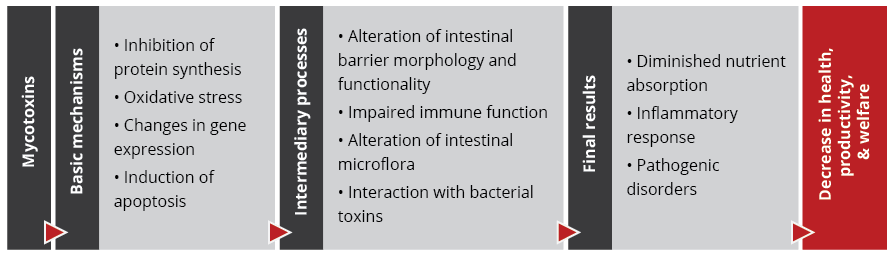
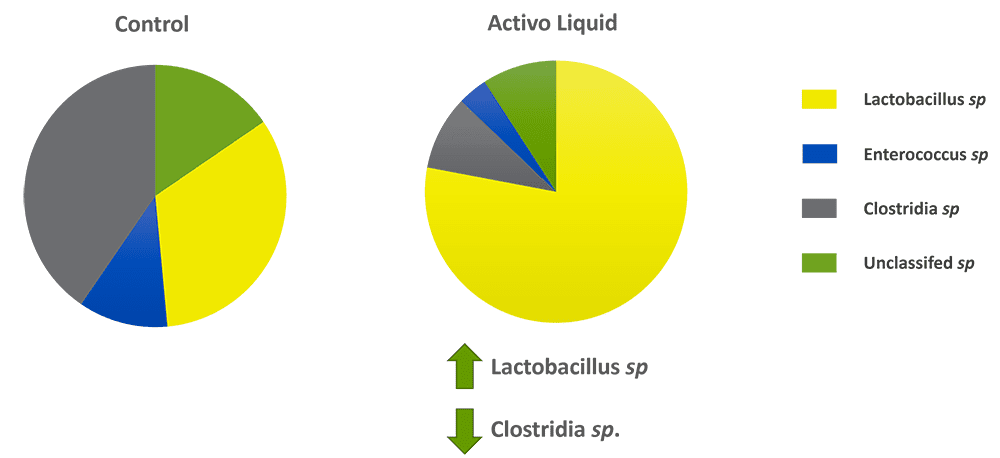Coccidiosis management without increasing antimicrobial resistance – it’s up to us
By Tingting Fan, Regional Technical Manager Poultry, EW Nutrition
Chicken coccidiosis is a common and important disease in poultry production, with an incidence of infection as high as 50-70%. The mortality rates are around 20-30% or higher in highly severe cases. In addition to losses due to mortality, producers lose money due to poor growth as well as decreased meat yield and quality. Additionally, the birds get more susceptible to secondary infections, e.g., necrotic enteritis (Moore, 2016).
The costs caused by coccidiosis in poultry are about 13 billion US $ (Blake, 2020). These costs globally divide into 1 billion costs for prophylaxis/treatment and 12 billion due to performance losses. Until now, only 5% of the prophylaxis costs have been created by natural solutions. That means that there is still a high potential to be tapped.
Natural solutions, unfortunately, are only used by a minority
For a long time, ionophores fitting the classical definition of antibiotics and chemicals were used in coccidia-fighting programs – and contributed to the development of antimicrobial resistance (Nesse et al., 2015). Nowadays, the combination with vaccination in rotation or shuttle programs has reduced this danger, but there is still potential. Meanwhile, some natural solutions are available that can be integrated into coccidiosis-fighting programs. However, producers using natural solutions are still a minority.
For thousands of years, plants have been used in human and veterinary medicine. Before the discovery of antibiotics in 1928, diseases were fought with plants. To regain the effectiveness of antibiotics, using natural solutions for prophylaxis should be once more standard, and the use of antibiotics is the treatment only for critical cases.
How does Eimeria damage broilers
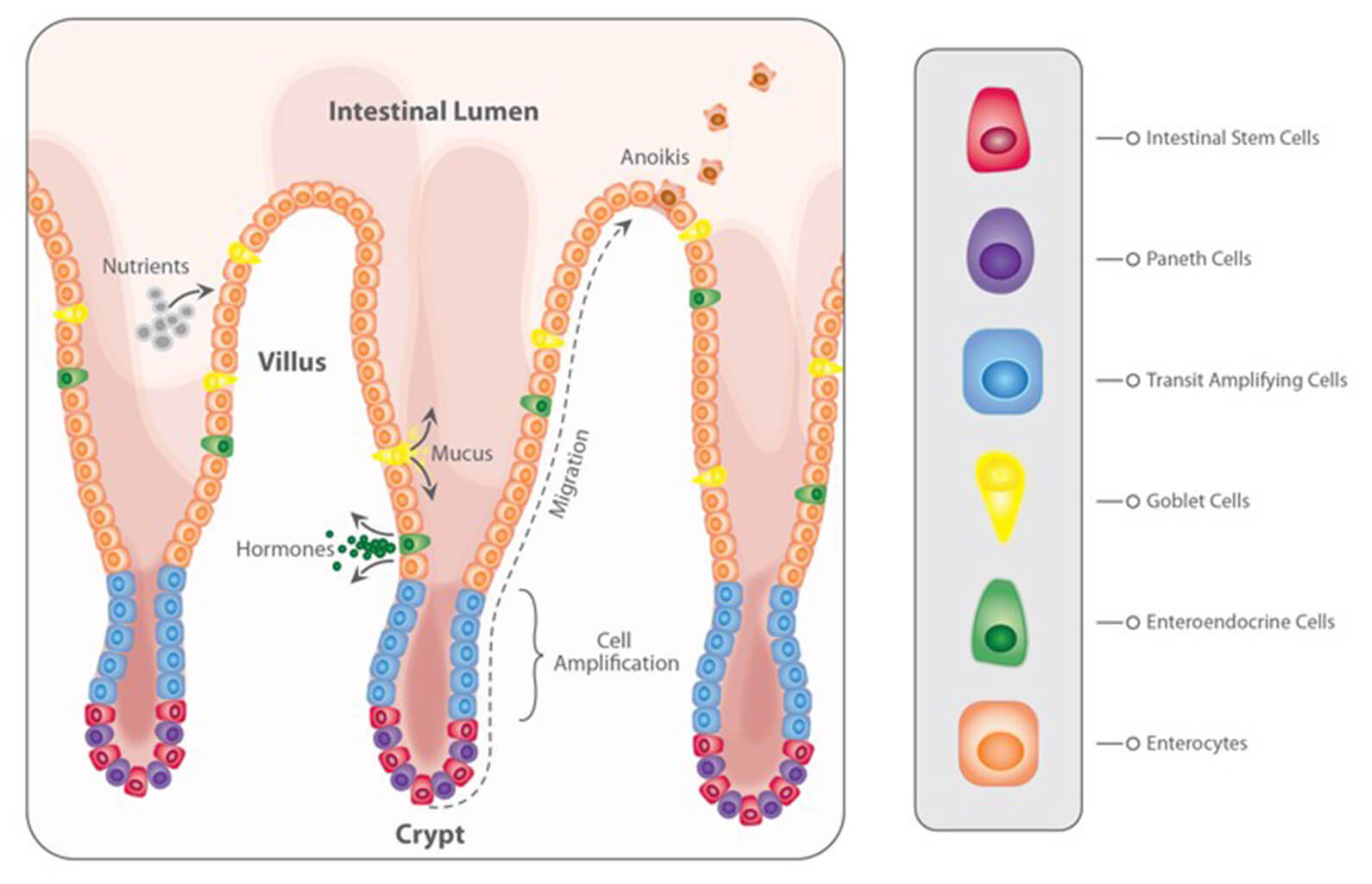
The pathogenic mechanism of coccidia or Eimeria spp. is mainly the massive destruction of host intestinal cells when it reproduces, resulting in severe damage to the intestinal mucosa. On the one hand, the damaged gut wall loses its capability for effective digestion and absorption of nutrients, leading to worse feed conversion and lower weight gain.
On the other hand, this damage reduces the chicken’s immunity and paves the way for other infections, such as necrotic enteritis, and raises mortality.
Table 1:The seven most known Eimeria species in broilers and their main site of occurrence
Eimeria species
Predilection site
E. tenella
Ceca
E. acervulina
Duodenum and prox. jejunum
E. maxima
Central jejunum
E. mitis
Distal jejunum and ileum
E. necatrix
Central jejunum and ceca
E. brunetti
Ileum, entrance of the ceca and rectum
E. praecox
Duodenum and prox. jejunum
Concerning their pathogenicity, for poultry, the Eimeria species must be ordered in the following way: E. necatrix> E. tenella > E. brunetti > E. maxima > E. acervulina > Eimeria mitis, and Eimeria praecox.
Prevention is better than treatment
Thanks to its bi-layered wall with a robust structure, the oocysts of coccidia are extremely resilient. They can survive 4 to 9 months in the litter or soil and are resistant to common disinfectants. Farm personnel and visitors are also important vectors, so good biosecurity practices can reduce the number of oocysts contaminating the premises and help prevent clinical out-brakes. Coccidiosis control in poultry should focus on “prevention” rather than “treatment”, combining biosecurity practices, feed additives, and/or vaccination.
Effective hygiene on the farm is crucial
To prevent coccidia infections, one of the most critical points is hygiene. Biosecurity practices are crucial and include cleaning and disinfection of the poultry houses and their surroundings, pest control and prevention, restriction, control, and management of the entry of personnel, visitors, vehicles, and equipment, among others.
Coccidia oocysts are ubiquitous and survive for a long time, and even effective cleaning and disinfection cannot completely remove them. After a severe outbreak, it is recommended to take drastic biosecurity measures such as flame or caustic soda disinfection to prevent further spread of the disease.
When there are birds in the house, it must be paid attention that the litter is not excessively humid. Litter moisture should be maintained around 25%; turning and replacing moist litter are the best practices to follow. For keeping the litter dry, adequate ventilation and appropriate stocking density are beneficial.
To avoid unnecessary stress and gut health issues, the birds must be fed according to their requirements with high-quality feed so that the animals build up good immunity and resilience.
Coccidiosis can be controlled with effective programs
Anticoccidial drugs were the first means of preventing and controlling coccidiosis in chickens and once achieved very good results. Since Sulfaquinoxaline was found to be effective in the 1850s, about fifty other drugs have been developed for the prevention and control of coccidiosis. Generally, the anticoccidials used for years to prevent the disease can be divided into ionophores and chemicals.
Ionophores, produced as by-products of bacterial fermentation, are technically antibiotics. The great benefits of ionophores are that they kill the parasite before it can infect the bird and thus prevent damage to the host cells. Eimeria species also take a long time to develop resistance to ionophores (Chapman, 2015). Well-established ionophores are products that contain monensin, lasalocid, salinomycin, narasin, or maduramycin; the trade names are Coban/Monensin, Avatec, Coxisstac, Monteban, and Cygro.
Chemicals, these molecules, are produced by chemical synthesis. They differ from each other and ionophores as each one has a unique mode of action against coccidia. In general, they act by interfering with one or more stages of the life cycle of Eimeria, e.g., supplying fake nutrients (Amprolium, Vit. B1) to the parasite, starving them out. The active components here are nicarbazin, amprolium, zoalene, decoquinate, clopidol, robenidine and diclazuril, and the respective trade names Nicarb, Amprol, Zoamix, Deccox, Coyden, Robenz and Clinacox. Eimeria species develop resistance to these chemical molecules; therefore, they must be used carefully and with strict planning. However, cross-resistance does not develop, making them highly valuable in rotation programs.
Vaccination against coccidiosis is accepted by many farmers as a good solution to control coccidiosis in chickens. Vaccination aims to replace resistant field strains with vaccine strains, which are sensitive to anticoccidials. Currently, commercial chicken vaccines are available in natural and attenuated strains; research to obtain safer and more efficient vaccines is also ongoing.
Non-attenuated vaccines are less expensive and make for good immunity, but as they may mildly damage the intestinal epithelium, the risk of necrotic enteritis can increase. On the contrary, attenuated strains – usually “precocious” strains with shorter reproduction cycles, cause less intestinal damage and thus have a lower risk of provoking bacterial or necrotic enteritis. The immunity is like after normal infections; however, you have a controlled epidemiology, fewer coccidiosis outbreaks, and an improved uniformity of the flock.
