expected to trend significantly downward, from 5.5-5.9% in 2021 to 4.1-4.4% in 2022 and 3.2% in 2023. The causes are similar across industries:
In early 2022, this “perfect storm” quickly stifled the moderate optimism of Q4 2021. Of course, the worst was yet to come.
With the ongoing crisis in Eastern Europe, economic perspectives are tilting down to a new level of uncertainty. The new variables now thrown into the mix are crude oil and natural gas prices, as well as added concerns over other raw materials coming out of Russia and Ukraine.
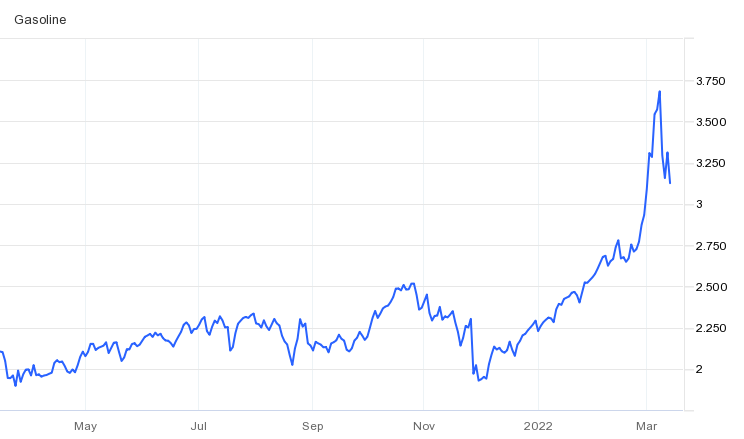
Source: tradingeconomics.com, March 2022
Russia accounts for 25% of the global natural gas market and 11% of the crude oil market. It is also the largest wheat exporter (China and India are still the largest producers, but Russia exports appreciably more). Together with Ukraine, also a powerhouse of agricultural exports, the two now enemies account for 29% of international annual wheat sales.
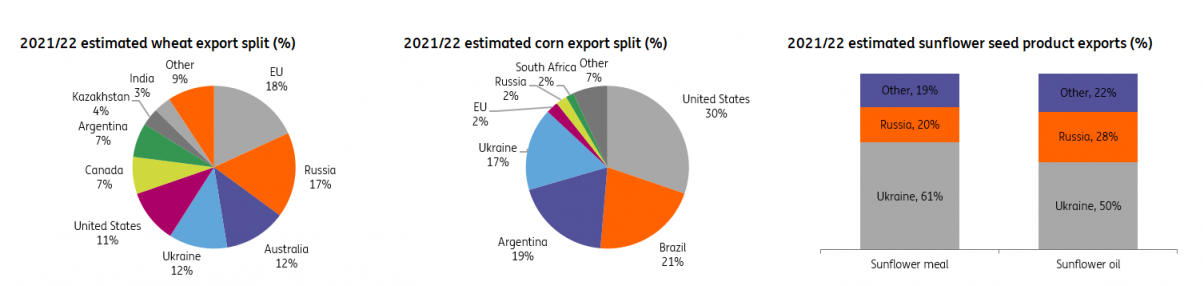
Source: ING, March 2022
Wheat prices were already nearly double the five-year average shortly before the invasion; after February 24, they rose by another 30%. Today we are at a staggering 53% increase in wheat prices in just the last few months. We are at a 14-year peak. And the countries that import the most from Russia and Ukraine (such as Egypt or Indonesia) will bear the brunt of this crisis.
Together, Russia and Ukraine’s exports account for 12% of the world’s traded calories. The two countries account for almost 30 percent of global wheat exports, almost 20 percent of corn exports, and more than 80 percent of the world supply of sunflower oil. However, the compounded effect of embargo and devastation in the two countries will surely exert tremendous influence on the global economic outlook for years to come.
Agriculture was already hurting before February 24th. Poor harvests caused by extreme weather conditions, continued losses along the production chain, supply chain issues, and abnormal pandemic buying patterns combined to sink global wheat stocks one third lower than the five-year average. Reserves, in other words, are low – and will be significantly lower.
We need to be realistic about the coming months and years. Corn (where Ukraine accounts for 13% of global exports) and wheat will be severely hit by the war and its aftermath. This will compound all the pre-existing factors (transportation costs, supply chain slowdown, continuing weather disruptions, energy costs), none of which will trend down. Fertilizer prices have also gone up exponentially, and Russia – the largest exporter – has banned fertilizer exports at the beginning of March. The effects will be ultimately reflected in the cost of raw materials.
Ukraine and Russia have all but banned grains exports – either for security reasons or to protect internal needs. On top of this, the last harvests collected in Ukraine are now sitting in bins where ventilation and temperature controls have been affected by power cuts.
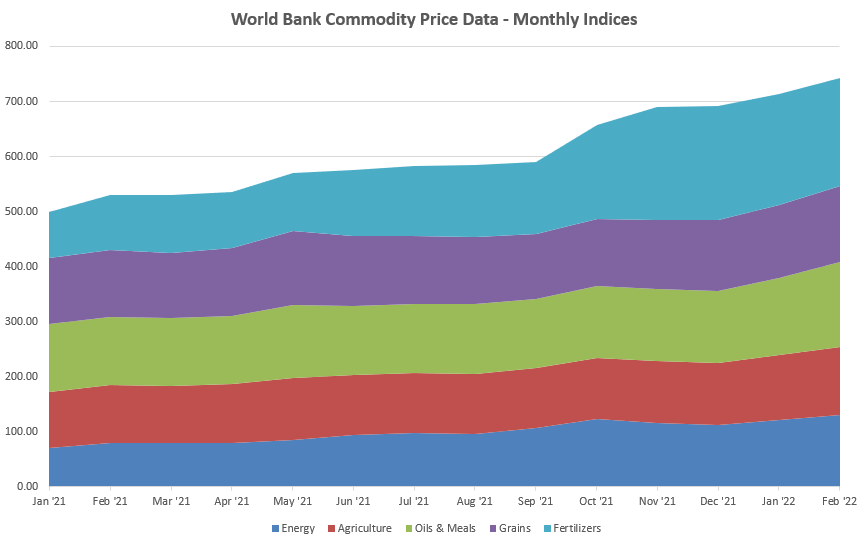
Source: World Bank, March 2022
At the end of February, World Bank data already showed upward movement for nearly all categories; whatever was not trending up at that time is catching up fast. The last time things looked like this, experts warn, was in 2008-2009 – and social unrest followed around the world, to serious global consequences.
However, the perspective is not catastrophic and there is room to conserve profitability. The essential is to intervene with fast, targeted action that favors smart optimization, localization, and long-term planning.
Most feed producers will be caught in the middle of all rising costs, from raw materials to transport and energy. Where, then, can they look for shelter when the storm hits?
One of the first things feed producers will focus on will be cutting down feed costs. At this point, it is essential that this basic optimization does not impact animal health and performance. Here is what should be kept in mind.
Whatever raw materials you choose to use, minimizing losses and maintaining quality should be the first step. Losses caused by storage are often the easiest to mitigate.
Quick intervention #1: Use mold inhibitors and mitigate the impact of mycotoxins
Freight costs will continue to cause pressure on transported raw materials, driving producers to local/regional options. When you replace one feed ingredient with a cheaper one, the first effects will be on the active principle and on the digestibility of the feed. Often something you are taking out of the diet cannot be replaced 1:1.
Quick intervention #2: Maximize the use of enzymes to ensure high feed digestibility; for poultry, pigments can replace corn-derived coloration (to control color variability)
Adjusting the feed composition doesn’t only have effects on paper.
Even if you choose the best replacements, adjust the balance, compensate for loss of digestibility and optimize everything in every possible way, one thing remains:
The animal receives a new diet.
New diets are textbook stressors. But sometimes the nutritionist or the producer is so stressed that it is easy to overlook the stress placed inside the animal. Since animal efficiency is key for productivity, it is essential that the effects of diet stress are mitigated for the animal.
Quick intervention #3: Precautionary use of gut-health mitigating additives; also consider palatable feed materials and taste enhancers
To optimize costs on the production floor, there are three essential areas where feed producers can act:
To answer these challenges, there are solutions that can operate individually. More importantly in such times, there are products that can impact all three areas without negatively influencing the quality of output. One such solution, for instance, can decrease energy costs, increase throughput and pellet quality, and reduce fines.
Quick intervention #4: Choose a solution that satisfies 3/3 of your issues
Climate change will continue to wreak havoc on the predictability of harvests. Freight costs are projected to keep rising. And the costs of war and (hopefully) reconstruction will take a toll on the cost of living and cost of doing business around the world, for years to come.
In the storm that has already started, it is unwise to take shelter for a while and hope for good weather soon. Cutting down on ingredients here and additives there won’t keep profitability high in the long run. Feed producers must look at all aspects – from feed storage and composition to process improvement – and consider holistic measures that protect animals and profitability at the same time.

By Technical Team, EW Nutrition
Contamination with multiple mycotoxins is the rule for animal feeds, rather than the exception. Trial data shows that producers can prevent negative effects on animal health and performance by using high-performing toxin binders.

Mycotoxins pose an exceptional challenge for feed and animal producers. Generated by common molds, they occur in a great variety and numbers. Difficult to diagnose, mycotoxicosis in farm animals shows in a range of acute and chronic symptoms: decreased performance, feed refusal, poor feed conversion, reduced body weight gain, immune suppression, reproductive disorders, and residues in animal food products.
Regulatory thresholds for permissible mycotoxin levels in feed are derived from toxicological data on the effects of exposure of a certain species, at a certain production stage, to a single mycotoxin. This makes practical sense: while aflatoxins are carcinogens, fumonisins attack the pulmonary system in swine, for example. Mycotoxins also affect poultry in a different way than cattle, and broilers in a different way than breeders or laying hens, to mention more cases.
The problem is that, in reality, individual mycotoxin challenges are the exception. Animal diets are usually contaminated by multiple mycotoxins at the same time (Monbaliu et al., 2010; Pierron et al., 2016). Since 2014, EW Nutrition has conducted more than 50,000 mycotoxin tests on both raw material and finished feeds samples, across the globe. 85% of these samples were contaminated with more than one mycotoxin and one third positive for four or more mycotoxins.
The concurrent appearance of mycotoxins in feed can be explained as follows: each mold species has the capacity to produce several mycotoxins simultaneously. Each species, in turn, may infest several raw materials, leaving behind one or more toxic residue. In the end, a complete diet is made up of various raw materials with individual mycotoxin loads, resulting in a multitude of toxic challenges for the animals.
If animals were exposed to only one mycotoxin at a time, following the regulatory guidelines on maximum challenge levels would usually be enough to keep them safe. However, several studies have shown that the effects of exposure to multiple mycotoxins can differ greatly from the effects observed in animals exposed to a single mycotoxin (Alassane-Kpembi et al., 2015 & 2017). The simultaneous presence of mycotoxins may be more toxic than one would predict based on the known effects of the individual mycotoxins involved. This is because mycotoxins interact with each other. The interactions can be classified into three main different categories: antagonistic, additive, and synergistic (Grenier and Oswald, 2011).