Phytomolecules-based natural anticoccidials saponins and tannins are natural components that can also help control coccidiosis (e.g., Pretect D, EW Nutrition GmbH). These ingredients act in different ways: the tannins improve the intestinal barrier function locally and systemically. The saponins directly impact the oocysts by preventing their growth, interacting with the cholesterol in the cell membrane (triterpenoid saponin), or hindering further sporulation and causing cell death by causing pores in the cell membrane of the parasite. Altogether, Pretect D promotes the beneficial microbial population and reduces the harmful one, improves the gut barrier function, reduces mucosal inflammation, inhibits growth and replication of Eimeria, preventing their lesions, and fosters birds’ immune response against Eimeria spp.
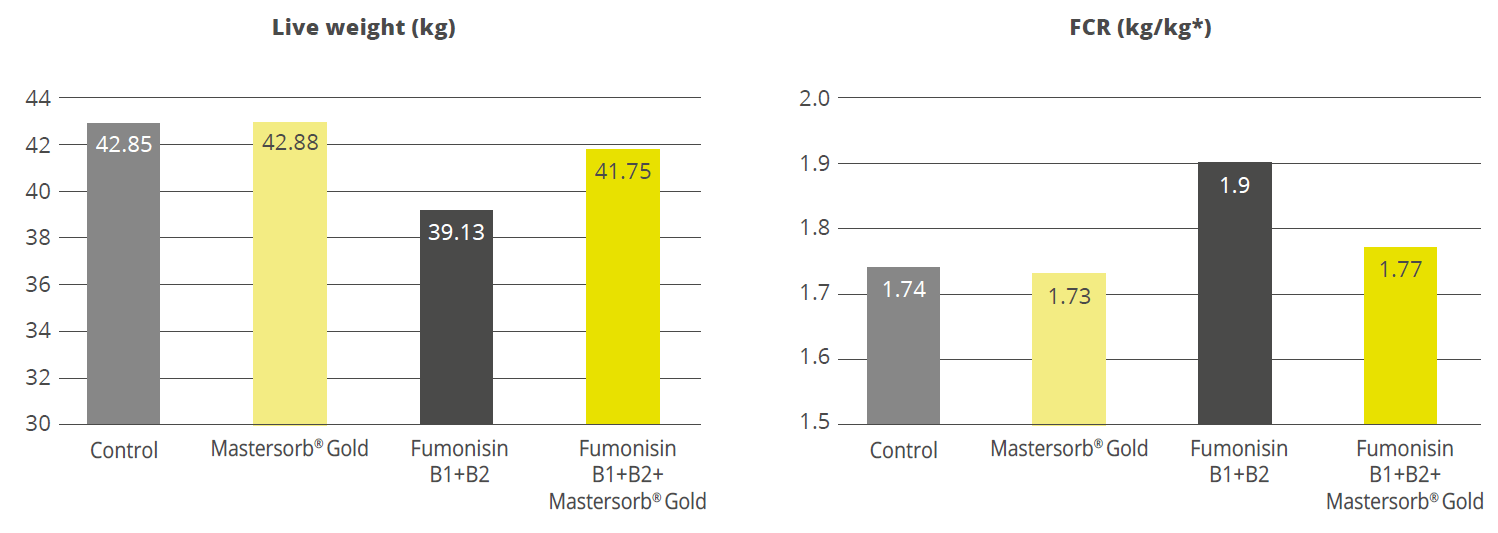
To prove Pretect D’s effectiveness in the reduction of coccidiosis, several trials were conducted. One of the trials was carried out in Poland with 360.000 broilers in commercial conditions. The animals were divided into ten houses, and two cycles were tested. Half of the birds served as control and received Narasin and Nicarbazin in the starter and grower I diet and salinomycin in the grower II diet. The other half also were fed Narasin and Nicarbazin in the starter and grower I diet, but Pretect D @1kg/t in grower II and 0.5kg/t in the finisher diet. The results are shown in figure 1: The application of Pretect D in the grower II and finisher diet decreased the number of oocysts in the droppings more than the application of salinomycin and, therefore, reduced the spreading of coccidiosis. In addition, the performance of the broilers receiving Pretect D was nothing short of the control’s performance showing Pretect as an optimal completion in shuttle or rotation programs (see more HERE).
Figure 1: Reduction of oocysts in the droppings by Pretect D
Managing coccidiosis without promoting antimicrobial resistance is not easy, but feasible
Coccidiosis is a challenge aggravated by our current high level of production. Tools such as ionophores, chemicals, but also vaccines, and natural products are available to fight coccidiosis. However, due to the high probability of resistance development, these tools must be used carefully and in structured programs. The phytomolecules-based product Pretect D gives the possibility to reduce antimicrobial resistance as part of programs against coccidiosis.
References upon request
Effective phytomolecules combine superior processing stability and strong action in the animal
By Dr. Inge Heinzl, Editor, and Dr. Ruturaj Patil, Global Product Manager – Phytogenics, EW Nutrition
For millennia, plants have been used for medicinal purposes in human and veterinary medicine and as spices in the kitchen. Since the ban of antibiotic growth promoters in 2006 by the European Union, they also came into focus in animal nutrition. Due to their digestive, antimicrobial, and gut health-promoting characteristics, they seemed an ideal alternative to compensate for the reduced use of antibiotics in critical periods such as brooding, feed change or gut-related stress.
To optimize the benefits of phytomolecules, it is crucial that
the phytomolecules levels are standardized for consistent results and synergy
they show the highest stability during stringent feed processing; being often highly volatile substances, they should not get lost at high temperatures and pressure
the phytomolecules are preferably completely released and available in the animal to achieve the best effectiveness.
First step: Standardized phytomolecules
Essential oils and other phytogenics are sourced from plants. The composition of the plants substantially depends on genetic dissimilarity within accessions, plant origin, the site conditions, such as weather, soil, community, and harvest time, but also sample drying, storage, and extraction processes (Sadeh et al., 2019; Yang et al., 2018; Ehrlinger, 2007). For example, the oil extracted from thyme can contain between 22 and 71 % of the relevant phenol thymol (Soković et al., 2009; Shabnum and Wagay, 2011; Kowalczyk et al., 2020).
Modern technology enables the production of standardized phytomolecules with the highest degree of purity and lowest possible batch-to-batch variation for high-quality products. It also offers increased environmental and economic sustainability due to reliable and cost-effective sourcing technology.
Second step: Selection of the most suitable phytomolecules
Phytomolecules have different primary characteristics. Some support digestion (Cho et al., 2006, Oetting, 2006; Hernandez, 2004); others act against pathogens (Sienkiewitz et al., 2013; Smith-Palmer et al., 1998; Özer et al., 2007) or are antioxidants (Wei and Shibamoto, 2007; Cuppett and Hall, 1998). To optimize gut health in animal production, one of the main promising mechanisms is reducing pathogens while promoting beneficial microbes. The decrease of pathogens in the gut not only decreases the risk of enteritis incidence but also eliminates the inconvenient competitors for feed.
In order to find out the best combination serving the intended purpose, a high number of different phytomolecules need to be evaluated concerning their structure, chemical properties, and biological activities first. Availability and costs of the substances are further factors to consider. With the selection of the most suitable phytomolecules, different mixtures are produced and tested for their effectiveness. Here, it is essential to concern synergistic or antagonistic effects.
For an effective and efficient blend of phytomolecules, many steps of selection and tests are necessary – and as a result, possibly only a few mixtures can meet the requirements.
Third step: Protecting the ingredients
Many phytomolecules are inherently highly volatile. So, only having a standardized content of phytogenics in the product can not ensure the full availability of phytomolecules when used through animal feed. Some parts of the ingredients might already get lost in the feed mill due to the stringent feed hygienization process followed by feed millers to reduce pathogenic load. The heating is a significant challenge for the highly-volatile components in a phytomolecule-based product. So, protecting these phytomolecules becomes imperative to guarantee that the phytomolecules put into the feed will reach the animal.
A delicate balancing act is required to ensure the availability and activity of phytomolecules at the right site in the gut. The phytomolecules must not get lost during feed processing but must also be released in the intestine. A carrier with capillary binding of the phytomolecules together with a protective coating can be one of the available effective solutions. It protects the ingredients during feed processing, and ensures the release in the animal.
Study shows excellent stability of Ventar D under challenging conditions
Ventar D is a latest generation phytomolecule-based solution for gut health optimization introduced by EW Nutrition, GmbH. A scientific study was conducted to compare the stability of Ventar D, in the pelleting process, with two leading phytogenics competitor feed supplements.
For this trial, feed with the different added phytogenic feed supplements had to undergo a conditioning and pelletization process. The active ingredients were analyzed before and after the pelletization process. All phytogenic feed supplements under testing were added to standard broiler feed at the producer’s recommended inclusion rate. The tests took place under conditioning times of 45, 90, and 180 seconds and pelleting temperatures of 70, 80, and 90°C (158, 176, and 194°F). After cooling, triplicate samples were collected and analyzed. The respective marker substance was analyzed through gas chromatography/mass spectrometry (GC/MS) analysis to measure the recovery rate in the finished feed. The phytomolecule content of the mash feed (before pelletization) found by the laboratory was used as a baseline and set to 100% recovery. The recovery rates of the pelleted feed were evaluated relative to this baseline.
The results are presented in figure 1. Ventar D showed the highest stability of active ingredients with recovery rates of 90% at 70°C/45 sec. or 80°C/90 sec and 84% at 90°C/180 sec. The modern production technology used for Ventar D ensures that the active ingredients are well protected throughout the pelletization process.
Figure 1: Phytomolecule stability under processing conditions, relative to mash baseline (100%)
Another trial was conducted in a feed mill in the US. For this trial, ten samples were collected from different batches of mash feed where Ventar D was added at 110g/t. Conditioning of the mash feed was at 87.8°C (190°F) for 6 minutes and 45 seconds. After the pelleting process, ten samples from the pelleted feed were collected from the continuous flow with a 5 min gap between the samplings to determine Ventar D’s recovery.
The average recovery achieved for Ventar D was 92%.
Trials show improved growth performance
Initial trials showed Ventar D’s complete release in digestion models. To examine the benefit in in-vivo conditions, Ventar D was tested in broilers at an inclusion rate of 100 g/MT.
Several in vitro studies proved the antimicrobial activity of Ventar D. One test also confirms that Ventar D could exhibit differential antimicrobial activity by having stronger activity against common enteropathogenic bacteria while sparing the beneficial ones (Heinzl, 2022). Moreover, Ventar D’s antioxidant and anti-inflammatory activity support better gut barrier functioning. Better gut health leads to higher growth performance and improved feed conversion, which could be demonstrated in several trials with broilers (figures 2 and 3). In the tests, a group fed Ventar D was compared to either a control group with no such feed supplement or groups supplied with competitor products at the recommended inclusion rates.
Compared to a negative control group, the Ventar D group consistently showed a higher average daily gain of 0.3-4.1 g (0.5-8.5 %) and a 3-4 points better feed conversion. Compared to competitor products, Ventar D provided 1-1.7 g (2-3 %) higher average daily gain and a 3 points better /1 point higher FCR than competitors 2 and 1.
Figure 2: Average daily gain (g) – results of several trials conducted with broilers
Figure 3: FCR – results of several trials conducted with broilers
Standardization and new technologies for higher profitability
Several in vitro and in vivo studies proved that Ventar D takes “phytomolecules’ power” to the next level: Combining standardized phytomolecules and optimal active ingredient protection leads to superior product stability during feed processing. The higher amount of active ingredients arriving in the gut improves gut health and increases the production performance of the animals. Ventar D shows how we can use phytomolecules more effectively and benefit from higher farm profitability.
References:
Cho, J. H., Y. J. Chen, B. J. Min, H. J. Kim, O. S. Kwon, K. S. Shon, I. H. Kim, S. J. Kim, and A. Asamer. “Effects of Essential Oils Supplementation on Growth Performance, IGG Concentration and Fecal Noxious Gas Concentration of Weaned Pigs”. Asian-Australasian Journal of Animal Sciences 19, no. 1 (2005): 80–85. https://doi.org/10.5713/ajas.2006.80.
Cuppett, Susan L., and Clifford A. Hall. “Antioxidant Activity of the Labiatae”. Advances in Food and Nutrition Research 42 (1998): 245–71. https://doi.org/10.1016/s1043-4526(08)60097-2.
Ehrlinger, M. “Phytogenic Additives in Animal Nutrition.” Dissertation, Veterinary Faculty of the Ludwig Maximilians University, 2007.