Most of the mycotoxin mixtures lead to additive or synergistic effects. The actual consequences for the animal will depend on its species, age, sex, nutritional status, the dose and duration of exposure as well as environmental factors. What is clear is that mycotoxin interactions pose a significant threat to animal health and critically impede risk assessment.
Given their complex interactions, the toxicity of combinations of mycotoxins cannot merely be predicted based upon their individual toxicities. Mycotoxin risk assessments have to consider that even low levels of mycotoxin combinations can harm animal productivity, health, and welfare. Feed and animal producers need to be aware of which raw materials are likely to be contaminated with which mycotoxins, be able to accurately link them to the risk they pose for the animal and consequently take actions before the problems appear in the field.
Toxin binders that are effective against a broad spectrum of mycotoxins significantly reduce the risks of mycotoxin exposure. In vitro trial data shows that EW Nutrition’s cost-effective toxin-mitigating product Solis Max shows a high mitigation capacity, even at low inclusion rates (Figure 1). Importantly, Solis Max helps to reduce various mycotoxins’ negative effects on performance without any negative effects on nutrient absorption.
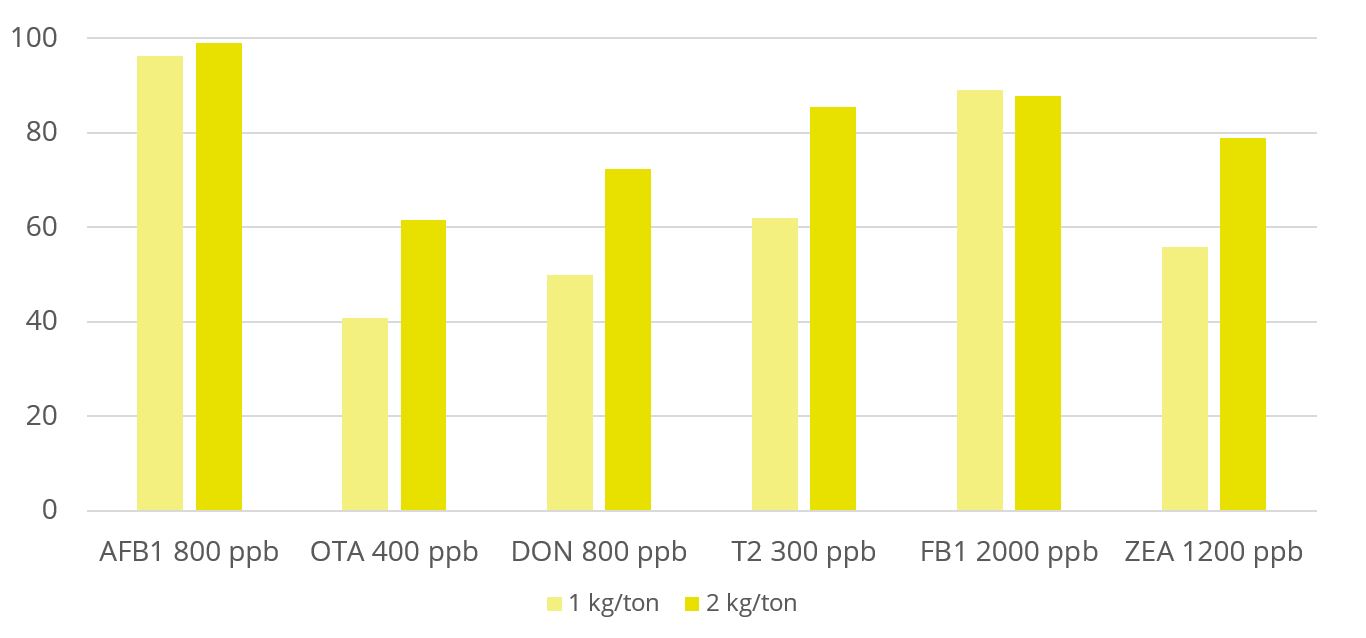
In another trial with 480 Ross 308 broilers, Solis Max 2.0 has demonstrated its ability to support animals coping with multiple mycotoxin challenges. For broilers challenged with 30 ppb AFB1 and 500 ppb OTA, Solis Max 2.0 supplementation resulted in a significantly 16% higher weight gain, an 18.5% higher final weight, and a 22% better FCR than the challenged group (Figure 2), which means a total recovery of the performance when compared with the non-challenged control.

Liver health also improved: after 42 days of mycotoxin exposure, broilers receiving Solis Max 2.0 showed lower AST (-13%) and ALT (-44%) levels compared to the challenged group.
Mycotoxins interactions are the norm, not the exception. Yet, regulatory standards currently only cover the effects of individual mycotoxins, leaving productions exposed to risks of additive and synergistic mycotoxin interactions animals’ health and performance. Luckily, management options are available: Careful risk evaluation explicitly includes the threat of multiple contaminations. And producers can proactively ensure better health, welfare and productivity of their animals by investing in the right toxin mitigation solution for their business.
Alassane-Kpembi, Imourana, Olivier Puel, and Isabelle P. Oswald. “Toxicological Interactions between the Mycotoxins Deoxynivalenol, Nivalenol and Their Acetylated Derivatives in Intestinal Epithelial Cells.” Archives of Toxicology 89, no. 8 (August 2015): 1337–46. https://doi.org/10.1007/s00204-014-1309-4.
Alassane-Kpembi, Imourana, Gerd Schatzmayr, Ionelia Taranu, Daniela Marin, Olivier Puel, and Isabelle Paule Oswald. “Mycotoxins Co-Contamination: Methodological Aspects and Biological Relevance of Combined Toxicity Studies.” Critical Reviews in Food Science and Nutrition 57, no. 16 (November 2017): 3489–3507. https://doi.org/10.1080/10408398.2016.1140632.
Bensassi, Fatma; Gallerne, Cindy; Sharaf el dein, Ossama; Rabeh Hajlaoui, Mohammed; Lemaire, Christophe and Bacha, Hassen. “In vitro investigation of toxicological interactions between the fusariotoxins deoxynivalenol and zearalenone” Toxicon 84 (2014): 1-6. https://doi.org/10.1016/j.toxicon.2014.03.005.
Grenier, B., and I. Oswald. “Mycotoxin Co-Contamination of Food and Feed: Meta-Analysis of Publications Describing Toxicological Interactions.” World Mycotoxin Journal 4, no. 3 (May 5, 2011): 285–313. https://doi.org/10.3920/wmj2011.1281.
Miazzo, R., M.F. Peralta, C. Magnoli, M. Salvano, S. Ferrero, S.M. Chiacchiera, E.C.Q. Carvalho, C.A.R. Rosa, and A. Dalcero. “Efficacy of Sodium Bentonite as a Detoxifier of Broiler Feed Contaminated with Aflatoxin and Fumonisin.” Poultry Science 84, no. 1 (January 2005): 1–8. https://doi.org/10.1093/ps/84.1.1.
Monbaliu, Sofie, Christof Van Poucke, Christ’l Detavernier, Frédéric Dumoulin, Mario Van De Velde, Elke Schoeters, Stefaan Van Dyck, Olga Averkieva, Carlos Van Peteghem, and Sarah De Saeger. “Occurrence of Mycotoxins in Feed as Analyzed by a Multi-Mycotoxin LC-MS/MS Method.” Journal of Agricultural and Food Chemistry 58, no. 1 (2010): 66–71. https://doi.org/10.1021/jf903859z.
Pierron, Alix, Imourana Alassane-Kpembi, and Isabelle P. Oswald. “Impact of Mycotoxin on Immune Response and Consequences for Pig Health.” Animal Nutrition 2, no. 2 (2016): 63–68. https://doi.org/10.1016/j.aninu.2016.03.001.

When you think of a “digitally mature company,” what do you see? The term might conjure up images of tech-savvy millennials in Silicon Valley, but digitally mature companies are all around you—in every industry, in every corner of the globe. They’re enterprises that have embraced digital technology to transform the way they do business and connect with their customers.

So what does a digitally mature company look like? It’s an enterprise that:
However you define it, digital maturity matters—and the research shows a direct correlation between being a digitally mature company and having bottom line success.
The economic benefits of digital maturity are well documented. A McKinsey study of 2,400 companies found that those able to exploit ICT to boost efficiency grew revenue 3.2% faster annually than their peers over a five-year period. Another study looked at more than 180 publicly listed companies from around the world, finding that digitally mature organizations outperformed their less mature competitors across all financial measures. This included a return on invested capital (ROIC) 50% higher than laggards.
In an increasingly digital world, companies that embrace technology are simply becoming more competitive and profitable than those who don’t. And it’s no wonder that digitally mature companies have a competitive edge. According to the 2016 State of Digital Business Transformation study conducted by SAP, organizations that see themselves as digitally mature outperform their peers in terms of net income by 20%. In fact, those with advanced digital capabilities enjoy a 42% net income margin compared with 18% for those without any digital capability.
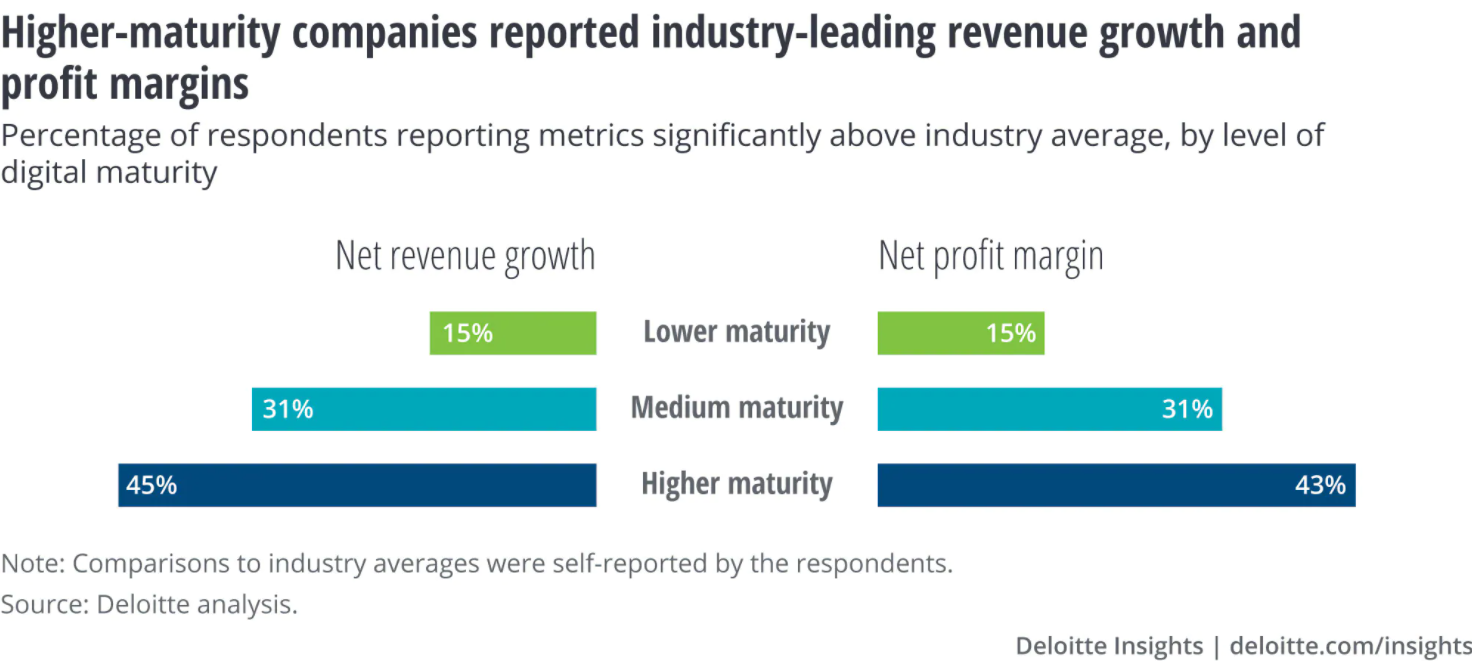
According to PwC’s Annual Global CEO Survey, digitally mature companies are out-performing their less digitally mature peers by 53 percent when it comes to revenue growth over the past three years.
Research shows that digitally mature companies are significantly more likely to have revenues over $1 billion than their digitally novice counterparts. And the larger the enterprise is, the greater the gap in revenue. For example, 50% of large enterprises with strong digital maturity have annual revenues over $1 billion, compared with only 10 percent of small organizations with weak digital maturity.
The numbers show an undeniable trend and help shape an answer to the question “why do digitally mature companies have better results”.
A digitally mature company can be defined as a business that has invested in digital technologies to transform the way it does business to enhance customer relationships, improve employee engagement, and streamline processes.
Digital maturity is not just about having the latest gadgets or software packages. It’s a holistic approach to using technology to create value and transform your organization into one that can compete and win in today’s marketplace.
Many organizations are still struggling to make this shift. Their strategies for deploying digital technologies are often limited, and their applications are not fully functional or properly integrated. They don’t have the right tools or the right talent to execute on their plans. As a result, they’re increasingly vulnerable to disruption from start-ups, new entrants, and other competitors who are much more adept at using digital technologies to achieve competitive advantage.
Digital maturity is also essential for any organization seeking to achieve great things through innovation – whether bringing new products or services to market or delivering an enhanced experience for customers.
But digital maturity does not come easily – or quickly. If a company has reached maturity, it’s probably because the management team has been working on it for years. They’ve had plenty of time to get the details right. They’ve also built a solid customer base and cultivated positive relationships with partners along the value chain. In other words, the sooner you set off, the faster you get there.
In the last decade, organizations have begun to realize that they must implement a digital approach to their business if they want to stay competitive. In the last few years, we’ve seen many companies begin to make this shift.
What does it take for an organization to become digitally mature? There are four components:
1) Business strategy. Companies that put together a digital strategy from the top down with support from the board are more likely to move forward than those that don’t create one.
2) Technology strategy. A technology strategy is essential for development, but it must be in line with an organization’s overall business goals.
3) Governance. Establishing governance structures can help organizations focus on what’s most important during times of uncertainty.
4) Employee skills. Organizations need a skilled workforce to be successful digitally, but training and development aren’t enough—digital maturity programs must be incorporated into daily operations.
Truly digitally mature organizations—that is, those that are able to reap the benefits of digital transformation—are already seeing demonstrable results. They’re seeing a return on their investment of time, money and resources; they’re becoming more efficient and effective; and they’re capturing new revenue streams.

by Technical Team, EW Nutrition
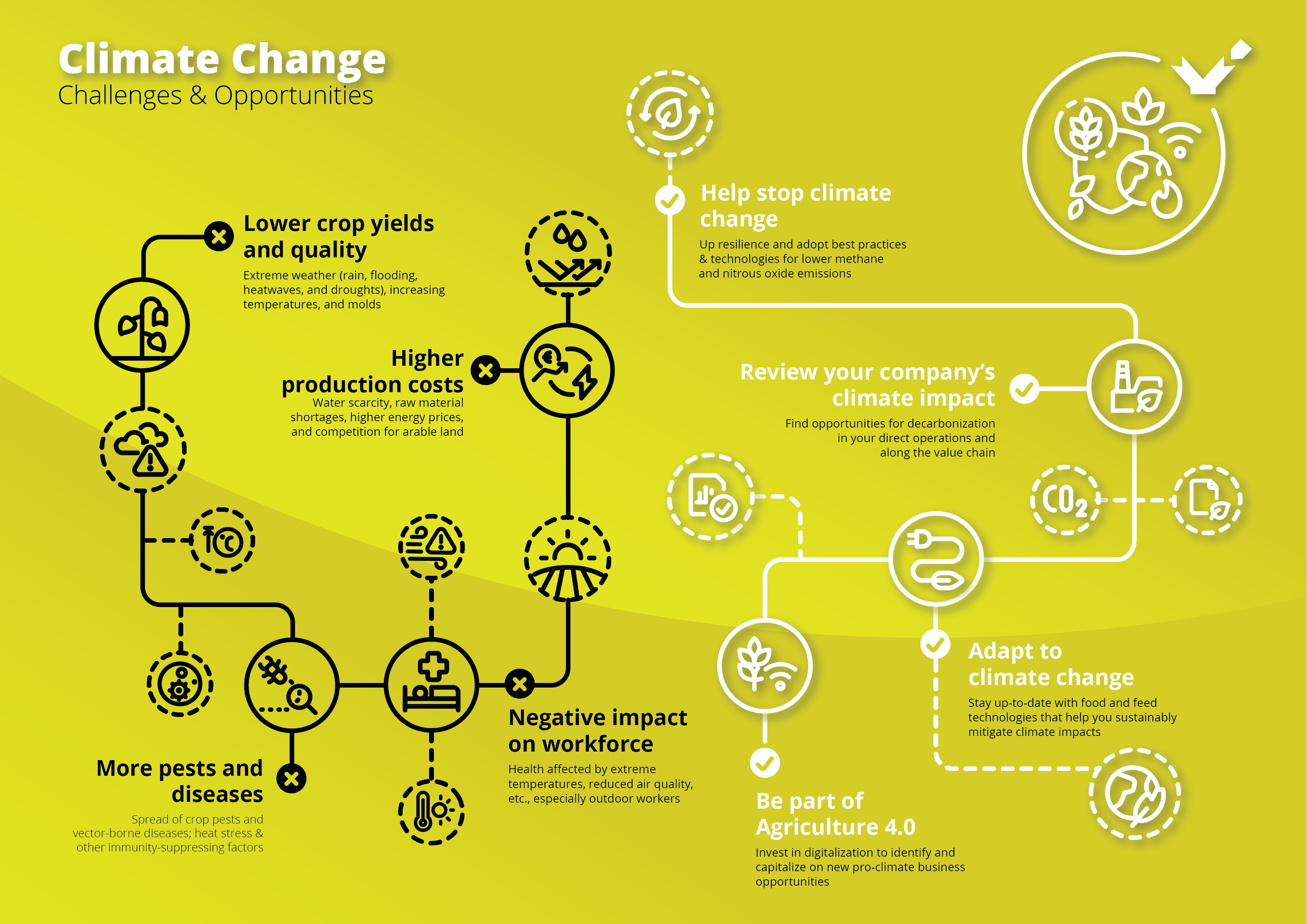
The Glasgow Climate Pact reached at the COP26 summit this November sent a clear message to businesses across the globe: Put sustainability on top of your corporate agenda or risk losing out. But how can food and feed producers translate the knowledge that climate change is happening into good business decisions? What impact is it causing, and which actions can we take today?
There is an overwhelming consensus among scientists that climate change is happening and that we need to stop it. Long-term changes in global temperature and weather patterns are nothing new. What is new is that these shifts are primarily driven by human activities.
Activities such as burning fossil fuels, livestock farming, and deforestation release carbon dioxides and other greenhouse gases (GHG) into the atmosphere. GHG trap the sun’s heat and cause average temperatures to rise.
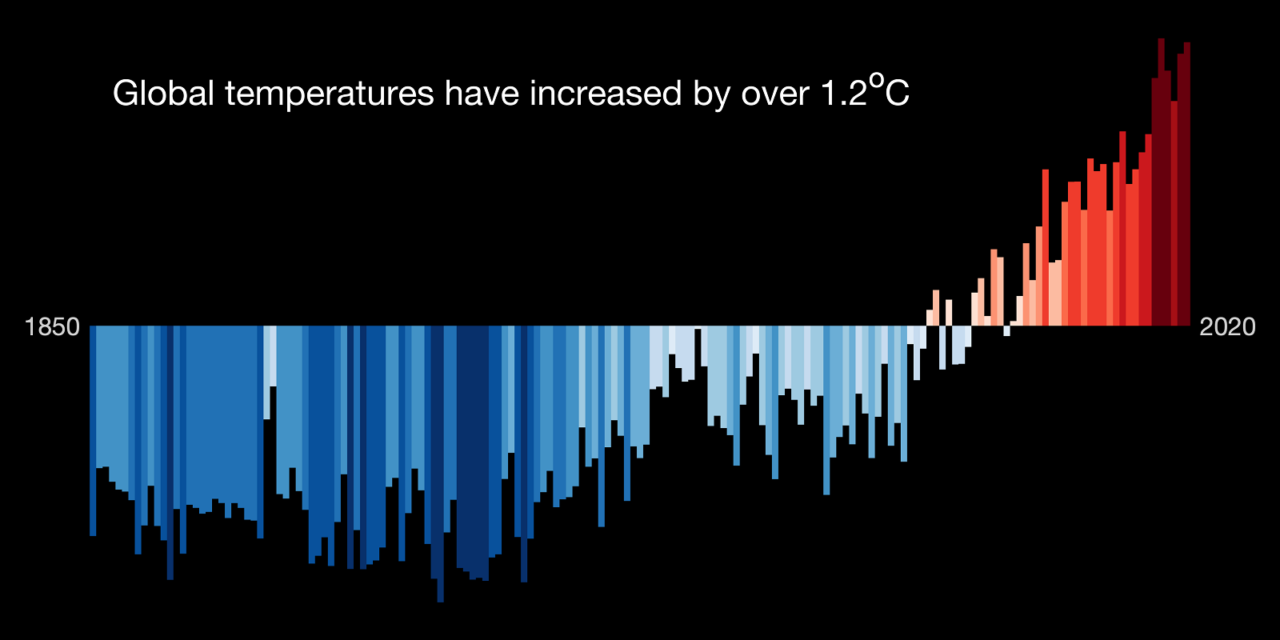
Compared to the pre-industrial era, average temperatures are now 1.2° C warmer. That doesn’t sound like a lot. But the effects are disastrous and disproportionately so for people in low-income communities and developing countries.
Climate change is already causing sea level rises, threatening coastal regions, and ocean acidification, which disrupts global seafood supplies. Unprecedented losses in biodiversity are compromising food security and ecosystem services (such as pollination and irrigation). Biodiversity losses also expose us to zoonotic diseases – for example, the ongoing COVID-19 pandemic has zoonotic origins.
Due to climate change, the frequency and severity of extreme weather events are increasing. As countries in Europe and China experience historic rainfall and flooding, other parts of the world, such as Australia, the western US, and many African countries, face intense droughts. With further warming, Pacific islands will disappear under rising sea levels. And regions such as the Middle East will suffer from extreme heatwaves and see farmland turn into deserts.
While some crops and areas benefit from higher temperatures and changed weather patterns, on the whole, it is becoming more difficult to feed the world. Extreme weather events such as droughts, floods, hurricanes, heatwaves, and wildfires pose severe challenges for agricultural businesses.
Agricultural production costs are and will be increasing further because climate change effects entail water scarcity, raw material shortages, higher energy prices, and stiffer competition for land as certain areas become climatically unsuitable for production.