Heinzl, I. “Efficient Microbiome Modulation with Phytomolecules”. EW Nutrition, August 30, 2022. https://ew-nutrition.com/pushing-microbiome-in-right-direction-phytomolecules/.
Hernández, F., J. Madrid, V. García, J. Orengo, and M.D. Megías. “Influence of Two Plant Extracts on Broilers Performance, Digestibility, and Digestive Organ Size.” Poultry Science 83, no. 2 (2004): 169–74. https://doi.org/10.1093/ps/83.2.169.
Kowalczyk, Adam, Martyna Przychodna, Sylwia Sopata, Agnieszka Bodalska, and Izabela Fecka. “Thymol and Thyme Essential Oil—New Insights into Selected Therapeutic Applications.” Molecules 25, no. 18 (2020): 4125. https://doi.org/10.3390/molecules25184125.
Lindner, , U. “Aromatic Plants – Cultivation and Use.” Düsseldorf: Teaching and Research Institute for Horticulture Auweiler-Friesdorf, 1987.
Oetting, Liliana Lotufo, Carlos Eduardo Utiyama, Pedro Agostinho Giani, Urbano dos Ruiz, and Valdomiro Shigueru Miyada. “Efeitos De Extratos Vegetais e Antimicrobianos Sobre a Digestibilidade Aparente, O Desempenho, a Morfometria Dos Órgãos e a Histologia Intestinal De Leitões Recém-Desmamados.” Revista Brasileira de Zootecnia 35, no. 4 (2006): 1389–97. https://doi.org/10.1590/s1516-35982006000500019.
Sadeh, Dganit, Nadav Nitzan, David Chaimovitsh, Alona Shachter, Murad Ghanim, and Nativ Dudai. “Interactive Effects of Genotype, Seasonality and Extraction Method on Chemical Compositions and Yield of Essential Oil from Rosemary (Rosmarinus Officinalis L”.).” Industrial Crops and Products 138 (2019): 111419. https://doi.org/10.1016/j.indcrop.2019.05.068.
Shabnum, Shazia, and Muzafar G. Wagay. “Essential Oil Composition of Thymus Vulgaris L. and Their Uses”. Journal of Research & Development 11 (2011): 83–94.
Sienkiewicz, Monika, Monika Łysakowska, Marta Pastuszka, Wojciech Bienias, and Edward Kowalczyk. “The Potential of Use Basil and Rosemary Essential Oils as Effective Antibacterial Agents.” Molecules 18, no. 8 (2013): 9334–51. https://doi.org/10.3390/molecules18089334.
Smith-Palmer, A., J. Stewart, and L. Fyfe. “Antimicrobial Properties of Plant Essential Oils and Essences against Five Important Food-Borne Pathogens”. Letters in Applied Microbiology 26, no. 2 (1998): 118–22. https://doi.org/10.1046/j.1472-765x.1998.00303.x.
Soković, Marina, Jelena Vukojević, Petar Marin, Dejan Brkić, Vlatka Vajs, and Leo Van Griensven. “Chemical Composition of Essential Oils of Thymus and Mentha Species and Their Antifungal Activities”. Molecules 14, no. 1 (2009): 238–49. https://doi.org/10.3390/molecules14010238.
Wei, Alfreda, and Takayuki Shibamoto. “Antioxidant Activities and Volatile Constituents of Various Essential Oils.” Journal of Agricultural and Food Chemistry 55, no. 5 (2007): 1737–42. https://doi.org/10.1021/jf062959x.
Yang, Li, Kui-Shan Wen, Xiao Ruan, Ying-Xian Zhao, Feng Wei, and Qiang Wang. “Response of Plant Secondary Metabolites to Environmental Factors”. Molecules 23, no. 4 (2018): 762. https://doi.org/10.3390/molecules23040762.
Özer, Hakan, Münevver Sökmen, Medine Güllüce, Ahmet Adigüzel, Fikrettin Şahin, Atalay Sökmen, Hamdullah Kiliç, and Özlem Bariş. “Chemical Composition and Antimicrobial and Antioxidant Activities of the Essential Oil and Methanol Extract of Hippomarathrum Microcarpum (Bieb.) from Turkey”. Journal of Agricultural and Food Chemistry 55, no. 3 (2007): 937–42. https://doi.org/10.1021/jf0624244.
Efficient microbiome modulation with phytomolecules
By Dr. Inge Heinzl, Editor, EW Nutrition
From day 1, young animals are confronted with the pathogens of their environment. Feed and feed ingredients also significantly increase exposure to microbes. This article will look closely at three critical bacteria in poultry production. The trials of phytomolecules-based products shared in this article prove the unique benefit of lowering harmful pathogens while simultaneously sparing health-promoting microbes. The targeted selection of the blend’s phytomolecules contributes to this distinctive mode of action.
E. coli can be valuable… and dangerous
E.coli are commensal bacteria that usually belong to the natural gut flora. However, there are several E. coli strains that, due to certain virulence factors, can cause disease. These bacteria are called avian pathogenic E. coli or APEC. The disease ‘Colibacillosis’ can occur in different forms:
Omphalitis – a noncontagious infection of the navel and/or yolk sac in young poultry
peritonitis – inflammatory response on “internal laying” (yolk material in the peritoneum)
salpingitis – inflammation of the oviduct
cellulitis – discoloration and thickening of the skin, inflammation of the subcutaneous tissues
synovitis – lameness with swollen joints
coligranuloma (Hjärre disease) – lesions similar to tuberculosis, not of economic importance
meningitis, and
septicemia or blood poisoning.
Since some of the E. coli strains can sometimes be transmitted vertically to offspring, it is crucial to keep the pathogenic pressure in the parent generation as low as possible (Mc Dougal, 2018).
Due to the, mostly in young chicks, common use of antibiotics, E. coli strains resistant to ß-lactam antibiotics (ESBL-producing E. coli) or fluoroquinolones (e.g., Enrofloxacin) have developed.
Clostridium perfringens: the cause of necrotic enteritis
Clostridium perfringens belong to the normal caecal flora. However, its overgrowth in the intestine is linked to necrotic enteritis, causing estimated losses of up to USD 6 billion yearly in global poultry production, which corresponds to USD 0.0625 per bird (Wade and Keyburn, 2015). Necrotic enteritis can occur in a clinical and a subclinical form.
In the case of clinical necrotic enteritis, the birds suffer from diarrhea resulting in wet litter and increased flock mortality of up to 1 % per day (Ducatelle and Van Immerseel, 2010). Mortality rates sometimes sum up to 50 % (Van der Sluis, 2013). If birds die without clinical signs, it may be peracute necrotic enteritis.
The subclinical version, however, is more critical. Due to the lack of symptoms, it often remains undetected and, therefore, not treated. Mainly through the impaired utilization of feed, representing 65-75 % of the total costs in broiler production, subclinical necrotic enteritis permanently impacts production efficiency (Heinzl et al., 2020).
Salmonella enterica: a zoonosis relevant for birds and humans
Most concerning in (non-typhoid) salmonellosis is that it can be transferred to humans. The transmission occurs via direct contact with an infected animal, consuming contaminated animal products such as meat or eggs, contact with infected vectors (insects or pets) or contaminated equipment, or cross-contamination in the kitchen. Frozen or raw chicken products, as well as the eggs, are frequent causes of animal-origin Salmonella infections in humans.
Salmonella is the more critical the younger the birds. If the hatching eggs already carry salmonellae, the hatchability dwindles. During their first weeks of life, infected chicks show higher mortality and systemic infections.
Adult animals usually do not die from salmonellosis; often, the infection remains unnoticed. During an acute salmonella outbreak, the animals might show weakness and diarrhea. They lose weight, resulting in decreased egg production in layers.
Trials with phytomolecules show promising results
To check if phytomolecules-based products can effectively influence gut flora, a product specially designed for gut health (Ventar D) was tested for its antimicrobial activity. Additionally, the extent to which the same blend impacted the beneficial bacteria, such as Lactobacilli, was evaluated.
Trial 1: phytomolecules act against E. coli and Salmonella enterica
The in vitro study using the agar dilution method was conducted at a German laboratory.
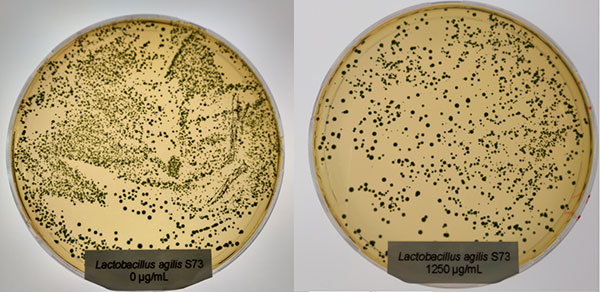
The bacteria (Salmonella typhimurium and ESBL-producing E. coli) stored at -80°C were reactivated by cultivating them on Agar Mueller Hinton overnight. After this incubation, some colonies were picked and suspended in 1 ml 0.9% NaCl solution. 100 µl of the suspension were pipetted and evenly spread (plate spread technique) on new Agar Mueller Hinton containing different concentrations of a phytomolecules-based product (Ventar D): 0 µg/mL – control; 500 µg/mL; 900 µg/mL; 1.250 µg/mL and 2.500 µg/mL. After 16-20 h incubation at 37°C, growth was evaluated. The results can be seen in pictures 1 and 2:
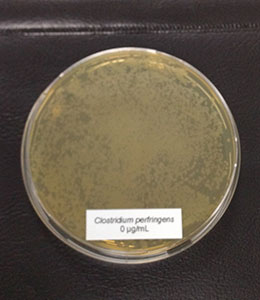
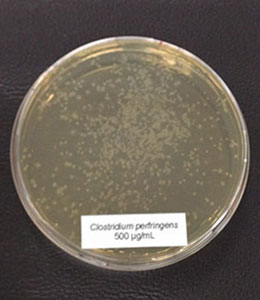
Figure 1: E. coli exposed to different concentrations of Ventar D (upper row from left to right: control 0 µg/ml, 500 µg/ml, 900 µg/ml; lower row from left to right: 1250 µg/ml and 2500 µg/ml)
E. coli colonies exposed to 900 µg/mL of Ventar D’s phytogenic formulation were smaller than the control colonies. At 1250 µg/mL, fewer colonies were detected, and at 2500 µg/mL, growth couldn’t be seen anymore.
The salmonella colonies showed a similar picture; however, the reduction could be seen from a concentration of 1.250 µg/ml of Ventar D onwards (picture 2).
Figure 2: Salmonella enterica exposed to different concentrations of Ventar D (upper row from left to right: control 0 µg/ml, 500 µg/ml, 900 µg/ml; lower row from left to right: 1250 µg/ml and 2500 µg/ml)
Trial 2: Phytomolecules inhibit Clostridium perfringens and spare Lactobacilli
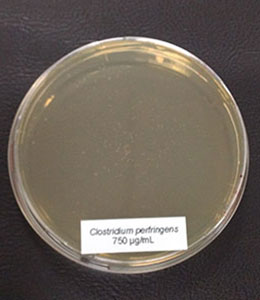
In this trial, the bacteria (Clostridium perfringens,Lactobacillus agilis S73, and Lactobacillus plantarum) were cultured under favorable conditions (RCM, 37°C, anaerobe for Clostr. perfr., and MRS, 37°C, 5 % CO2 for Lactobacilli) and exposed to different concentrations of Ventar D (0 µg/ml – control, 500 µg/ml, 750 µg/ml, and 1000 µg/ml).
The results are shown in figures 3a-d.
Figure 3a: control, 0 µg/ml
Figure 3b: 500 µg/ml
Figure 3c: 750 µg/ml
Figure 3d: 1000 µg/m
In the case of Clostridium perfringens, a significant reduction of colonies could already be observed at a concentration of 500 µg/ml of Ventar D. At 750 µg/ml, only a few colonies remained. At a Ventar D concentration of 1000 µg/ml, Clostridium perfringens could no longer grow.
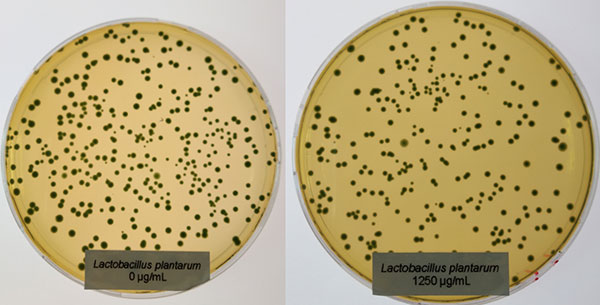
In contrast to Clostridium, the Lactobacilli showed a different picture: only at the higher concentration (1250 µg/ml of Ventar D), Lactobacillus plantarum and Lactobacillus agilis S73 showed a slight growth reduction (figures 4 and 5).
Figure 4: Lactobacillus plantarum exposed to 0 (left) and 1250 µg/ml (right) of Ventar D
Figure 5: Lactobacillus agilis S73 exposed to 0 (left) and 1250 µg/ml (right) of Ventar D
Improve gut health by positively influencing the intestinal flora
The experiments show that even at lower concentrations, phytomolecules impair the growth of harmful bacteria while sparing the beneficial ones. Phytomolecule-based products can be regarded as a valuable tool for controlling relevant pathogens in poultry and influencing the microflora composition in a positive way.
The resulting better gut health is the best precondition to reducing antibiotics in animal production.
Improve health and productivity in breeders with phytomolecules
By Dr.Inge Heinzl, Editor, and Technical Team, EW Nutrition
Careful management of the breeders isa must to get their best reproductive efficiency. In today’s hatching egg production, factors such as stress, inflammation, body weight, and altered mating behaviorlead to decreased performance, meaning fewer hatchable eggs and, therefore, fewer day-old chicks per hen (Grandhaye, 2020). The use of antibiotics to increase performance in farm animals is no longer allowed in many countries, and, since it may lead to the development of resistance, it is also not recommended. So, also in breeders, alternatives are requested to maintain animal health, welfare, and a high level of performance.
Optimal gut health is the cornerstone for breeder performance
As the organ responsible for digestion of the incoming feed, the absorption of nutrients, and the defense of the organism against pathogens or toxins, a healthy gut is a pre-condition for optimal performance (Shini and Bryden, 2021). A healthy gut, according to Bailey (2018), has optimally developed gut tissues, a well-functioning gut immune system, and well-balanced gut microbiota. It shows efficient functionality in terms of digestion and absorption and protects the organism against harmful agents.
The gut directly or indirectly provides the elements for egg production
Efficient feed digestion and absorption of nutrients are essential for the breeder hen to obtain the “material” for maintenance, growth, and egg production. Gut health is crucial since dysbacteriosis and diarrhea, characteristics of gut health challenges, increase dirty eggs, creating favorable conditions for pathogens to enter the egg and infect the embryo.
Egg yolks consist of water (70%), proteins (10%), and lipids (20%). The yolk lipids are lipoproteins rich in triglycerides, built up in the liver and transported to the ovary. Cholesterol carried via lipoproteins to the egg yolk is also built up there, thus showing the importance of the liver in egg production. The gut plays a crucial role in protecting the liver from damage, constituting a barrier against harmful pathogens and toxins, potentially passing into the bloodstream and reaching this vital organ.
Phytomolecules support performance in different ways
Phytomolecules, are an excellent tool to support gut health and animal performance. Phytomolecules are plant-derived secondary metabolites that exert insect-attracting or defensive functions in the plant. They are used in their natural but also nature-identical forms in humans and animals to exert their digestive, immune-modulating, antimicrobial effects.
Phytomolecules support gut health by balancing the gut microbiome
Diverse examples can be found in the scientific literature, where phytomolecules improve the gut microbiome, resulting in better performance of layer and breeder hens. This support happens in two ways:
Promoting beneficial bacteria
Rabelo-Ruiz and co-workers (2021), asserted that adding garlic and onion extracts to the diet of layers led to more eggs with a bigger size, accompanied by an increase in Lactococci in the ileum and Lactobacilli in the cecum. Another example is provided by Park et al. (2016). When supplementing the diet of layers with a fermented phytogenic feed additive, egg production and weight raised with increasing dosage of the additive, and a higher number of Lactobacilli could be observed in the cecum.
Phytomolecules can promote the growth of certain beneficial bacteria and therefore act like prebiotics. As these changes took place in the lower gut, they assumed an improved digestibility of the feed.
Lowering pathogenic bacteria
In the study by Park et al. (2016) and in an in vitro study by Ghazanfari et al. (2019),E. coli in the cecum was reduced.
According to Burt (2007b), several essential oils / phytomolecules, amongst them, carvacrol, thymol, eugenol, and cinnamaldehyde, are effective against pathogens such as Listeria, Salmonella, E. coli, Shigella, and Staphylococcus. The hydrophobic essential oils can partition the lipids of the cell membranes. The resulting permeability of the membrane enables the leakage of cell content.
Changing virulence factors
Another mode of action is the change of virulence factors. Carvacrol, e.g., is known to decrease the motility of Campylobacter jejuni (Van Alphen et al., 2012); oregano and thyme oil reduced the motility of E. coli by inhibiting the synthesis of flagellin (Burt, 2007a). Vidanarachchi et al. (2005) mentioned that the hydrophobicity of microbes increases when some plant extracts are present, affecting their virulence characteristics. Also, the inhibition of defense measures such as efflux pumps in Gram-negative bacteria has been researched (Savoia, 2012).
Phytomolecules support gut health by improving digestion
For many years, phytomolecules have been studied and known for their digestive characteristics. In poultry and other animals, they influence feed digestion in two main ways.
Stimulating enzyme secretion
Platel and Srinivasan (2004) described different spices promoting not only the salivary flow, gastric juice and bile secretion but also the stimulation of the activity of enzymes such as pancreatic lipase, amylase, and proteases in rats. Hashemipour et al. (2013) saw the same effect in broilers supplemented with carvacrol and thymol in the diet. Research has also concluded on a higher nutrient digestibility: Hernandez et al. (2004) and Basmacioğlu Malayoğlu, 2010 noticed that supplementing plant extracts or essential oils improved apparent whole-tract and ileal digestibility of different nutrients.).
Maintaining gut integrity and enlarging the digestion area
An intact gut with a large area for digestion guarantees optimal utilization of nutrients. Different researchers found that adding plant extracts or essential oils (Khalaji et al., 2011; Ghazanfari et al., 2015; Chowdhury et al., 2018) promotes intestinal gut morphology, reflected in higher villi and deeper crypts, which might lead to higher nutrient absorption.
Concerning gut integrity, thymol and carvacrol showed protecting effects and mitigated gut lesions in broilers challenged with C. perfringens (Du et al., 2016). Probably, the lower pathogenic pressure due to the antimicrobial activity of phytogenic substances leads to minor damage to the gut wall and, in the end, to better absorption of the nutrients.
Phytomolecules mitigate the effects of stress
Environmental stress in breeders may decrease performance: the heat-stress-induced disruption of the tight junctions often leads to higher gut permeability, poor nutrient absorption, and higher electrolyte and water secretion (Abdelli, 2021). Sahin et al. (2010) achieved a linear improvement in egg production in quails when applying two doses of green tea catechin.
Cold-stressed layers also reacted positively to supplementation of oregano essential oil, improving egg production compared to a non-supplemented control (Migliorini, 2019).
Positive influence of phytomolecules results in higher performance
As described, phytomolecules improve gut health and support the animal in multiply ways, allowing better utilization of resources for growth and production. Literature provides many articles showing the promoting effects of these substances on the performance of layers or breeders, some of them summarized in Table 1.
Table 1: Benefits of phytomolecules in layers and breeders
Compounds
Reference
Main effects: Improved egg weight, egg mass, and higher hen-day-egg production
In-feed and in-water phytomolecules-based products show efficacy
Much of the research done with phytomolecules focuses on essential oils (with variable inclusions of the active compounds or on single plant extracts. EW Nutrition is a research-driven company proposing phytomolecule-based solutions for the animal production industry. These products combine selected, synergistically acting phytomolecules to achieve optimal results.
EW Nutrition has tested the combined use of
a microencapsulated blend of phytomolecules (Activo) for the feed and designed to maintain a good gut-health status during the whole life-cycle of the breeders, and
Activo Liquid, a liquid combination of phytomolecules and organic acids, which is conveniently applied on the farm via the waterline.
A trial conducted at the University of Central Queensland (Australia) showed that phytomolecules enhance beneficial bacteria such as Lactobacilli and, on the other hand, repress harmful bacteria such as Clostridium perfringens.
For the trial, caecal microbiota of layers was used. They were grown with and without Activo Liquid in vitro, and the changes in microbiota were monitored.
Result: The in vitro study clearly shows that Activo Liquid increases the number of lactobacilli and decreases clostridia and Enterococcus sp.
Cie Chart
Figure 1: Shifting intestinal balance with phytomolecules
2.Three field trials with Activo Liquid showed an increased laying rate in breeders
Many operations started testing phytomolecules in a farm-application-based program to reaffirm the gut health-improving activity of phytomolecules in broiler breeder performance. Especially the flexibility of assisting animals through the water for drinking during stress periods makes phytomolecules an optimal tool to support gut health.
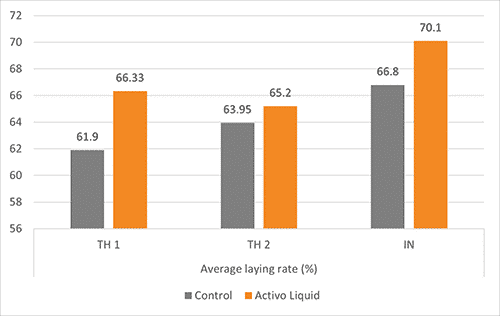
Two broiler breeder farms in Thailand (TH1 and TH2) and one grandparent farm in India (IN) are good examples of the effectiveness of phytomolecules. On each farm, the birds were always divided into two groups. Besides the standard management, feed, and water, one group got 200 ml Activo Liquid per 1,000 L of water. The periods when the birds received Activo in the water differed:
TH1 & TH2: 5 days per week, during weeks 24 – 32
IN: 5 days per week, every third week from weeks 18 to 24 and every fourth week from 28 to 36
The trials lasted for 9 weeks (Thailand 1 and 2) and 30 weeks (India).
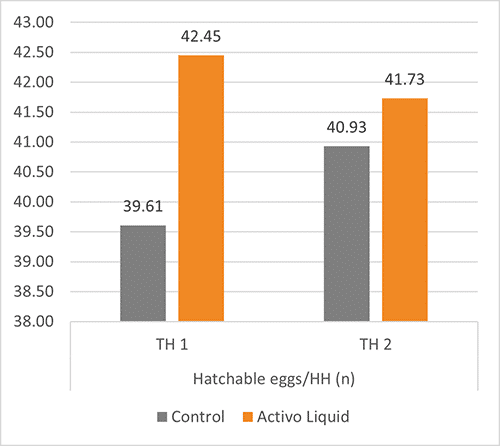
The results are shown in figure 2. The animals supplemented with Activo Liquid showed an up to 4.4 % higher laying rate and up to three more hatchable eggs per hen housed.
Figure 2+3: Results of three trials conducted In Asia concerning laying rate and hatchable eggs
3. Customers tell about lower breeder mortality and more DOCs due to phytomolecules
The benefits of a tailored phytomolecule program have been demonstrated in several broiler breeder operations worldwide. For example, a combination of the in-feed (Activo) and the in-water solution (Activo Liquid) was tested in the Middle East. For the study, 75,000 23-weeks-old broiler breeders were divided into groups: 4 houses with the program, and 6 houses served as control (standard feed and water). The program, tailored to customer needs, was designed as follows:
AC+AL group:
Activo 100 g/ton of feed during the whole trial (weeks 23-41) +
Activo Liquid 250 ml/1000 L water, four days per week, weeks 23-30.