Feed and food crop yields and quality suffer both from torrential rain and flooding and heat waves and droughts. Researchers from the University of Minnesota have found that climate change is already reducing global rice yields by 0.3% and wheat yields by 0.9% on average each year. Another study showed that every 1° C increase in global mean temperature would, on average, reduce global yields of wheat by 6.0%, rice by 3.2%, maize by 7.4%, and soybean by 3.1%.
We also see increased problems with pests and diseases. Pests already destroy 40 % of global crop production each year. As temperatures rise, pests from fall armyworms to desert locusts expand into new territories. Due to warmer temperatures, disease vectors such as mosquitoes, flies, and ticks also proliferate and migrate, carrying new pathogens to previously unexposed livestock. Additionally, decreased forage quality, heat stress, and water shortages already compromise livestock immunity.
And let’s not forget that the wellbeing and safety of the workforce are directly affected by climate change effects such as extreme temperatures and reduced air quality. This is even more true for outdoor workers and the 143 million “climate migrants” we expect to see by 2050. Climate change has also triggered a significant cultural shift, especially in younger generations. The climate-conscious GenZ talent only wants to work for employers who genuinely commit to sustainability.
Global warming must be limited to 1.5° C to avert the most devastating impacts. To achieve this, we have to cut greenhouse gas emissions in half by 2030. And by 2050, the world has to reach “net-zero” emissions, i.e., removing as many greenhouse gases from the atmosphere as we release into it.
Climate change regulations are becoming more stringent, especially for the energy, transport, and agricultural sectors. As UN High-level Climate Action Champion Nigel Topping puts it:
“If you haven’t got a net-zero target now, you’re looking like you don’t care about the next generation, and you’re not paying attention to regulations coming down the pipe. Your credit rating is at risk, and your ability to attract and keep talent is at risk.”
What can we do? Agriculture is unusual in that its contribution to anthropogenic climate change mostly comes from methane and nitrous oxide instead of carbon dioxide. Nitrous oxide emissions stem from soils, fertilizers, and manure, while ruminants and rice cultivation release methane.
Across these sectors, much more needs to be done at the policy level to incentivize sustainable husbandry, knowledge transfer, and targeted R&D. Still, wider adoption of existing best practices and technologies would help the global food and feed sectors to be more resilient and substantially reduce its GHG.
Enhanced efficiency fertilizers can reduce nitrous oxide emissions from soils, while phytogenic feed additives curb methane emissions in ruminants. And exciting research on topics such as pheromone-based pest control shows that climate change adaptation opens up new business opportunities.
Spend time on a detailed review of your company’s climate impact. This will reveal opportunities for emission reductions and decarbonization, both in your direct operations and along the value chain. More energy-efficient compound feed production, for example, helps feed mills but also improves the carbon footprint of the final animal products. And in times of Agriculture 4.0, investing in digital technologies, platforms, and processes will be vital to identifying and capitalizing on new business opportunities for climate-conscious production.
No one bears the brunt of climate change quite as intensely as agribusinesses. Let’s champion sustainability for improved corporate reputation, stronger risk management, long-run cost savings, market access, and attracting and retaining qualified employees and customers.

By Technical Team, EW Nutrition
COVID-19 is definitely still a concern across the globe. With only about 47% of the world fully vaccinated, chances are we will not see the end of it next year. And with the unequal distribution of vaccines, globalization will keep bringing it home.
However, while everyone has an eye on the disruptions caused by COVID-19 to businesses and society in general, there are other looming challenges to prepare for. Here is a brief look at the top 5 challenges that rose to the surface in 2021, which are forecast to cause even bigger waves in 2022.

Greenwashing doesn’t wash anymore.
In 2021, sustainability has become an unavoidable topic not just for ecologists and (once a year or so) heads of state, but also for businesses and the masses. With extreme weather phenomena looming large in the news, climate change has become a fact of life. It no longer needs to be accepted as much as it needs to be managed.
The challenge is twofold. One the one hand, businesses need to demonstrate corporate accountability by monitoring the environmental footprint of their activities. On the other hand, businesses need to preserve the financial viability of their sustainability initiatives.
The balance between accountability and financial viability is still hard to find. Sustainability in business is an expensive proposition. Not only is it costly to implement fully sustainable measures, but finding the sustainability hotspots in your value chain and retrieving the data is sometimes nearly impossible. And, while reporting for scope 1 and 2 is somewhat easier, scope 3 accounting is still a tough nut to break.
Based on the Greenhouse Gas (GHG) Protocol, emissions are divided by source into 3 major scopes:
Scope 1 – direct emissions of the reporting company. This includes owned or controlled sources such as company facilities or company vehicles.
Scope 2 – indirect emissions from consumption. Here are included purchased electricity, steam, heating and cooling for the company’s use.
Scope 3 – all indirect emissions along the value chain. Under scope 3 fall upstream and downstream emissions through purchased goods and services, capital goods, processing, transportation and distribution, waste, use of sold products, and much more.
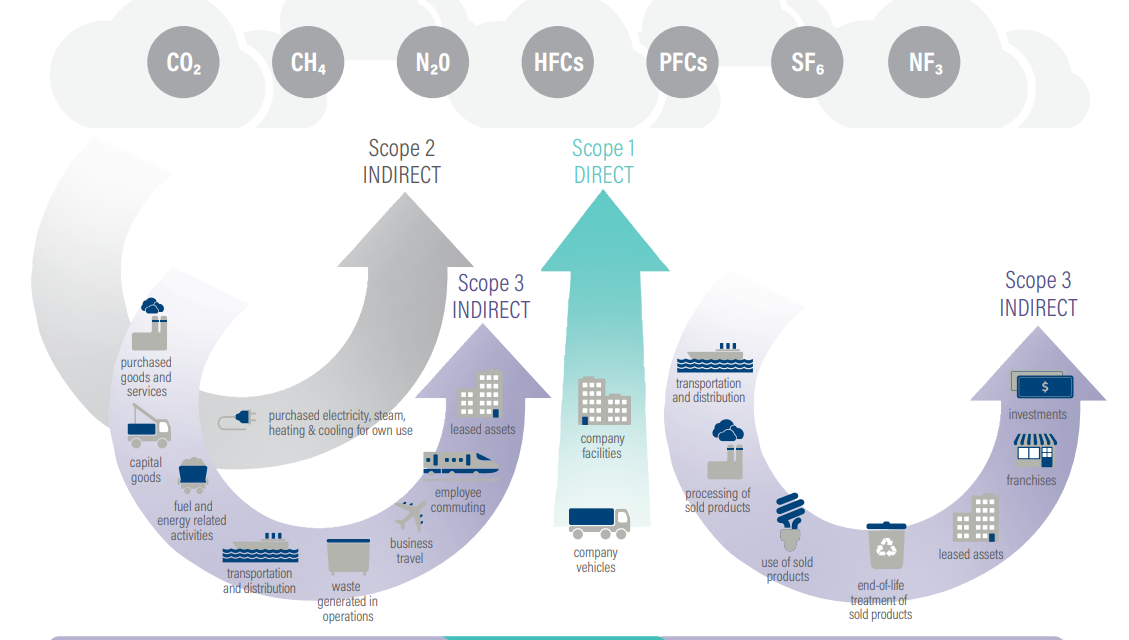
Source: GHG Protocol
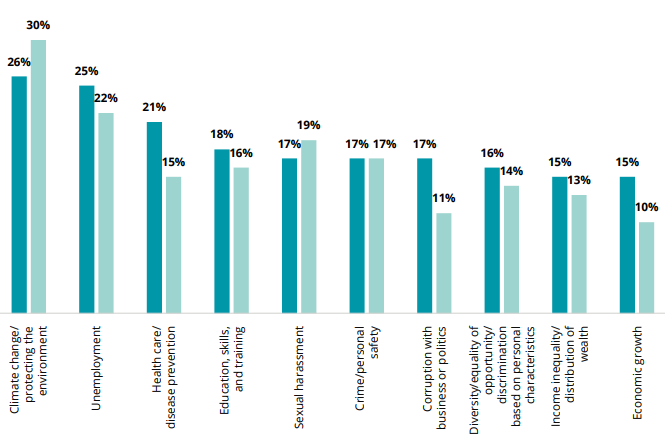
Nevertheless, the equation must balance. A study by Pew Research Center shows that sustainability and addressing climate change are not just thoughts, but immediate concerns for the younger generation. Gen Z (people born between 1997-2012) are actively engaged in following and/or combatting the effects of climate change. Research by Deloitte indicates that, for Gen Z, climate change and protecting the environment are the top concern.
When your upcoming workforce demonstrates such strong interest, it is remiss to ignore it.
This is getting serious.
Deadly snowstorm in Spain in January. A deadly heatwave that killed 569 people in Canada in June. Deadly floods that affected western Germany, The Netherlands, and Belgium in July. Deadly fires that raged across Italy and Greece in August. And many more deadly extreme weather events whose roots lie in climate change.
They are not just a reason to join the fight for sustainability. They are, in more immediate terms, a reason to examine how well your business is prepared to face the next extreme weather event. Because it is definitely coming. And it may affect your business – possibly directly, depending on your location, but definitely indirectly. The impact of resource scarcity could already be felt this year, when extreme weather events around the world affected harvest quantity and quality. Coffee and sugar futures have been on the rise for years, and so have wheat prices.
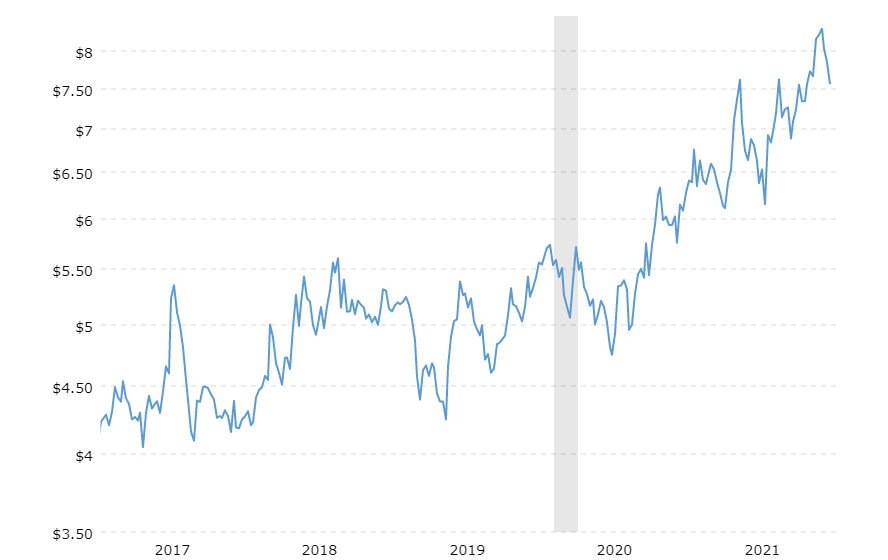
Source: Macrotrends
In fact, according to a 2021 Deloitte report, companies are actively concerned about the impact of climate change issues on their activities, with climate-related disasters topping the list and scarcity of resources a close second.
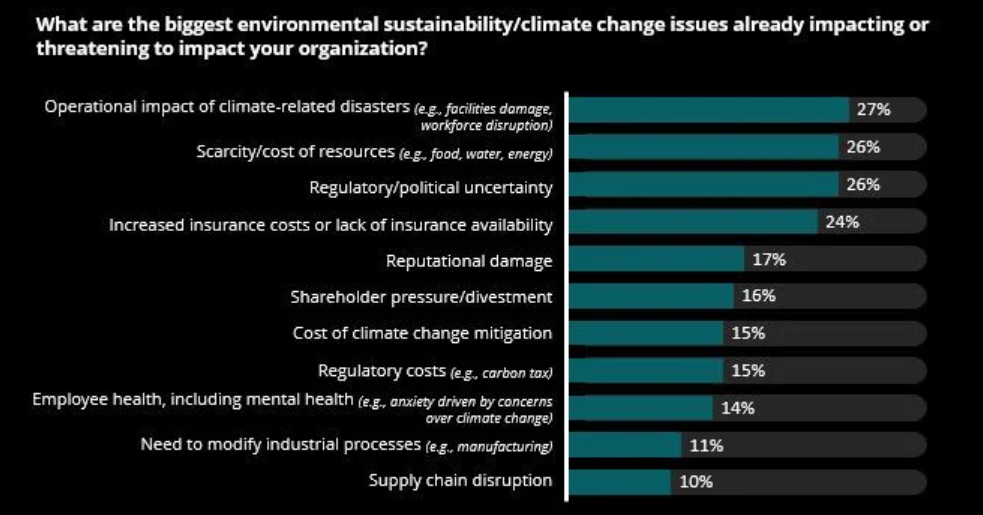
Source: Deloitte
Being prepared for extreme events means, in most cases, managing risks across company facilities, as well as across the value chain. The more widespread and varied the production footprint and sourcing capabilities, the better can a company weather the storm.
No, it’s not getting better soon.
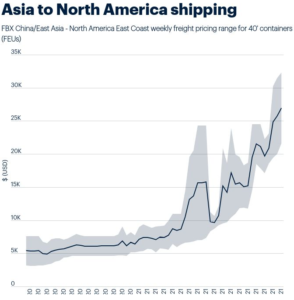
In fact, after over a year of constant disruptions, problems have accumulated into a perfect storm. Unpredictable demand and availability, as well as unpredictable labor shortages, both due to COVID-19 and various side-effects and accidents, have pushed container availability and port unloading infrastructure into a frenzied downward spiral. While container prices have gone up (over $20,000 for a standard container coming to the US from east Asia), ports have been unable to handle the incoming ships’ unloading needs.
Some ports have already moved to 24/7 operations, yet the issues will take much longer to fix – and prices are not likely to come down in a hurry. However, reliable forecasting in such turbulent times is nearly impossible.
In these circumstances, old wisdom can be reactivated. Instead of the cheaper and more agile just-in-time model that many companies practiced, increasing safety stocks and developing a wider global footprint are going to be the norm. On top, developing excellent relations with suppliers and managing customer expectations are critical. As Dan Swan, McKinsey Operations expert, put it: Supply chains are no longer a “necessary evil”, but more and more “a real differentiator for companies”. Product availability is the new name of the game.
In the meantime, expect prices and lead time to remain high for quite a while yet.
To the surprise of no one with any knowledge of markets, a 2019 study by Deloitte confirmed that businesses on their path to digital transformation tend to perform better financially. In the meantime, COVID-19 has accelerated this trend. With the Zoom boom and remote working, digital communications have become the norm – and time is not going back. In 2022, a mature company should also be a digitally mature company.
What does digital maturity mean? By and large, according to Deloitte’s concept of “pivoting to digital maturity”, it means that organizations use data and technology “to continuously evolve all aspects of its business models—what it offers, how it sells (interacts with its customers) and delivers, and how it operates”.
In 2022, organizations will feel even more under pressure to step up: connecting employees across multiple locations, integrating “business technologists” into their organigrams and empowering them to bring digitalization home, agility through autonomous modular processes, automation in every possible quarter, and more.
In some sectors, digitalization is self-implied. ICT, media, finance, and professional services are more digitally advanced than others. Logistics, retail, and other industries that rely on distributed databases will also make progress. However, even in agriculture and animal production, where most outsiders still picture tractors and barns, the fourth revolution is in full progress. The power of data, the possibilities brought by the cloud, the ease of synchronous communication will impact even the most conservative industries. Whoever is not prepared for digitalization will inevitably fall behind.
How is it possible to have both high employment and labor shortages at the same time?
#2 on the list of concerns for Gen Z, as evidenced in Deloitte’s research, is unemployment. And yet, the evidence is mounting that most companies (nearly 9 in 10, in one study) are having trouble filling certain positions – especially entry- and mid-level.
Source: Deloitte
In the US, analysts have been scratching their heads at a disconnect between 8.4 million unemployed and 10 million job openings, with radical differences in sectors such as professional and business services or education and health services.
Most answers focus on the realities of the “Corona times”. People who were forced to work remotely discovered that they enjoyed it. They left inflexible jobs for more flexible ones – or just to spend more time with the family.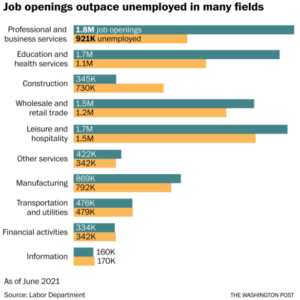
At the same time, childcare insecurity made it compulsory for some parents to give up their jobs to be home for children whose schools or care centers were closed in lockdown.
Workers in retail and hospitality, who saw their jobs being threatened, again and again, by COVID-19. Many were furloughed or sent into unemployment. While out of work, unemployment benefits allowed people to reconsider career paths – and many decided to move to more secure industries.
And, finally, workers whose jobs are not threatened fall prey to burnout during intense work-from-home bouts during long and/or repeated lockdown periods.
If companies have trouble filling positions, they might consider offering more flexibility. Another study by Gartner shows that, at least theoretically, employers and employees are aligned in seeing flexibility as critical to the organization. This includes as a top consideration the ability to work both from home and from the office, as well as somewhat flexible work times.
Yes, work-from-home will outlast COVID-19. At the same time, the concept of happynomics – the economics of personal happiness and well-being – is being transposed at work, with more corporate care for productivity through employee workplace satisfaction. It is by now quite clear that, the happier people are at work, the better the organizational outcomes.
***
Five challenges, each of them compounded by the prolonged uncertainty of COVID-19 and by looming political tensions in various hotspots around the world. Businesses large and small are already critically affected by all. No business will be spared from at least one of the 5 factors; many will be impacted by several or all.
In 2022, companies must be ready to pivot, change strategies, and adjust course mid-route. That doesn’t mean the first step shouldn’t be, always and most critically, planning for these 5 challenges. Dwight Eisenhower was right: Plans are worthless, but planning is everything.

By Ilinca Anghelescu, EW Nutrition
“Supply chain issues” is now a buzzword that has gone beyond the realm of business into our lives and households. We know holiday gifts might be delayed because of supply chain issues, mobile phones or electric cars are facing essential parts shortages never before imagined, and entire companies’ production and delivery are affected by supply chain issues. Why is this happening and what can be done?

The supply chain is an interdependent network of companies, individuals, databases, and actions whose ultimate goal is to ensure product or service delivery. The supply chain goes from the raw material to the end product or service that reaches the buyer. In most cases, this also includes the return of the goods or product servicing.
In the case of feed or food, for instance, a simplified version of the chain includes harvesting, storage, transportation, receiving and processing other raw materials, manufacturing, packaging, distribution, sale to the end customer, as well as potential returns at every stage along the way.
Supply chains are absolutely essential to the functioning of nearly everything in today’s society. If you eat tomatoes grown in your own garden, you are indeed outside the tomato supply chain; but your tools and fertilizer are probably not.
Supply chains are usually long, complex, highly interdependent, and often global because of cost pressure. No wonder, then, that one blow can bring down the whole domino chain. What is worse, the last year and a half brought not just one, but two concomitant blows.
The world is still reeling from the effects of COVID-19 – not just in terms of healthcare, but in terms of economic impact as well. In the immediate months after COVID-19 became a global concern, several things happened simultaneously to constrict the supply chain:
These and other similar measures put pressure on the global supply chain. Employee layoffs or furloughs, together with output decreases, meant fewer people could operate within the supply chain, which then translated into diminished production and delivery capacity.
To cope with the immediate state of things and with the predicted economic downturn, lots of companies (among those who did not shut down altogether) decided to reduce their output in order to save costs.
Despite the decreased demand for restaurant or hotel deliveries, lockdowns around the world brought about one other change: demand for endpoint deliveries soared. This would normally spell great news for producers, if only the supply chain weren’t in the way.
Supply chains are built to be as efficient as possible: fast, agile, and as economical as can be. In other words, “lean”. That also means that massive disruptions – whether positive or negative – are not easily handled. When demand for home deliveries went up dramatically, that would have been a good opportunity to beef up the delivery chain. Unfortunately, that increase in demand coincided with a limited personnel and product availability (because of blow no. 1), as well as shipping issues around the world.
How so? From the beginning of the pandemic until now, ships have been queuing up in ports around the world. The first Corona aftershock relocated production around the world and created unexpected demand. Containers were filled with high-margin goods (such as facial masks or home electronics) for the richer countries, while low-margin goods for other areas had to wait for free space.

This short-term unexpected move created competition for the limited number of containers on the market. Prices soared and containers traded at five times the cost of the previous year. (The Ever Given crisis in mid-2021 did not help, of course.) At the same time, unloading the cargo at the destination point had to be managed with reduced personnel, as pandemic restrictions still applied and docking availability remained the same. Ships had to wait for days to unload their cargo, despite high demand for their empty containers.
It turns out, not surprisingly, that we have all grown more addicted to technology during the pandemic. Increased demand in home electronics, coupled with issues in the production of silicone-based parts (mostly led by a drive to reduce power consumption in China), led to a worldwide chip shortage that is affecting companies across many industries. Most notably, Tesla, Apple or Qualcomm are struggling to deliver orders and are readjusting their launch and delivery plans. If your mobile phone is delivered weeks late, blame it on the supply chain.
In 2022, keep your expectations low. The issues will not be resolved in a couple of months, so you must prepare for a year of very slow progress.
Prepare for at least two more years of high costs. Delivery delays will hit some markets more dramatically. Chip manufacturers have already announced shortage expectations throughout 2023, with lead times growing from 9 to 20+ weeks. Many large players in tech have announced plans to build their own plants or replace chips (as Tesla did) with different technologies. Do not be overly optimistic, though: such plans take a long time – and ports are reeling from a new wave of COVID-19 infections and restrictions. At this point, it is not just the shipping costs that are running a little wild, with no incentive for the cargo players to lower them; it is also the ports themselves. In the port of Los Angeles, as in many around the world, it is not unusual for cargo to wait a month out at sea to be given access to docking and unloading.
Order early. This seems like a commonsensical step – and it is, at this point. As mentioned above – do not expect things to be resolved quickly and painlessly. Protein markets are hit just as much as tech components; ship and truck deliveries will take a very long time to recover across all sectors. Demand is increasing and supply is still low – and slow. Order early and strategize to cover the new cost structures.
Explore options around the world. With China and Western Europe getting hit by new variants and suboptimal vaccination levels, the rest of the world remains to be explored. Options in richer countries, that once seemed too expensive, now may have an edge. Look closer to home rather to what was formerly a more cost-effective source. With shipping costs 3x to 10x what they were two years ago, the incentive is gone. (Even so, air freight is prohibitively expensive, so land access may be the most realistic option where accessible.)
Expand your capacity. Not everyone can start building plants at the drop of a hat, like Apple; but many companies can consider adding production lines or partnering with allies (and even competitors) to build or exploit resources. Since the supply chain issues are expected to last a couple more years on the optimistic side, it’s a good idea to explore your options not just in geography, but in partnerships for capacity as well.
Prepare for worse times before the good ones. It’s always a good idea to be prepared, but even more so now. Some ports are already closing again in early December (for instance, Dalian, China, took early measures against an outbreak) and the world is reacting to another potential large-scale lockdown.
On top of that, recession seems inevitable in many industries, and the new Corona mutations have already pushed the stock market down a few notches. Inflation has increased – 6.2% in the US in October versus last year, and in the eurozone the 4.1% inflation is the highest in the past 13 years. Some industries fare well simply because the market need is there (food and feed among them), but remember that the supply chain has companies across industries fighting for the same cargo space.
The answer to the supply chain challenge is to practice caution, spread risk, and actively seek out opportunities – including partnerships, new business, and unexpected collaborations. But then, these are good business practices at all times.