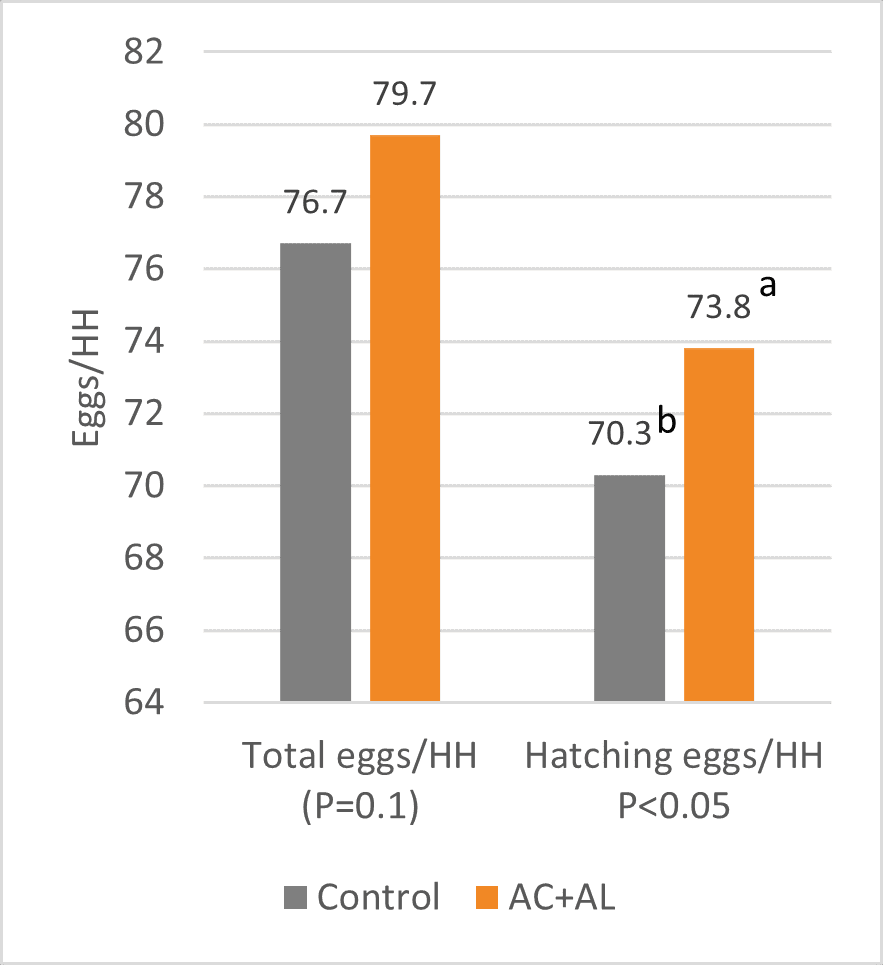
As a result, the peak and average laying rates were higher for the flocks with the program, and laying persistency was also higher. This allowed for a significant difference of 3 total and 3.5 hatching eggs/hen housed at week 41. In both cases, an increase equivalent to 5 % compared to the control group (figure 4) could be observed.
Figure 4: Total eggs and hatching eggs per hen housed
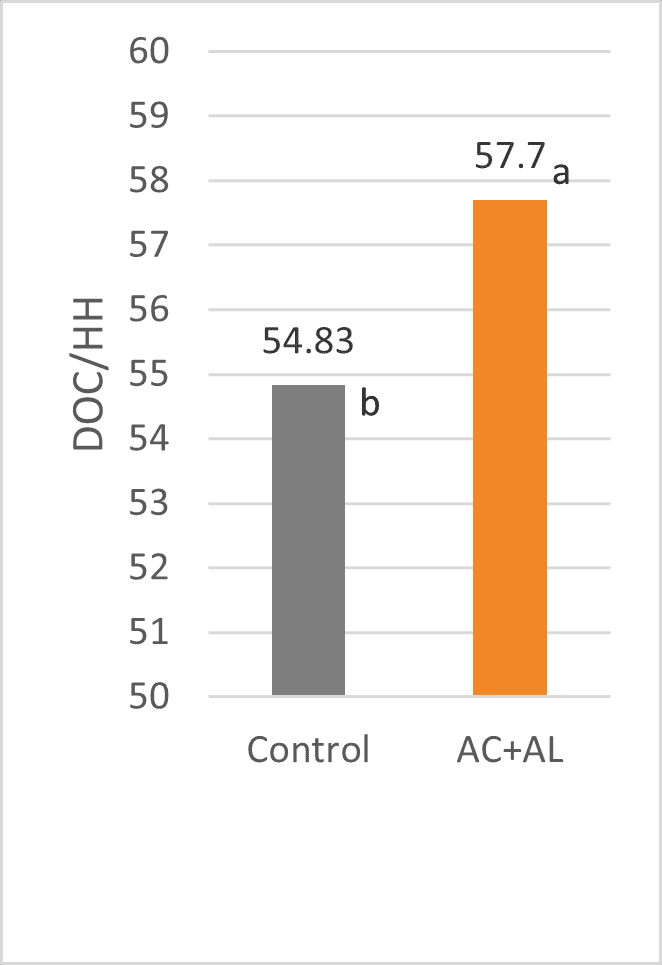
As fertility and hatchability were similar for both groups, the 5 % increase in hatching eggs resulted in a 5 % higher number of day-old chicks per hen housed (figure 5).
Figure 5: Number of DOSs per hen housed
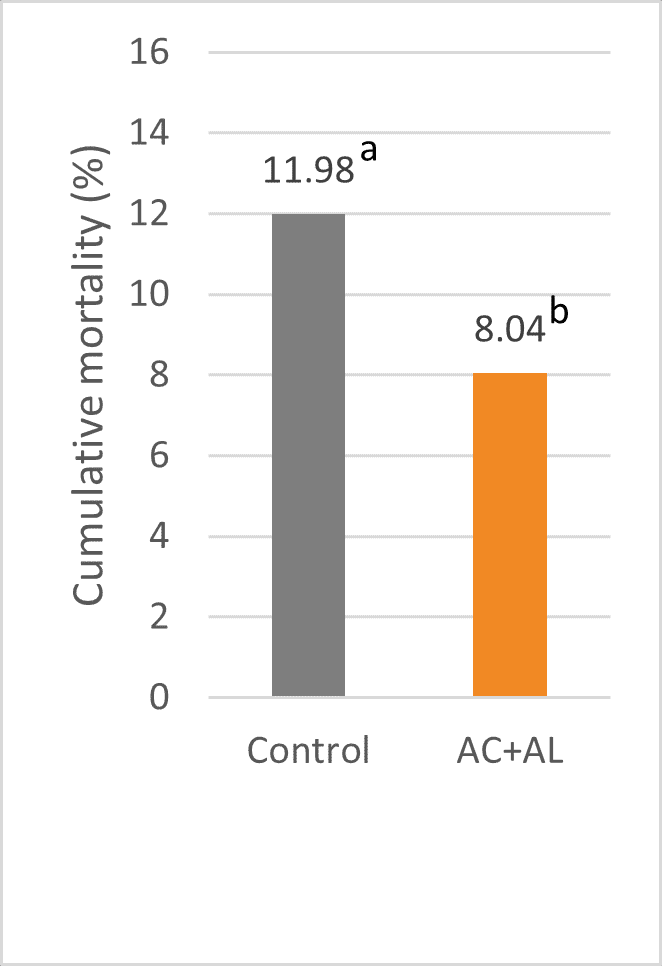
It must be mentioned that during the trial period, at 28 weeks of age, an NDV outbreak was diagnosed on the farm, which negatively impacted the overall results. However, this impact was reduced in the groups receiving the phytomolecule-based products, which also was reflected in a lower mortality rate (figure 6).
Figure 6: Cumulative mortality rate wk 41
4. Scientific trial shows that Activo can increase post-peak productivity in breeders
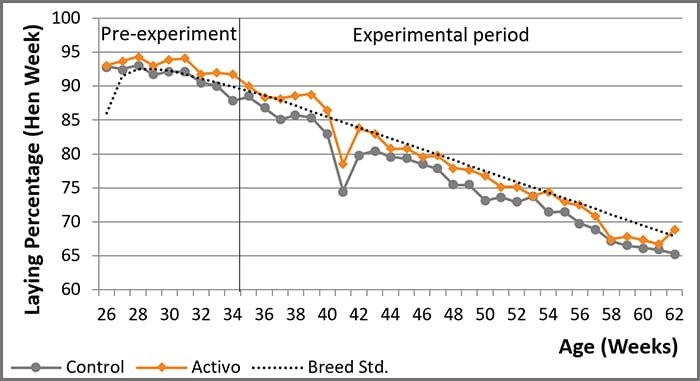
When thinking about the use of phytomolecules, most broiler breeder operations would like to consider scientific trial results in this type of animal. For EW Nutrition, it is crucial to accurately evaluate every product that reaches a market. Thus several scientific trials with broiler breeders have been performed. For one of them, Hubbard breeders (JA57 females with 80 M77 males) were divided into 2 treatments, having 5 replicate pens for each. The experiment started after the peak production period, at 34 weeks of age, and ended at week 62. To make the trial fair, the production data of 6 (pre-experimental) weeks was used to allocate the pens for each treatment, resulting in two (statistically) similar groups.
The control group was fed the standard mash diet. For the Activo group, 100g Activo/MT was added to the diet.
With Activo, breeders kept their high productivity after the peak, while the control group showed a steady decline from breed target values. During the experiment, Activo supplemented birds produced 3.6 more eggs than control birds (P=0.06) while consuming a similar amount of feed. As a result, a lower feed consumption per egg produced was achieved (169.9 vs. 173.6 g/egg, respectively).
As the dietary treatment did not influence hatchability, the 3.6 extra eggs resulted in 2.9 extra day-old chicks per hen during the post-peak period, showing a positive return.
Phytomolecules as gut health and performance promoters– antibiotics can be reduced!
With their gut health-promoting activity, phytomolecules support breeders to better utilize nutrients. They can be invested for maintenance and the production of hatchable eggs, obtaining good quality day-old chicks.
References
Abad, Paloma, Natalia Arroyo-Manzanares, Juan J. Ariza, Alberto Baños, and Ana M. García-Campaña. “Effect of Allium Extract Supplementation on Egg Quality, Productivity, and Intestinal Microbiota of Laying Hens.” Animals 11, no. 1 (2020): 41. https://doi.org/10.3390/ani11010041.
Abdel-Wareth, A.A.A., and J.D. Lohakare. “Effect of Dietary Supplementation of Peppermint on Performance, Egg Quality, and Serum Metabolic Profile of Hy-Line Brown Hens during the Late Laying Period.” Animal Feed Science and Technology 197 (2014): 114–20. https://doi.org/10.1016/j.anifeedsci.2014.07.007.
Abdel-Wareth, A.A.A., Z.S.H. Ismail, and K.-H. Südekum. “Effects of Thyme and Oregano on Performance and Egg Quality Characteristics of Laying Hens.” World’s Poultry Sci. Journal, Supplement, 69 (2013): 1–6.
Abdelli, Nedra, David Solà-Oriol, and José Francisco Pérez. “Phytogenic Feed Additives in Poultry: Achievements, Prospective and Challenges.” Animals 11, no. 12 (2021): 3471. https://doi.org/10.3390/ani11123471.
Abou-Elkhair, Reham, Shaimaa Selim, and Eman Hussein. “Effect of Supplementing Layer Hen Diet with Phytogenic Feed Additives on Laying Performance, Egg Quality, Egg Lipid Peroxidation and Blood Biochemical Constituents.” Animal Nutrition 4, no. 4 (2018): 394–400. https://doi.org/10.1016/j.aninu.2018.05.009.
Akbari, Mohsen, Mehran Torki, and Keyomars Kaviani. “Single and Combined Effects of Peppermint and Thyme Essential Oils on Productive Performance, Egg Quality Traits, and Blood Parameters of Laying Hens Reared under Cold Stress Condition (6.8 ± 3 °C).” International Journal of Biometeorology 60, no. 3 (2015): 447–54. https://doi.org/10.1007/s00484-015-1042-6.
Al-Harthi, M.A. “Responses of Laying Hens to Different Levels of Amoxicillin, Hot Pepper or Green Tea and Their Effects on Productive Performance, Egg Quality and Chemical Composition of Yolk and Blood Plasma Constituents.” Egypt. Poult. Sci. 24, no. IV (2004): 845–68.
Al-Yasiry, A.R.M., B. Kiczorowska, W. Samolińska, E. Kowalczuk-Vasilev, and D. Kowalczyk-Pecka. “The Effect of Boswellia Serrata Resin Diet Supplementation on Production, Hematological, Biochemical and Immunological Parameters in Broiler Chickens.” Animal 11, no. 11 (2017): 1890–98. https://doi.org/10.1017/s1751731117000817.
Ali, Akhtar, Eric N. Ponnampalam, Gamini Pushpakumara, Jeremy J. Cottrell, Hafiz A. Suleria, and Frank R. Dunshea. “Cinnamon: A Natural Feed Additive for Poultry Health and Production—a Review.” Animals 11, no. 7 (2021): 2026. https://doi.org/10.3390/ani11072026.
Bailey, Richard. “Maintaining Gut Health without Antibiotics.” Poultry World, April 19, 2021. https://www.poultryworld.net/health-nutrition/maintaining-gut-health-without-antibiotics/.