By Ilinca Anghelescu, EW Nutrition
Momentous changes are happening in our lives and our industry. A lot of them are caused by globalization – COVID-19 or the effects of supply chain disruptions, among the more recent ones. Many more, though, that impact our personal and professional lives, are caused by digital advances omnipresent in contemporary society. And, although some view agribusinesses as a conservative industry, that has long not been the case. Mature companies have their eyes on the fourth agricultural revolution – and digitalization is a large part of it.
Agriculture has long been a motor of progress in human lives. Each agricultural revolution has brought about enormous improvements in living standards and consequently in life expectancy.
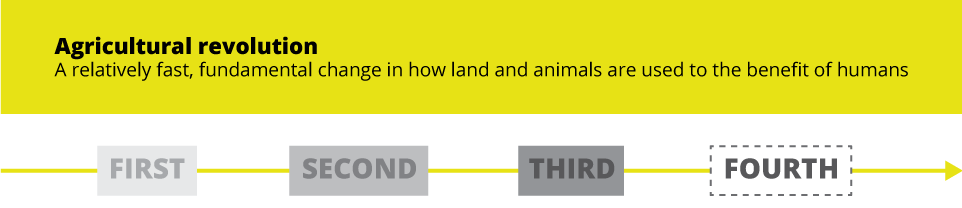
The first agricultural revolution took place about 10,000 ago, when hunter/gatherer communities began to settle and grow crops for sustenance. The revolution largely consisted of the domestication of plants and animals, as well as with agricultural processes. This revolution, it is widely believed, altered the course of human history and even biology: humans were able to form settlements and have predictable and nourishing food sources. Through that we also developed caries, body fat, and deficiencies in fiber and micronutrients.
The second agricultural revolution happened much later, from the mid-17th to the late 19th century, starting in the British Isles. The revolution consisted largely of enormous production improvements, not just from increased human labor but also from innovations such as advanced ploughing techniques, crop rotation, plus selective animal breeding, improved transportation, and land drainage. These developments helped the empire sustain a demographic, as well and geographic expansion.
The third agricultural revolution, in the 1950s and ‘60s, was prompted by enormous strides made in chemical fertilizers, irrigation, mechanization, and the development of new, high-yielding crops. These advances made possible a yield increase of over 40% in the course of less than 50 years. The changes were even more dramatic in developing nations, where the standard of life saw huge improvements with the advent of high-yield rice, wheat, and corn.
Naturally, poverty is still a major concern. However, if poverty has declined by 85% between the 1860s and 2020, we largely have agriculture to thank for.
The fourth agricultural revolution has been touted for years now. Part of the wider 4IR (Fourth Industrial Revolution), it is widely seen as the next dramatic improvement in the quality and quantity of agricultural and livestock output. The fourth agricultural revolution (alternatively, Agriculture 4.0) has been prompted, just like 4IR, first and foremost by digital advances. It relies, among many other innovations, on automation, gene editing, nutrigenics, traceability, and precision livestock farming.
In many cases, these advances are not yet fully developed, not fully embraced, or not yet fully integrated into one standardized system. However, the trend is unmistakable and unstoppable: the fourth agricultural revolution has started.
EW Nutrition is starting a series of articles on the challenges of digitalization and digital transformation in the livestock production industry and in industry in general. These advances in the way crops and livestock are being developed, grown, processed, and delivered from the farm to the end consumer’s home are indeed radical.
More important, however, is to prepare the ground for what’s coming. If organizations are not digitally savvy and technologically advanced, how will they deal with the abundance of data that Agriculture 4.0 relies on?
There is a lot of confusion about the three terms, especially because digitization and digitalization are often used interchangeably. However, in terms of business strategy there are clear differences which could be clarified if we visualize the three terms as a funnel.
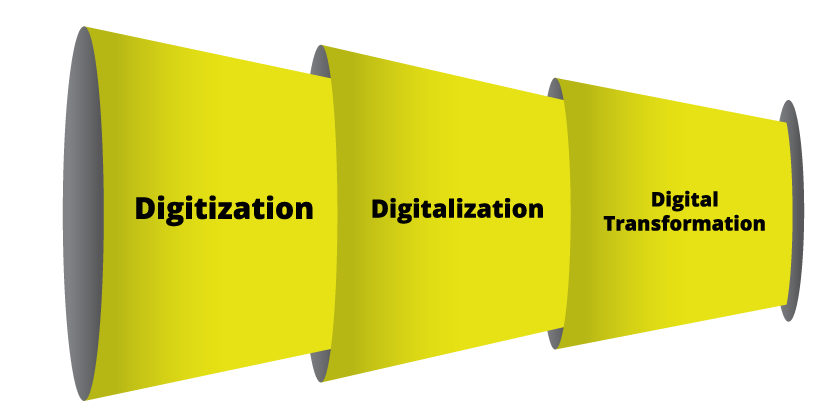
Digitization is the top level which most companies can easily reach. It refers to transferring a physical object to a digital representation. Think of it in terms of converting a 19th century novel to an e-book or turning a company ledger into an Excel spreadsheet.
Digitalization is the middle level of the funnel; fewer companies easily accede here, although it is nowadays an essential part of most businesses. Digitalization means using digital technologies to improve business processes and work more efficiently. Examples would be cloud storage for company files, virtual platforms for team conversations and project tracking, etc.
Digital transformation is transforming a business by using digital technologies, platforms, and processes to enable change, optimize the business model, and deliver better results for the organization. Examples would be an e-commerce company that leverages information from machine learning and combines it with big data across its vertical to collect prospect information, pool data into a SSoT (single source of truth), and conduct analytics that inform predictive algorithms.
Sure, not all companies can – or should – move completely into the bottom layer of the funnel. Digital transformation can be partial and should only be leveraged in companies and industries where it makes sense.
Where exactly is your company along the funnel? Assessing your current status is essential to developing a strategy to meet Agriculture 4.0 head on. And meeting it head on we must; it is no longer an option, but a necessity for organizations and businesses to remain relevant in tomorrow’s world.

Regardless of the vertical in which you are operating, in 2022 your organization should be able to check most of the boxes suggested by Deloitte in the table above.
At the most superficial level, digital maturity is a good predictor of improved financial performance. The more digitally savvy companies are cashing in on their maturity.
Behind this predictive factor, however, is the reason why. Digitally mature companies
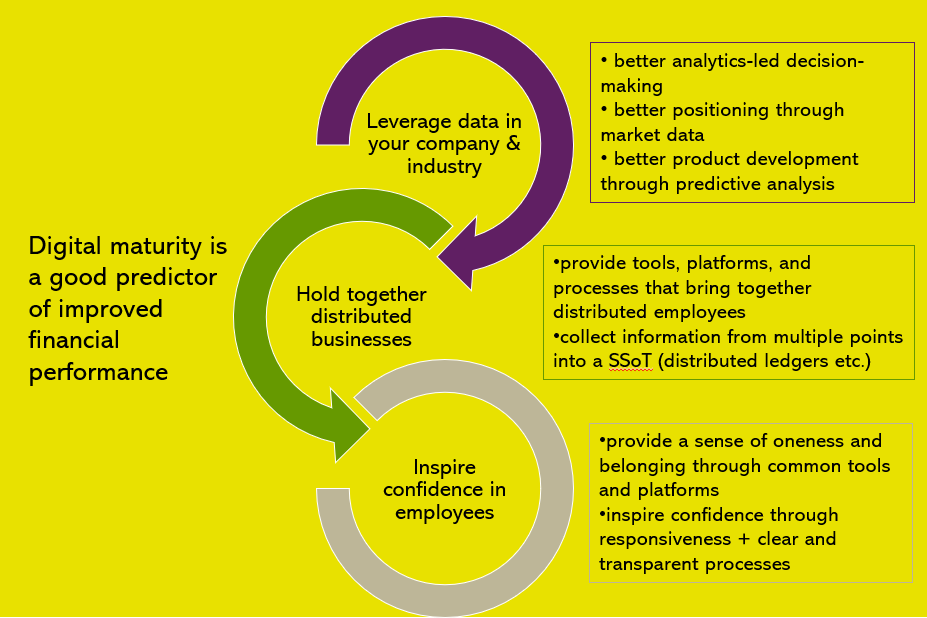
The road to digitalization begins, first and foremost, with the will to change. Digitalization is change – and change is not easy, especially in more conservative industries or companies. Once the management understands the benefits of undertaking this process (which benefits are confirmed in multiple studies), there are several roads to choose from. However, to enable the process of digitalization in general, McKinsey identifies five key factors:
Once the organization has undergone the basics of change, the company can then strategize on how to take advantage of the trends in its specific market. In the animal production industry, these trends – briefly mentioned at the beginning – amount to a revolution.
The revolution is already underway. For the moment, however, it is developing on so many disparate fronts that there is no formal coherence and very little oversight. Because of that, but most especially because of digital immaturity, very few companies or institutions are prepared to deal with what is coming. Now is the time to get in shape and get the process started.
As we grew over the past few years, both organically and through M&As, we faced a few challenges that many will be familiar with:
We were fortunately prepared to deal with most of the challenges. Even so, we were not 100% prepared. We have learned enormously in the course of these years and are now a few steps ahead.
Over the next couple of months, EW Nutrition is going to look at some of the most important topics around digitalization in general, digitalization in livestock and feed production, and obstacles to building a digitally mature company:
The process is never complete, of course. We just hope that, by learning in public and sharing our discoveries, we make our journey clearer – and perhaps other companies’ journey easier.
FAO. The role of digital technologies in livestock traceability and trade, 2020
Gartner. Manufacturing Industry Scenarios in 2023: Leading Through Innovation, published September 2018, updated February 2020.
Gartner, Top Strategic Technology Trends for 2021, 2020
Gurumurthy, Ragu and David Schatsky, Pivoting to digital maturity: Seven capabilities central to digital transformation, Deloitte Insights, March 13, 2019
Gurumurthy, Ragu et al., Uncovering the connection between digital maturity and financial performance, Deloitte Insights, May 2020, 2020
Kane, Gerald C., “Accelerating digital innovation inside and out: Agile teams, ecosystems, and ethics,” MIT Sloan Management Review and Deloitte Digital, June 4, 2019
McKinsey. Unlocking success in digital transformations, October 2018
Moatsos, Michail. Global extreme poverty: Present and past since 1820. In How Was Life? Volume II : New Perspectives on Well-being and Global Inequality since 1820, OECD 2021
Sharma, Deepak et al., Customer-centric digital transformation: Making customer success integral to the new organization, Deloitte Insights, September 5, 2019
Schwab, Klaus, The Fourth Industrial Revolution: what it means, how to respond, World Economic Forum, 14 Jan 2016
World Economic Forum, Innovation with a Purpose: The role of technology innovation in accelerating food systems transformation, January 2018
World Economic Forum, Technology Futures: Projecting the Possible, Navigating What’s Next. Insight Report, April 2021

By Dr. Inge Heinzl, Editor, EW Nutrition
Nowadays, the whole world is talking about sustainability. Many efforts aim to maintain our world for future generations, creating a balance between our current needs and those of our children, grandchildren, and great-grandchildren. The right animal nutrition choices play a crucial role in achieving the challenging aim of sustainable animal production.

The idea of sustainability is not new. Already the first humans lived sustainably, taking only as much as they needed and the environment could cope with, using all parts of the animals they killed. The German Hannss Carl von Carlowitz (1645-1714) coined the term sustainability in his oeuvre “Sylvicultura oeconomica” to counter a threatening raw material crisis. Wood was one of the most important raw materials. Besides heating, it was used for shipbuilding and mining. This was the reason that extensive areas in Europe were deforested and became deserted. Observing the impending disaster, von Carlowitz ” (1713) stated that only as many trees should be felled as can grow back through planned reforestation, sowing, and planting.
The Brundtland Report (1987), a document created by the World Commission on Environment and Development, is reckoned to be the starting signal for worldwide discussions about sustainability. In 2015, the result of a meeting of 193 members of the United Nations was the Agenda 2030 with 17 sustainable development goals for a “world we want” that should be achieved by 2030.
Sustainable Development Goals (SDG) of the Agenda 2030, fixed by the UN in 2015
The animal nutrition industry’s sustainability efforts play into different SDGs, notably no. 2, zero hunger, no. 3, good health and well-being, no. 12, responsible consumption and production, no. 13, climate action, no. 14, life below water, and no. 15, life on land. In addition to the overarching goal of fostering higher animal welfare (cf. Keeling et al., 2019), the feed sector’s measures center on three areas:
One of the 17 points on the list of the United Nations is “responsible consumption and production”. For the feed industry, this means making the most out of available feed sources. Improvements in feed conversion, the maintenance of feed quality, and the use of alternative ingredients are all part of this.
The feed conversion rate shows the amount of feed consumed in relation to the outputs produced, such as weight gain, eggs, or milk. The better or lower the feed conversion rate (FCR), the less feed you need to achieve your target, and the higher the yield. Products that improve feed conversion, therefore, can help to save resources.
Good feed conversion or an optimal utilization of nutrients depends on gut health. Only a healthy gut can digest the feed and absorb the nutrients adequately. Hence, products to improve feed conversion often do so by improving gut health.
Herbs and their active components have been used in human and veterinary medicine for thousands of years to treat digestive tract diseases. Nowadays, products based on phytomolecules help improve feed conversion through their digestive, anti-inflammatory, and antimicrobial effects on the intestinal tract.
How do these three characteristics contribute to a better FCR?
FCR improvements in broilers thanks to ACTIVO found in several studies
As phytomolecules are often volatile, EW Nutrition offers encapsulated phytomolecule-based products for the feed (ACTIVO product line). During episodes of elevated enteric challenge, e.g., weaning or following feed change, a liquid solution (ACTIVO LIQUID) can be applied via the waterline.
Some feed materials are hard to digest for certain animals. For example, pigs’ digestive systems do not have the enzymes required to break down non-starch polysaccharides (NSPs), such as cellulose, hemicellulose (ß-glucans and xylans), pectins or oligosaccharides. However, pig feed ingredients usually contain these substances.
Besides the non-usability of NSPs, the cage effect is a further problem. Cellulose and hemicellulose, water-insoluble NSPs, encage nutrients such as proteins or digestible carbohydrates. Encaged nutrients cannot be reached by the digestive enzymes and don’t become available to the animal.
Xylanases are available on the market to degrade structural substances in the feed and make them, as well as the nutrients they encaged, available for the organism.
Another possibility to save resources is the maintenance of feed quality. Bad weather conditions at harvest or incorrect storage can downgrade feed quality due to the development of molds and their mycotoxins or the oxidation of nutrients. Products mitigating the adverse effects of toxins, acidifiers that reduce microbial load, and antioxidants can help to keep your feed quality on a high level – or to re-establish it.
Feed materials contaminated with mycotoxins harm animals in different manners and should not be used without further treatment. Mycotoxins are not visible – even if no molds are visible, mycotoxins might be present. Additionally, they are pH- and thermo-stable, meaning that mycotoxins produced in the raw materials on the field remain in the finished feed. As mycotoxins often do not cause apparent, specific symptoms but manifest in decreased performance, feed refusal or lower feed intake, and higher disease susceptibility, it is difficult to notice contamination.
Products such as SOLIS or MASTERSORB contain clay minerals (bentonite and montmorillonite) that adsorb the toxins. MASTERSORB GOLD and MASTERSORB FM also include toxin-adsorbing yeast cell walls and herbal substances to help protect the liver.
 Feed spoilage through molds, yeasts, and mycotoxins wastes precious resources
Feed spoilage through molds, yeasts, and mycotoxins wastes precious resources
Acidifiers based on organic acids counter harmful microbes in the feed in two ways. Most pathogenic bacteria are susceptible to low pH. The proliferation of, e.g., E. coli, Salmonella, and Clostridium perfringens is minimized at pH < 5 (cf. Fuller 1977). Acidic-tolerant beneficial bacteria such as Lactobacilli or Bifidobacterium, however, survive.
Other than antimicrobial activity, organic acids also cause a significant reduction in ammonia (Eriksen et al., 2014). This finding could be due to a reduction in the microbial deamination of amino acids, which would then be available for absorption, resulting in increased nitrogen digestibility and reduced ammonia excretion, as observed in monogastrics fed organic acids (Pearlin et al., 2020).
The acidifier product lines ACIDOMIX, FORMYCINE, and PRO-STABIL all help protect feed from contamination with pathogenic microorganisms.
The oxidation of nutrients in the feed decreases its nutritional value and, thereby, the value of the whole diet. Fat, proteins, fat-soluble vitamins, pigments, and other biologically active molecules, including sugars and phospholipids, can get oxidized. Metal ions and other pro-oxidative factors can affect the ingredients of the feed during mixing, storage, and feeding. The oxidation of fats and fat-soluble vitamins results in color changes or odors and – this is even more serious – in the production of harmful substances such as aldehydes and ketones. An oxidized feed can lead to oxidative stress in the animals, reduce their immunity, productivity, and livability.
To protect valuable ingredients, the timely addition of effective antioxidants such as STABILON is recommended.
Soybeans are an excellent source of protein in animal nutrition. During the last 50 years, soy production has increased from 27 million tons to 269 million tons, causing environmental degradation of forests and savannas (WWF, 2021). The use of alternative protein sources helps protect our environment.
Ruminants partly cover their protein requirements with the help of rumen bacteria. These bacteria can turn nitrogen from urea into bacterial protein, provided they receive enough energy available from carbohydrates. Thanks to its encapsulation, PROTE-N, a feed-grade urea-based nitrogen source, slowly releases nitrogen into the rumen, synchronized with the energy supply. PROTE-N affords producers a degree of independence from soybean protein without compromising nutritional quality.
 Reducing soybeans in ruminant feeds helps to lower their environmental impact
Reducing soybeans in ruminant feeds helps to lower their environmental impact
Animal production generates gases such as ammonia and methane that negatively impact the environment. Measures to reduce these gases help to protect plants, animals, us, and our globe.
Besides nitrogen oxides, ammonia is one of the primary sources of nitrogen pollution. Ammonia damages ecological systems through acidification and nutritional oversupply. Fast-growing plants that need high amounts of nitrogen or plants that tolerate low soil pH proliferate, whereas more susceptible plants disappear, decreasing biodiversity. According to Max-Planck-Gesellschaft (2017), reducing ammonia emissions by 50 % could prevent 250.000 deaths caused by fine dust worldwide per year.
Improved protein digestion in animals reduces their ammonia production. Decreasing the intestinal pH through using organic acid-based products such as ACIDOMIX or FORMYCINE is essential for the activation and correct functioning of the enzymes responsible for protein digestion.
Together with CO2, N2O, and three fluorinated gases, methane belongs to the greenhouse gases listed in the Kyoto protocol. Being over 25 times more potent than carbon dioxide at trapping heat in the atmosphere, it dramatically affects the earth’s temperature and the climate system (United States Environmental Protection Agency). Methane is a final product of feed fermentation in the rumen and is produced by methanogenic bacteria. Ruminants can produce 250-500 L methane per day (Johnson & Johnson, 1995).
Reducing methane production in ruminants is a critical step towards climate protection. Herbal substances can change the microbiome, leading to improved protein and fiber degradation and reduced methane production (Ku-Vera et al., 2020). ACTIVO PREMIUM is a phytomolecules-based product for ruminants that helps reduce their methane emissions.
To preserve the environment, reducing energy needs is also an important topic. Using the surfactant SURF-ACE in the pelletizing process, feed mills can cut 10-15 % of their energy consumption or produce up to 10-15 % higher pellet output without increasing their energy consumption. When moisture is added together with the surfactant, the emulsion of the dietary fat and the added water leads to better general lubrication of the machinery and improved press throughput.
 Feed mill efficiency is key to animal nutrition’s carbon footprint
Feed mill efficiency is key to animal nutrition’s carbon footprint
Point 3 on the UN’s Agenda 2030 is good health and well-being. For many years, antibiotics, a very effective weapon, have been used to fight bacterial diseases. However, the occurrence of resistance is increasing. One of the reasons is the inappropriate use of antibiotics. These substances are often used preventively or for viral diseases against which they are ineffective. Also, the use of antibiotics as growth promoters at low dosages in animal production strongly contributed to the development of antimicrobial resistance.
Limiting antibiotic use to therapeutic treatment is possible through good farm management and feed supplements that support animals’ gut health, immune systems, and respiratory health. For this purpose, solutions ranging from phytomolecules (ACTIVO products, GRIPPOZON) to egg immunoglobulins (GLOBIGEN products, PROTEGG), products mitigating the impact of toxins (MASTERSORB products, SOLIS), beta-glucans/MOS (BGMOS), and acidifiers (ACIDOMIX, FORMYCINE) are available.
The animal nutrition industry provides many products to support animal producers in coping with their main challenges, including the shift to more sustainable production practices. Solutions exist to save feed resources, better protect the environment, and keep antibiotic tools effective. As an additional reward, implementing sustainability solutions leads to healthy animals with high performance. Let’s all help to preserve this planet for our next generations!
Eriksen, J., Nørgaard, J. V., Poulsen, H. D., Poulsen, H. V., Jensen, B. B., & Petersen, S. O. (2014). Effects of Acidifying Pig diets on emissions of AMMONIA, methane, and sulfur FROM Slurry during storage. Journal of Environmental Quality, 43(6), 2086–2095. https://doi.org/10.2134/jeq2014.03.0108
Fuller, R. (1977). The importance of lactobacilli in maintaining normal microbial balance in the crop. British Poultry Science, 18(1), 85–94. https://doi.org/10.1080/00071667708416332
Johnson, K. A., & Johnson, D. E. (1995). Methane emissions from cattle. Journal of Animal Science, 73(8), 2483–2492. https://doi.org/10.2527/1995.7382483x
Keeling, Linda, Håkan Tunón, Gabriela Olmos Antillón, Charlotte Berg, Mike Jones, Leopoldo Stuardo, Janice Swanson, Anna Wallenbeck, Christoph Winckler, and Harry Blokhuis. “Animal Welfare and the United Nations Sustainable Development Goals.” Frontiers in Veterinary Science 6 (October 10, 2019). https://doi.org/10.3389/fvets.2019.00336.
Ku-Vera, J. C., Jiménez-Ocampo, R., Valencia-Salazar, S. S., Montoya-Flores, M. D., Molina-Botero, I. C., Arango, J., Gómez-Bravo, C. A., Aguilar-Pérez, C. F., & Solorio-Sánchez, F. J. (2020). Role of secondary plant metabolites on enteric methane mitigation in ruminants. Frontiers in Veterinary Science, 7. https://doi.org/10.3389/fvets.2020.00584
Max-Planck-Gesellschaft. (2017, October 27). Reducing manure and fertilizers decreases atmospheric fine particles. Max-Planck-Gesellschaft. https://www.mpg.de/11667398/agricultural-emissions-fine-particulate-matter.
Pearlin, B. V., Muthuvel, S., Govidasamy, P., Villavan, M., Alagawany, M., Ragab Farag, M., Dhama, K., & Gopi, M. (2020). Role of acidifiers in livestock nutrition and health: A review. Journal of Animal Physiology and Animal Nutrition, 104(2), 558–569. https://doi.org/10.1111/jpn.13282
United Nations. (n.d.). How your company can advance each of THE SDGS: UN Global Compact. How Your Company Can Advance Each of the SDGs | UN Global Compact. https://www.unglobalcompact.org/sdgs/17-global-goals.
United States Environmental Protection Agency. (n.d.). Importance of methane. EPA. https://www.epa.gov/gmi/importance-methane.
von Carlowitz, H. C. (1713). Sylvicvltvra oeconomica, oder, Hausswirthliche Nachricht und Naturmässige Anweisung zur Wilden BAŬM-ZŬCHT: Nebst gründlicher darstellung, wie Zu FÖRDERST durch Göttliches Benedeyen Dem allenthalben und insgemein einreissenden Grossen Holtz-mangel: Vermittelst Säe-pflantz- und Versetzung Vielerhand Bäume zu prospiciren …: Worbey zugleich eine Gründliche nachricht von den in Churfl. Sächss. Landen gefundenen Turff Dessen Naturliche beschaffenheit, Grossen NÜTZEN, Gebrauch und nutzlichen verkohlung, Aus Liebe Zu BEFÖRDERUNG des Algemeinen Bestens beschrieben. Verlegts Johann Friedrich Braun.
World Wildlife Fund. (2021). Soja – die Nachfrage steigt. WWF Startseite. https://www.wwf.de/themen-projekte/landwirtschaft/produkte-aus-der-landwirtschaft/soja/.