Basmacioğlu Malayoğlu, H., Ş. Baysal, Z. Misirlioğlu, M. Polat, H. Yilmaz, and N. Turan. “Effects of Oregano Essential Oil with or without Feed Enzymes on Growth Performance, Digestive Enzyme, Nutrient Digestibility, Lipid Metabolism and Immune Response of Broilers Fed on Wheat–Soybean Meal Diets.” British Poultry Science 51, no. 1 (2010): 67–80. https://doi.org/10.1080/00071660903573702.
Burt, Sara A., Ruurd van der Zee, Ad P. Koets, Anko M. de Graaff, Frans van Knapen, Wim Gaastra, Henk P. Haagsman, and Edwin J. Veldhuizen. “Carvacrol Induces Heat Shock Protein 60 and Inhibits Synthesis of Flagellin in Escherichia Coli O157:H7.” Applied and Environmental Microbiology 73, no. 14 (2007a): 4484–90. https://doi.org/10.1128/aem.00340-07.
Burt, Sara Ann. “Antibacterial Activity of Essential Oils: Potential Applications in Food.” Thesis, S. Burt, 2007b.
Bölükbasi, S. Canan, and M. Kuddusi Erhan. “Effect of Dietary Thyme (Thymus Vulgaris) on Laying Hens Performance and Escherichia Coli (E. Coli) Concentration in Feces.” International Journal of Natural and Engineering Sciences 1, no. 2 (2007): 55–58.
Chowdhury, Subrata, Guru Prasad Mandal, Amlan Kumar Patra, Pawan Kumar, Indranil Samanta, Saktipada Pradhan, and Arup Kumar Samanta. “Different Essential Oils in Diets of Broiler Chickens: 2. Gut Microbes and Morphology, Immune Response, and Some Blood Profile and Antioxidant Enzymes.” Animal Feed Science and Technology 236 (2018): 39–47. https://doi.org/10.1016/j.anifeedsci.2017.12.003.
Dilawar, Muhammad Ammar, Hong Seok Mun, Dhanushka Rathnayake, Eun Ju Yang, Ye Seul Seo, Hyeoung Seog Park, and Chul Ju Yang. “Egg Quality Parameters, Production Performance and Immunity of Laying Hens Supplemented with Plant Extracts.” Animals 11, no. 4 (2021): 975. https://doi.org/10.3390/ani11040975.
Du, Encun, Weiwei Wang, Liping Gan, Zhui Li, Shuangshuang Guo, and Yuming Guo. “Effects of Thymol and Carvacrol Supplementation on Intestinal Integrity and Immune Responses of Broiler Chickens Challenged with Clostridium Perfringens.” Journal of Animal Science and Biotechnology 7, no. 1 (2016). https://doi.org/10.1186/s40104-016-0079-7.
Franciosini, Maria Pia, Patrizia Casagrande-Proietti, Claudio Forte, Daniela Beghelli, Gabriele Acuti, Dario Zanichelli, Alessandro dal Bosco, Cesare Castellini, and Massimo Trabalza-Marinucci. “Effects of Oregano (Origanum Vulgarel.) and Rosemary (Rosmarinus Officinalis l.)Aqueous Extracts on Broiler Performance, Immune Function and Intestinal Microbial Population.” Journal of Applied Animal Research 44, no. 1 (2015): 474–79. https://doi.org/10.1080/09712119.2015.1091322.
Ghazanfari, S, Z Mohammadi, and M Adib Moradi. “Effects of Coriander Essential Oil on the Performance, Blood Characteristics, Intestinal Microbiota and Histological of Broilers.” Revista Brasileira de Ciência Avícola 17, no. 4 (2015): 419–26. https://doi.org/10.1590/1516-635×1704419-426.
Grandhaye, Jeremy, Veronique Douard, Ana Rodriguez-Mateos, Yifan Xu, Alex Cheok, Antonella Riva, Rodrigo Guabiraba, et al. “Microbiota Changes Due to Grape Seed Extract Diet Improved Intestinal Homeostasis and Decreased Fatness in Parental Broiler Hens.” Microorganisms 8, no. 8 (2020): 1141. https://doi.org/10.3390/microorganisms8081141.
Hashemipour, H., H. Kermanshahi, A. Golian, and T. Veldkamp. “Effect of Thymol and Carvacrol Feed Supplementation on Performance, Antioxidant Enzyme Activities, Fatty Acid Composition, Digestive Enzyme Activities, and Immune Response in Broiler Chickens.” Poultry Science 92, no. 8 (2013): 2059–69. https://doi.org/10.3382/ps.2012-02685.
Hernández, F., J. Madrid, V. García, J. Orengo, and M.D. Megías. “Influence of Two Plant Extracts on Broilers Performance, Digestibility, and Digestive Organ Size.” Poultry Science 83, no. 2 (2004): 169–74. https://doi.org/10.1093/ps/83.2.169.
Khalaji, S., M. Zaghari, K.H. Hatami, S. Hedari-Dastjerdi, L. Lotfi, and H. Nazarian. “Black Cumin Seeds, Artemisia Leaves (Artemisia Sieberi), and Camellia L. Plant Extract as Phytogenic Products in Broiler Diets and Their Effects on Performance, Blood Constituents, Immunity, and Cecal Microbial Population.” Poultry Science 90, no. 11 (2011): 2500–2510. https://doi.org/10.3382/ps.2011-01393.
Khan, Sohail Hassan, Muhammad Ashraf Anjum, Abida Parveen, Tabinda Khawaja, and Naeem Mehmood Ashraf. “Effects of Black Cumin Seed (Nigella Sativa) on Performance and Immune System in Newly Evolved Crossbred Laying Hens.” Veterinary Quarterly 33, no. 1 (2013): 13–19. https://doi.org/10.1080/01652176.2013.782119.
Kim, Duk Kyung, Hyun S. Lillehoj, Sung Hyen Lee, Erik P. Lillehoj, and David Bravo. “Improved Resistance to Eimeria Acervulina Infection in Chickens Due to Dietary Supplementation with Garlic Metabolites.” British Journal of Nutrition 109, no. 1 (2012): 76–88. https://doi.org/10.1017/s0007114512000530.
Kothari, Damini, Woo-Do Lee, Kai-Min Niu, and Soo-Ki Kim. “The Genus Allium as Poultry Feed Additive: A Review.” Animals 9, no. 12 (2019): 1032. https://doi.org/10.3390/ani9121032.
Lutful Kabir, S. M. “Avian Colibacillosis and Salmonellosis: A Closer Look at Epidemiology, Pathogenesis, Diagnosis, Control and Public Health Concerns.” International Journal of Environmental Research and Public Health 7, no. 1 (2010): 89–114. https://doi.org/10.3390/ijerph7010089.
Migliorini, Marcos J., Marcel M. Boiago, Lenita M. Stefani, Aline Zampar, Lenilson F. Roza, Mauricio Barreta, Alessandra Arno, et al. “Oregano Essential Oil in the Diet of Laying Hens in Winter Reduces Lipid Peroxidation in Yolks and Increases Shelf Life in Eggs.” Journal of Thermal Biology 85 (2019): 102409. https://doi.org/10.1016/j.jtherbio.2019.102409.
Mohiti-Asli, Maziar, and Moein Ghanaatparast-Rashti. “Comparison of the Effect of Two Phytogenic Compounds on Growth Performance and Immune Response of Broilers.” Journal of Applied Animal Research 45, no. 1 (2017): 603–8. https://doi.org/10.1080/09712119.2016.1243119.
Nadia, L.,, Radwan, R.A. Hassan, E.M. Qota, and H.M. Fayek. “Effect of Natural Antioxidant on Oxidative Stability of Eggs and Productive and Reproductive Performance of Laying Hens.” International Journal of Poultry Science 7, no. 2 (2008): 134–50. https://doi.org/10.3923/ijps.2008.134.150.
Nadia, L.,, Radwan, R.A. Hassan, E.M. Qota, and H.M. Fayek. “Effect of Natural Antioxidant on Oxidative Stability of Eggs and Productive and Reproductive Performance of Laying Hens.” International Journal of Poultry Science 7, no. 2 (2008): 134–50. https://doi.org/10.3923/ijps.2008.134.150.
Park, Jae Hong, Tae Hwa Song, and Inho Kim. “Egg Production, Egg Quality, and Cecal Microbial Populations of Layers fed Diets Supplemented with Fermented Phytogenic Feed Additive.” TURKISH JOURNAL OF VETERINARY AND ANIMAL SCIENCES 40 (2016): 660–66. https://doi.org/10.3906/vet-1512-55.
Platel, Kalpana, and K. Srinivasan. “Digestive Stimulant Action of Spices: a Myth or Reality?” Indian J Med Res. 119, no. 5 (2004): 167–79.
Puvača, Nikola, Erinda Lika, Sonila Cocoli, Tana Shtylla Kika, Vojislava Bursić, Gorica Vuković, Mirela Tomaš Simin, Aleksandra Petrović, and Magdalena Cara. “Use of Tea Tree Essential Oil (Melaleuca Alternifolia) in Laying Hen’s Nutrition on Performance and Egg Fatty Acid Profile as a Promising Sustainable Organic Agricultural Tool.” Sustainability 12, no. 8 (2020): 3420. https://doi.org/10.3390/su12083420.
Rabelo-Ruiz, Miguel, Juan José Ariza-Romero, María Jesús Zurita-González, Antonio Manuel Martín-Platero, Alberto Baños, Mercedes Maqueda, Eva Valdivia, Manuel Martínez-Bueno, and Juan Manuel Peralta-Sánchez. “Allium-Based Phytobiotic Enhances Egg Production in Laying Hens through Microbial Composition Changes in Ileum and Cecum.” Animals 11, no. 2 (2021): 448. https://doi.org/10.3390/ani11020448.
Ren, Hao, Wilfried Vahjen, Temesgen Dadi, Eva-Maria Saliu, Farshad Goodarzi Boroojeni, and Jürgen Zentek. “Synergistic Effects of Probiotics and Phytobiotics on the Intestinal Microbiota in Young Broiler Chicken.” Microorganisms 7, no. 12 (2019): 684. https://doi.org/10.3390/microorganisms7120684.
Sahin, K., C. Orhan, M. Tuzcu, S. Ali, N. Sahin, and A. Hayirli. “Epigallocatechin-3-Gallate Prevents Lipid Peroxidation and Enhances Antioxidant Defense System via Modulating Hepatic Nuclear Transcription Factors in Heat-Stressed Quails.” Poultry Science 89, no. 10 (2010): 2251–58. https://doi.org/10.3382/ps.2010-00749.
Shini, S., and W. L. Bryden. “Probiotics and Gut Health: Linking Gut Homeostasis and Poultry Productivity.” Animal Production Science, 2021. https://doi.org/10.1071/an20701.
van Alphen, Lieke B., Sara A. Burt, Andreas K. Veenendaal, Nancy M. Bleumink-Pluym, and Jos P. van Putten. “The Natural Antimicrobial Carvacrol Inhibits Campylobacter Jejuni Motility and Infection of Epithelial Cells.” PLoS ONE 7, no. 9 (2012). https://doi.org/10.1371/journal.pone.0045343.
Vidanarachchi, J.K., L.L. Mikkelsen, I. Sims, P.A. Iji, and M. Choct. “Phytobiotics: Alternatives to Antibiotic Growth Promoters in Monogastric Animal Feeds.” Recent Advances in Animal Nutrition in Australia 15 (2005): 131–44.
Wang, Xiao-cui, Xiao-hong Wang, Jing Wang, Hao Wang, Hai-jun Zhang, Shu-geng Wu, and Guang-hai Qi. “Dietary Tea Polyphenol Supplementation Improved Egg Production Performance, Albumen Quality, and Magnum Morphology of Hy-Line Brown Hens during the Late Laying period1.” Journal of Animal Science 96, no. 1 (2018): 225–35. https://doi.org/10.1093/jas/skx007.
Zhu, Nianhua, Jun Wang, Longfei Yu, Qiman Zhang, Kai Chen, and Baosheng Liu. “Modulation of Growth Performance and Intestinal Microbiota in Chickens Fed Plant Extracts or Virginiamycin.” Frontiers in Microbiology 10 (2019). https://doi.org/10.3389/fmicb.2019.01333.
How can you compensate an activated immune system in piglets?
By Technical Team, EW Nutrition
As pig production specialists, we understand that our animals are under constant challenge during their life. Challenges can be severe or moderate, correlated to several factors – such as, for instance, stage of production, environment, and so on – but they will always be present. To be successful, we need to understand how to counter these challenges and support the healthy development of our pigs.