By Technical Team, EW Nutrition
Antibiotics are a precious resource whose long-term efficacy must be protected – for human and animal health. This is a difficult challenge for healthcare providers (veterinarians and medical doctors), as well as farmers and integrators. In this series of articles, we will explore the general and specific measures and solutions through which animal production can contribute to the overall reduction of antibiotic use.
 Responsible animal production contributes to maintaining antibiotic efficacy
Responsible animal production contributes to maintaining antibiotic efficacy
Shortly after the discovery of penicillin in 1929, Alexander Fleming already pointed out the possibility of resistance during an interview with the New York Times. The first case of penicillin resistance was reported only one year after clinical trials began; within 20 years, 80% of Staphylococcus aureus isolates were resistant to penicillin (Lobanovska and Pilla, 2017).
Over the years, clients and patients have gotten used to receiving a pill to quickly fix their ailments. Often, antibiotics have been prescribed for illnesses they were not effective against, including viral challenges. This has unnecessarily accelerated the rate of resistance development. To reverse this trend, education is key. At the same time, the judicious use of antibiotics, meaning the correct antibiotic for the challenge plus proper administration and duration of use, is paramount for all medical professionals to help preserve the efficacy of these critical substances.
For many years, animal producers have used antibiotics as growth promoters. The E.U. banned this type of use in 2006, and the United States followed in 2017. Evaluations have shown a decrease in antibiotic use in the U.S.: In 2014, according to the FDA, 17,000 tons of antibiotics were sold in the United States for livestock, representing 80 percent of all U.S. antibiotics sales. In 2019, a total of about 11,000 tons of antibiotics were sold for use in food-producing animals (FDA, 2020).
As the number of isolated multi-drug resistant bacteria increases and the discovery and approval of new antibiotics slows, it is imperative that the use of antibiotics in animal production, especially those that are critically important for humans, is reduced to a minimum. Hence, antibiotics should only be used to treat, control, or prevent diseases in case of imminent risk, but not for growth-promoting purposes.
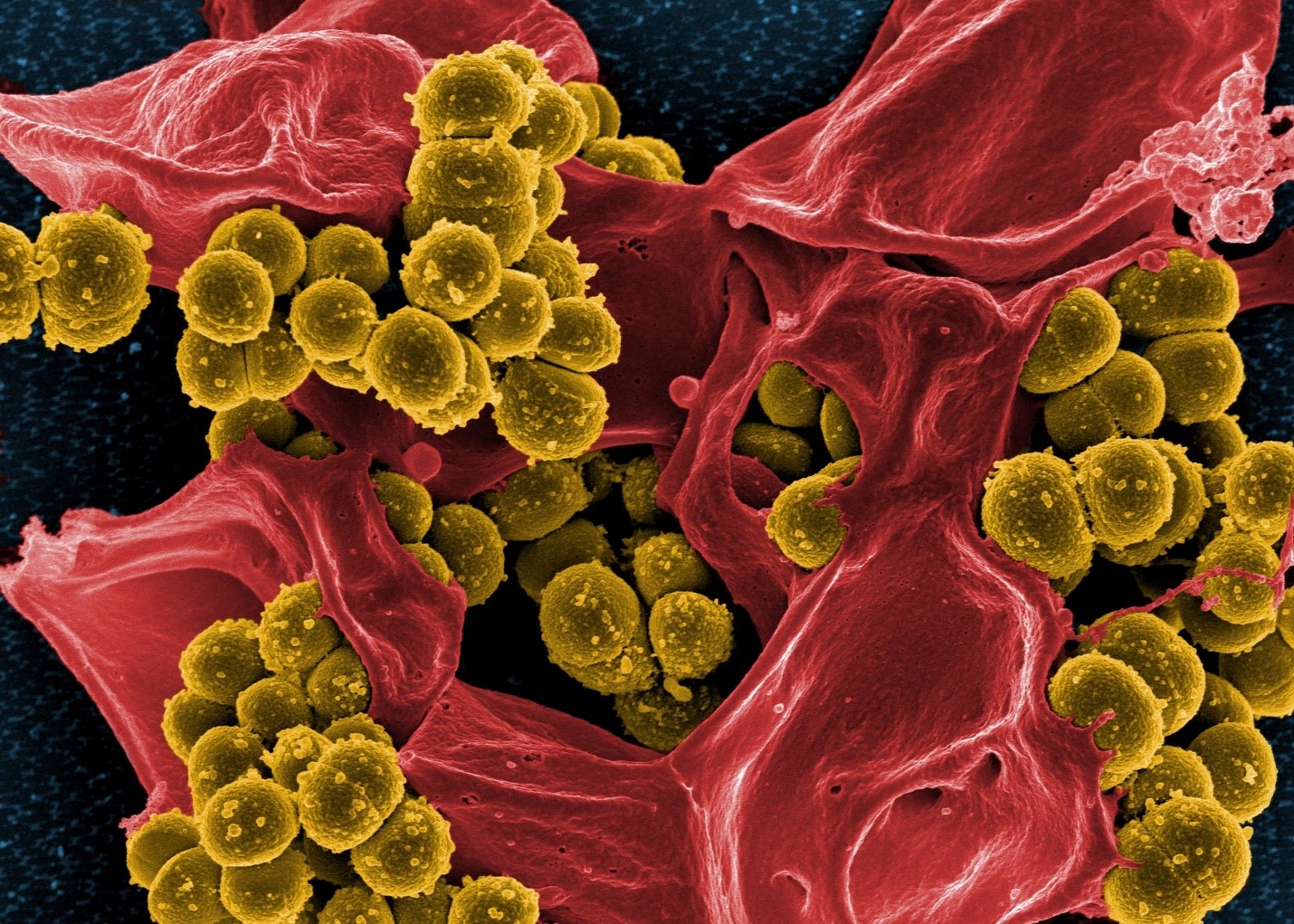 Scanning electron micrograph of methicillin-resistant Staphylococcus aureus bacteria (yellow) and a dead human white blood cell (red). Credit: National Institute of Allergy and Infectious Diseases/NIH
Scanning electron micrograph of methicillin-resistant Staphylococcus aureus bacteria (yellow) and a dead human white blood cell (red). Credit: National Institute of Allergy and Infectious Diseases/NIH
Many birds are already raised without antibiotics in the US and elsewhere because of the demands of the market. Since 2016, chicken antibiotic sales decreased by 62% (Dall, 2020). Frequently, the goal of these antibiotic-free (ABF) production programs is to differentiate products in a highly competitive commodity market. The reduction of antibiotic use has been a secondary, generally unintended consequence.
Nevertheless, to meet customer demands for ABF products, antibiotics that are not important to human health but for production (e.g., ionophores) have also been eliminated. In many cases, this has negatively affected growth performance and bird health. As the requirements for production efficiency and welfare standards increase, transitioning from “conventional” to ABF production poses a challenge for everyone involved.
One must never trade animal welfare for reduced antibiotics use, but the need for them can be decreased through improved management practices. Flock health starts with genetics companies selecting birds that are resilient to disease and management challenges and continues all the way to the processing plant. All of the inputs and practices must be optimized in modern poultry production to maintain a high level of performance and animal welfare while reducing reliance on antibiotics.
When antibiotics are not available, attention to detail becomes more decisive. All aspects of production are important, but the most critical stages are those that affect the downstream process. The pullets, breeders, and hatchery require the most meticulous care. Additionally, all production factors must meet the highest quality standards: feed, light, air quality, water quality, litter quality, biosecurity, vaccination, sanitation, nutrition and feeding.
 Antibiotic reduction requires meticulous attention to detail to safeguard animal welfare.
Antibiotic reduction requires meticulous attention to detail to safeguard animal welfare.
ABF production is all about sustainability. For agricultural operations to survive and thrive in the future, one has to move away from the old paradigm of “saving the way to success”. This is not impossible in ABF production, but misses out on the larger picture of long-term profitability, investment in innovation, and system change.
Non-antibiotic feed and water additives are essential resources to support sustainable management. To mention a few, probiotics, prebiotics, toxin binders, organic acids, and phytomolecules are all options for reducing the need for antibiotics based on different modes of action. Phytomolecules, for example, often have antimicrobial properties, some toxin binders can bind bacterial toxins, and pre- and probiotics support the gut flora. There are many kinds of solutions on the market; the key is to find the right ones for your issues.
There is already a large body of literature demonstrating the benefits of alternative or complementary solutions. More importantly, there are already many people that successfully raise birds and other animals without antibiotics. Whenever possible, leverage your professional network and talk to trusted people with unique experiences. Working together, we can build a healthier future for people and animals.
The series that debuts here consists of a set of articles offering professionals a practical overview of poultry production with reduced antibiotic use. The independent expert in charge, starting with the next article in the series, is Dr. TJ Gaydos, who holds a Master’s degree in Avian Medicine and is a diplomate of the American College of Poultry Veterinarians.
Dr. Gaydos works with integrated poultry companies and allied industries, focusing on bird health and antibiotic-free production performance. He has spent his veterinary career working to improve intestinal health, animal welfare, production efficiency, and reduce zoonotic diseases. He works extensively with intestinal health, probiotics and prebiotics, and other non-antimicrobial feed additives.
Topics covered under Dr. Gaydos’s guidance include biosecurity, nutrition, pullet management, hatchery sanitation, gut health, and more. Together they provide an extensive look at the producers’ pain points and potential strategies to maintain bird health while mitigating the need for antibiotics.
AccessScience Editors, “U.S. Bans Antibiotics Use for Enhancing Growth in Livestock.” Access Science. McGraw-Hill Education, January 1, 1970. https://www.accessscience.com/content/u-s-bans-antibiotics-use-for-enhancing-growth-in-livestock/BR0125171.
Dall, Chris. “FDA Reports Another Rise in Antibiotic Sales for Livestock.” FDA Reports Another Rise in Antibiotic Sales for Livestock | International Biosecurity and Prevention Forum, December 16, 2020. https://www.ibpforum.org/news/fda-reports-another-rise-antibiotic-sales-livestock.
Lobanovska, Mariya, and Giulia Pilla . “Penicillin’s Discovery and Antibiotic Resistance: Lessons for the Future?” Yale Journal of Biology and Medicine. 90, no. 1 (March 29, 2017): 135–45.
U.S. Food and Drug Administration. “2020 Summary Report On Antimicrobials Sold or Distributed for Use in Food-Producing Animals” Food and Drug Administration, 2019. https://www.fda.gov/media/144427/download.