Factors for successful pig production
For years we have been increasing our understanding of how to formulate diets to support a healthy intestine through the optimal use of the supplied nutrients. Functional proteins, immune-related amino acids, and fiber are now applied worldwide for improved pig nutrition.
What lies beyond formulation adjustments?
However, pig producers have also realized that these nutritional strategies alone are not always fully efficient in preventing an “irritation” of the immune system and/or in preventing diseases from happening.
Immune nutrition is gaining a strong foothold in pig production, and the body of research and evidence grows richer every year. At the same time, we see genetics continually evolving and bringing production potential to increasingly higher levels. We are also constantly increasing our understanding of the importance of farm and feed management, as well as biosecurity in this process.
Finally, the importance of a stable microflora is now uncontested. Especially around weaning, a stable microflora is necessary to prevent the proliferation of pathogens such as E.coli bacteria. Such pathogens can degrade the lysine (the main amino acid for muscle protein production) we have added to our formulations, rendering it useless.
Single molecules (or additives) are able to support the development of gut microflora, boost its integrity, and therefore help the animals use “traditional nutrients” in a more effective way.
The impact of immune system activation on the performance of pigs
Animal performance is influenced by complex processes, from metabolism to farm biosecurity. Environmental conditions, diet formulation and feed management, and health status, among others, directly affect the amount of the genetic potential that animals can effectively express.
Among these so-called non-genetic variables, health status is one of the most decisive factors for the optimal performance from a given genotype. Due to the occurrence of (sub-) clinical diseases, the inflammatory process can be triggered and may result in a decrease in weight gain and feed efficiency.
Not so long ago, pig producers believed that a maximized immune response would always be ideal for achieving the best production levels. However, after decades spent researching what this “maximized immune response” could mean to our pigs, studies from different parts of the globe proved that an activated immune system could negatively affect animal performance. The perception is nowadays common sense within the global pig production industry.
That understanding led us to increasingly search for production systems that will yield the best conditions for the pigs. This means minimum contact with pathogens, reduced stress factors, and therefore a lower need for an activated immune system.
How immune system stimulation works
The immune system has as main objective to identify the presence of antigens – substances that are not known to the body – and protect the body from these “intruders”. The main players among these substances are bacteria and viruses. However, some proteins can also trigger an immunological reaction. Specific immune cells are responsible for the transfer of information to the other systems of the body so that it can respond adequately. This response from the immune system includes metabolic changes that can affect the demand for nutrients and, therefore, the animals’ growth.
The stimulation of the immune system has three main metabolic consequences:
behavioral responses
direct connection with the endocrine system and regulation of the secretions
release of leukocytes, cytokines, and macrophages
In general, the immune system responds to antigens, releasing cytokines that activate the cellular (phagocytes) and humoral components (antibody), resulting in a decreased feed intake and an increased body temperature/heat production.
When feed formulation is concerned, possibly even more important is to understand that the activation of the immune system leads to a change in the distribution of nutrients. The basal metabolic rate and the use of carbohydrates will have completely different patterns in such an event. For instance, some glucose supplied through the feed follows its course to peripheral tissues; however, part of the glucose is used to support the activated immune system. As a consequence, the energy requirement of the animal increases.
Protein synthesis and amino acid utilization also change during this process. There is a reduction of body protein synthesis and an increased rate of degradation. The nitrogen requirement increases because of the higher synthesis of acute-phase proteins and other immunological cells.
However, increased lysine levels in the diets will not always help the piglets compensate for this shift in the protein metabolism. According to Shurson & Johnston (1998), when the immune system is activated, there is further deamination of amino acids and increased urinary excretion of nitrogen. Therefore we need to understand better which amino acids must be supplied in a challenging situation.
In pigs, the gastrointestinal tract is, to a large extent, responsible for performance. This happens because the gut is the route for absorption of nutrients, but also a reservoir of hundreds of thousands of different microorganisms – including the pathogenic ones.
Understanding Gut Health
Gut health and its meaning have been the topic of several peer-reviewed articles in the last few decades (Adewole et al., 2016, Bischoff, 2011, Celi et al., 2017, Jayaraman and Nyachoti, 2017, Kogut and Arsenault, 2016, Moeser et al., 2017, Pluske, 2013). Despite the valuable body of knowledge accumulated on the topic, a clear and widely-accepted definition is still lacking. Kogut and Arsenault (2016) define it in the title of their paper as “the new paradigm in food animal production”. The authors explain it as the “absence / prevention / avoidance of disease so that the animal is able to perform its physiological functions in order to withstand exogenous and endogenous stressors”.
In a recently published paper, Pluske et al. (2018) add to the above definition that gut health should be considered in a more general context. They describe it “a generalized condition of homeostasis in the GIT, with respect to its overall structure and function”. The authors add to this definition that gut health in pigs can be compromised even when no clinical symptoms of disease can be observed. Every stressful factor can undermine the immune response of pigs and, therefore, the animals’ performance.
All good information on this topic leads us to the conclusion that, without gut balance, livestock cannot perform as expected. Therefore, balance is the objective for which we formulate our pigs’ feed.
Current nutritional strategies for a stable gut microbiota
Feeding: quality of raw materials
The photos included here were taken in the field and show that taking action against this reality is a must for keeping animals healthy.
Much of this action is related to farm management. The most effective way to minimize such situations is to implement a strict control system in the feed production sites, including controlling raw material quality.
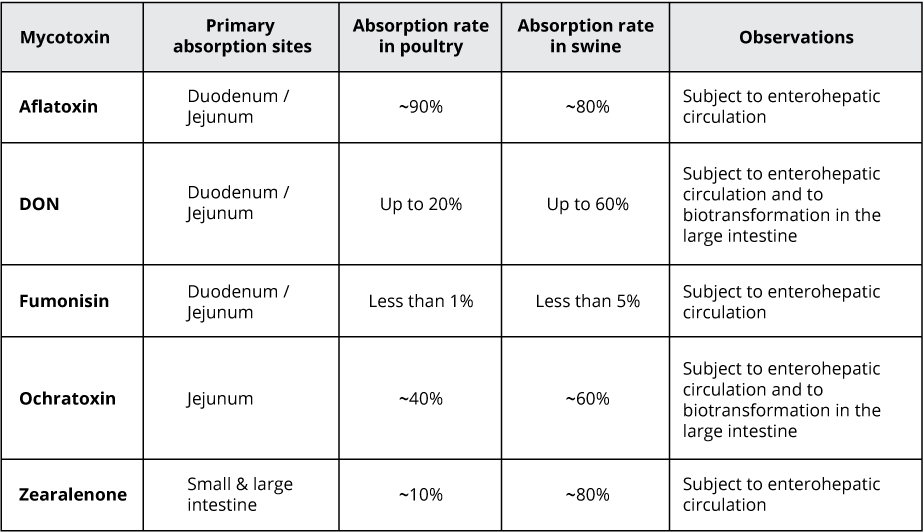
Additives can be used to improve the safety of raw materials. As already extensively discussed, everything that goes into the intestine of the animals will affect gut health and performance. Therefore, the potential harmful load of mycotoxins should be taken into account. Besides careful handling at harvest and the proper storage of grains, mycotoxin binders can be applied to further decrease the risk of mycotoxin contamination.
Figure 1. Grain storage in a home pig farm
Figure 2. Feed mixer in a home mixer pig farm
The effect of nutrition on microflora: commercial weaning diet after focusing on gut health
The gut-health-focused formulation of diets must take into account the following essentials:
decrease of gut pH
gut wall integrity
minimization of (pathogenic) microbial growth
microflora modulation with consequently improved colonization resistance
Gut pH
A lower pH in the stomach slows the passage rate of the feed from the stomach to the small intestine. A longer stay of the feed in the stomach potentially increases the digestion of starch and protein. The secretion of pancreatic juices stimulated by the acidic stomach content will also improve the digestion of feed in the small intestine.
For weaned pigs, it is essential that as little as possible of the substrate will reach the large intestine and be fermented. Pathogens take advantage of undigested feed to proliferate. Lowering these “nutrients” will decrease the risk of bacterial overgrowth.
The same is true where protein sources and their levels are concerned. It is essential to reduce protein content as much as possible and preferably use synthetic (essential) amino acids. The application of such sources of amino acids has been proven long ago, and yet in some cases, it is still not fully utilized. Finally, using highly digestible protein sources should, at this point, be a matter of mere routine.
All these strategies have the same goal: the reduction of undigested substances in the gut. Additionally, the reduction of the protein levels can also decrease the costs of the diets.
Further diet adjustments
Further diet adjustments, such as increasing the sulfur amino acids (SAA) tryptophan and threonine to lysine ratio, must also be considered (Goodband et al., 2014; Sterndale et al., 2017). Although the concept of better balancing tryptophan and threonine are quite clear among nutritionists, SAA are sometimes overestimated. Sulfur amino acids are the major amino acids in proteins related to body maintenance, but not so high in muscle proteins. Therefore, the requirement of SAA must also be approached differently. Unlike lysine, the requirements of SAA tend to be higher in immunologically stimulated animals (Table 1).
Pig weight (kg)
ISA*
SID Lysine (%)
SAA (%)
SAA:Lys
9
High
1,34
0,64
0,48
Low
1,07
0,59
0,55
14
High
1,22
0,62
0,51
Low
0,99
0,57
0,58
Table 1. Effect of the immune system activation on the demand for lysine and sulfur amino acids in pigs (Stahly et al., 1998)
*ISA – immune system activation
Vitamins and minerals are classic nutrients to be considered when formulating gut health-related diets. Maybe not so extensive as the amino acids and protein levels, these nutrients have, however, been found to carry benefits in challenging situations. In the past several years, a lot was published on the requirements of pigs facing an activation of the immune system. Stahly et al. (1996) concluded that when the immune system is activated, the phosphorous requirements change.
Parameters
ISA*
High
Low
Feed intake (g/d)
674
833
Weight gain (g/d)
426
566
Available P (%)
0,45
0,65
Table 2. Effect of the immune system activation on the performance and phosphorous requirements of pigs (Stahly et al., 1998)
*ISA – immune system activation
Another example is vitamin A. It is involved in the function of macrophages and neutrophils. Vitamin A deficiency decreases the migratory and phagocytic abilities of the immune cells. A lower antibody production is observed in vitamin A deficiency as well. Furthermore, vitamin A is an important factor in mucosal immunity, because this vitamin plays a role in lymphocyte homing in the mucosa (Duriancik et al., 2010).
Phytomolecules: key additives to support gut health
Phytomolecules are currently considered one of the top alternatives to in-feed antibiotics for pigs worldwide. Programs sponsored by the European Union are once more evaluating the effectiveness of these compounds as part of a strategy to produce sustainable pigs with low or no antibiotic use. The EIP-Agri (European Innovation Partnership “Agricultural Productivity and Sustainability”) released a document with suggestions to lower the use of antibiotics in feed by acting in three areas:
improving pig health and welfare
changing attitudes and human habits
finding specific alternatives to antibiotics
Under the last topic, the commission recommends plant-based feed additives to be further examined.
Antibiotics have been used for many years for supporting performance in animal production, especially in critical moments. The mode of action consists of the reduction of pathogen proliferation and inflammation processes in the digestive tract. These (soon-to-be-) banned compounds therefore reduce the activation of the immune system, helping keep pigs healthy through a healthy gastrointestinal tract. As potential alternatives to antibiotic usage, phytomolecules should be able to do the same.
The mode of action of phytomolecules
Antimicrobial
Most phytomolecules used nowadays aim to control the number and type of bacteria in the gut of animals. According to Burt (2004), the antimicrobial activity of phytomolecules is not the result of one specific mode of action, but a combination of effects on different targets of the cell. This includes disruption of the membrane by terpenoids and phenolics, metal chelation by phenols and flavonoids, and protective effects against viral infections for certain alkaloids and coumarins (Cowan, 1999).
Digestion support
The antimicrobial efficacy is one of the most important activities of secondary plant compounds, but it also impacts digestion. Windisch et al. (2008) states that growth-promoting agents decrease immune defense stress during critical situations. They increase the intestinal availability of essential nutrients for absorption, thus promoting the growth of the animal.
Indeed, phytomolecules are a good tool for stabilizing the gut microbiota. But more can be expected when adding this class of additives into your formulation and/or farm operations. Mavromichalis, in his book “Piglet Nutrition Notes – Volume 2”, brings attention to the advantages of using phytomolecules such as capsaicin, which is often related to increased feed intake. Recent research has demonstrated that capsaicin increases the secretion of digestive enzymes that may result in enhanced nutrient digestibility. According to Mavromichalis, this can lead to a better feed conversion rate as more nutrients are available to the animal. Indirectly, this also helps control the general bacterial load in the gut.
Antioxidant support
This results from the polyphenols’ capacity to act as metal-chelators, free radical scavengers, hydrogen donators, and inhibitors of the enzymatic systems responsible for initiating oxidation reaction. Furthermore, they can act as a substrate for free radicals such as superoxide or hydroxyl, or intervene in propagation reactions.
This variety of benefits explains at least partially the high level of interest in this group of additives for pigs under challenging conditions. For the production of effective blends, it is crucial to understand the different modes of action of the phytomolecules and the probable existing synergies. Furthermore, the production technology must be considered. For instance, microencapsulation techniques that prevent losses during feed processing are an important consideration.
Not to be discarded: Biosecurity
The recent outbreak of African Swine Fever focused our attention on something that is sometimes neglected on the farm: biosecurity rules. According to the report “Good Practices For Biosecurity In The Pig Sector” (2010), the three main elements of biosecurity are:
segregation
cleaning
disinfection
In general terms, the following steps must be adopted with the clear goal of reducing the challenges that the pigs are facing.
Farms must be located far from other farms (regardless of the species) and ideally must be protected with natural (forest/woods) or physical barriers.
Only one entrance must be used to go into the farm (for both vehicles and people) and a disinfection procedure must be in place, either by an automatized system or by manual application of disinfectants. Equipment disinfection systems must also be in place.
Workers and any other person that enters the facility should adhere to strict biosecurity measures 24/7. The farms must have a visitors’ book including relevant data on previous visits to farms (regardless of the species).
Trucks and visitors should not have been in contact with other pigs recently (at least 48 hours previous to the visit).
Only farm workers are allowed to go into the barns unless special approval is given (followed by strict biosecurity measurements prior to the visit).
The use of clothing and footwear that are worn only in the pig unit (and certainly not during visits to other pig farms) is recommended.
No materials (e.g. tools) can be moved from one barn to another barn. People that enter a barn should change footwear and wash their hands with soap for at least 10 seconds.
These simple actions can make a big difference to the performance of the pigs, and as a consequence to the profitability of a swine farm.
Take-home messages
Different formulations and reassessed nutritional level recommendations have been on the radar for a couple of years. It is high time to consider using efficient additives to support the pigs’ gut health. Phytomolecules appear as one of the most prominent tools to reduce pathogenic stress in pig production. Either via feed or water, phytomolecules are proven to reduce bacterial contamination and therefore reduce the need for antibiotic interventions. Furthermore, a more careful look at our daily activities in the farm is crucial. Paying attention to biosecurity and to feed safety should be standard tools to improve performance and the success of pig production operations.
References are available upon request.
*The article was initially published in the PROCEEDINGS OF THE PFQC 2019
How phytomolecules support antibiotic reduction in pig production
by Merideth Parke, Regional Technical Manager, EW Nutrition
To contain and reverse antimicrobial resistance, consumers and government regulators expect changes in pork production with the clear goal to reduce antibiotic use. For healthy, profitable pig production with simultaneous antibiotic reduction, a holistic strategy is required: refocusing human attitudes and habits, optimal pig health and welfare, and applying potential antibiotic alternatives.
Corn is often contaminated with Aspergillus fungi that can produce poisonous mycotoxins
Pig producers need to manage pathogenic pressure while reducing antibiotics
Intensive pig production has stress points associated with essential husbandry procedures such as weaning, health interventions, and dietary modifications. Stress is widely accepted to have a negative impact on immune system effectiveness, enhancing opportunities for pathogenic bacteria to invade at a local or systemic level. The gastrointestinal and respiratory systems are highly susceptible to developing disease as a result of these combined factors. Interventions such as antibiotics are commonly implemented to reduce the impact of pathogens and manage pig health. Processes that minimize the number of pathogens in the environment are the foundation for a successful antibiotic reduction plan. The challenge is to smartly combine strategies to keep the gastrointestinal and respiratory tract intact and robust.
Phytomolecules, the specific active defense compounds found in plants, have been identified as capable of enhancing pig health through antimicrobial (Cimanga et al., 2002, Franz et al., 2010), antioxidative (Katalinic et al., 2006, Damjanovic-Vratnica et al., 2007, Lee et al., 2011), digestion-stimulating and immune-supportive functions. As many thousands of phytomolecules exist, laboratory research has focused on identifying those with the capability of microbial management, facilitating the end goal of reducing the reliance on antibiotics for pig health and welfare and the production of safe pork (Zhai et al., 2018).
Which roles can phytomolecules play in reducing antibiotics?
The gastrointestinal tract benefits from applying phytomolecules such as capsaicin, carvacrol, and cinnamaldehyde, as they:
support a balanced and stable biome,
prevent dysbiosis, maintain tight junction integrity (Liu et al., 2018),
Pigs most susceptible and in need of phytomolecule gastrointestinal supportive actions are piglets at weaning and pigs of all ages undergoing stress, pathogen challenges, and/or dietary changes.
Porcine respiratory disease is a complex multifactorial disorder. It frequently requires antibiotics to manage infection pressure and clinical disease to maintain pig health, welfare, and production performance. Causal pathogens may be transmitted by direct contact between pigs in saliva (Murase et al., 2018) or bioaerosols (LeBel et al., 2019), via the nasal or oral cavities (inhalation directly into the airways and lungs), or via an unhealthy gut. Phytomolecules such as carvacrol and cinnamaldehyde have antimicrobial properties. Hence, they may help contain respiratory pathogens in their natural habitat (the upper respiratory tract) or during transit through the oronasal cavity and gastrointestinal tract (Swildens et al., 2004, Lee et al., 2001).
In addition to supporting the gastrointestinal and respiratory systems, phytomolecules such as menthol and 1,8-cineole have been shown to enhance the physical and adaptive immune systems in multiple species (Brown et al., 2017, Barbour et al., 2013). When applied via drinking water, adherence to the oronasal mucosa facilitates the inhalation of the active phytomolecule compounds into the respiratory tract. There, they act as mucolytics, muscle relaxants, and enhancers of the mucociliary clearance mechanism (Başer and Buchbauer, 2020). Phytomolecules have also been documented to positively influence the adaptive immune system, promoting both humoral and cell-mediated immune responses (Awaad et al., 2010, Gopi et al., 2014, Serafino et al., 2008).
How phytomolecules feature in the holistic approach to antibiotic reduction
Antibiotic reduction programs positively enact social responsibility by reducing the risk to farmworkers of exposure to antimicrobial-resistant bacteria. They also help maintain or increase efficiency in safe pork production – pork with minimal risk of antibiotic residues.
Implementation of a successful health program with reduced antibiotic use will require:
application of strict internal and external biosecurity processes;
evaluation and monitoring of AMR bacteria;
partnerships with specialist nutritionists to target a lifetime healthy gut biome; and
phytomolecule-assisted health management (Figure 1).
Figure 1: The role of phytomolecules within EW Nutrition’s holistic Antibiotic Reduction program
A combination of in vitro and in vivo studies provides evidence that specific phytomolecules can support both enteric and respiratory systems through biome stabilisation and pathogen management (Bajabai et al., 2020). Antimicrobial activity of thymol, carvacrol, and cinnamaldehyde has been reported against respiratory pathogens including S. suis, A. pleuropneumoniae, and H. parasuis (LeBel et al., 2019); multi-drug resistant and ESBL bacteria (Bozin et al., 2006); enteric pathogens including E. coli, Salmonella enteritidis, Salmonella cholerasuis, and Salmonella typhimurium (Penalver et al., 2005); Clostridium spp., E. coli spp., Brachyspira hyodysenteriae (Vande Maelle et al., 2015); and Lawsonia intracellularis (Draskovic et al., 2018). These results have shown phytomolecules to be effective antimicrobial alternatives for incorporation into holistic pig health programs.
Additionally, the inclusion of phytomolecules into pig production systems also enhances production performance by reducing the negative impact of stress on the pig and increasing the positive effects on gut health and nutrient utilization (Franz et al., 2010). Phytomolecules that directly impact digestive actions include capsaicin, which optimizes the production of digestive enzymes and increases serotonin for gut contraction maintenance and improved digesta mixing (Zhai et al., 2018). Cineol’s antioxidative activities provide support during times of stress (Cimanga et al., 2002).
Phytomolecules are key to reducing antibiotics in pig production
The pig industry searches for alternatives to therapeutic, prophylactic, and growth-promoting antibiotic applications to keep available antibiotics effective for longer – and to address the social responsibility of mitigating AMR. This search for ways to produce safe pork has made it clear that only a combination of management and antibiotic alternatives can achieve these aligned goals.
Biosecurity, hygiene, stress reduction, and husbandry and nutritional advances form the foundation for the strategic application of specific phytomolecules (Zeng et al. 2016). Supporting pig production and health, this complete holistic solution (EIP-AGRI) moves the pig industry into a future where antibiotic reduction or removal, with equivalent or increased production of safe pork, becomes a reality.
References
Awaard M, Abdel-Alim G, Sayed K, Kawkab, Ahmed1 A, Nada A , Metwalli A, Alkhalaf A. “Immunostimulant effects of essential oils of peppermint and eucalyptus in chickens”. Pakistan Veterinary Journal (2010). 2:61-66. http://www.pvj.com.pk/
Bajagai YS, Alsemgeest J, Moore RJ, Van TTH, Stanley D. “Phytogenic products, used as alternatives to antibiotic growth promoters, modify the intestinal microbiota derived from a range of production systems: an in vitro model”. Applied Microbiology and Biotechnology (2020). 104:10631-10640. https://doi.org/10.1007/s00253-020-10998-x
Barbour EK, Shaib H, Azhar E, Kumosani T, Iyer A, Harakey S, Damanhouri G, Chaudary A, Bragg RR. “Modulation by essential oil of vaccine response and production improvement in chicken challenged with velogenic Newcastle disease virus”. Journal of Applied Microbiology (2013). 115, 1278-1286. https://doi:10.1111/jam.12334
Biljana Damjanovic-Vratnica, Tatjana Dakov, Danijela Sukovic, Jovanka Damjanovic. “Antimicrobial effect of essential oil isolated from Eucalyptus globulus Labill” (2011). Czech Journal of Food Science 27(3):277-284. https://www.agriculturejournals.cz/publicFiles/39925.pdf
Bozin B, Mimica-Dukic N, Smin N, Anackov G. “Characterization of the volatile composition of essential oils of some Lamiaceae spices and the antimicrobial and antioxidant activities of the entire oils” Journal of Agriculture and Food Chemicals (2006). 54:1822-1828https://pubs.acs.org/doi/10.1021/jf051922u
Brown SK, Garver WS, Orlando RA. “1,8-cineole: An Underappreciated Anti-inflammatory Therpeutic” Journal of Biomolecular Research &Therapeutics (2017). 6:1 1-6 https://doi: 10.4172/2167-7956.1000154
Cimanga K., Kambu K., Tona L., Apers S., De Bruyne T., Hermans N., Totte J., Pieters L., Vlietinck A.J. “Correlation between chemical composition and antibacterial activity of essential oils of some aromatic medicinal plants growing in the Democratic Republic of Congo”. Journal of Ethnopharmacology (2002) 79: 213–220. https://doi.org/10.1016/s0378-8741(01)00384-1
Draskovic V, Bosnjak-Neumuller J, Vasiljevic M, Petrujkic B, Aleksic N, Kukolj V, Stanimirovic Z. “Influence of phytogenic feed additive on Lawsonia intracellularis infection in pigs” Preventative Veterinary Medicine (2018). 151: 46-51 https://doi.org/10.1016/j.prevetmed.2018.01.002
Franz C., Baser KHC, Windisch W. “Essential oils and aromatic plants in animal feeding-a European perspective. A review Flavour”. Flavour and Fragrance Journal (2010) 25:327-40. https://doi.org/10.1002/ffj.1967
Gopi M, Karthik K, Manjunathachar H, Tamilmahan P, Kesavan M, Dashprakash M, Balaraju B, Purushothaman M. “Essential oils as a feed additive in poultry nutrition”. Advances in Animal and Veterinary Sciences (2014) 1:17. https://doi.10.14737/journal.aavs/2014.2.1.1.7
Başer, Kemal Hüsnü Can, and Gerhard Buchbauer. Handbook of Essential Oils Science, Technology, and Applications. Boca Raton: CRC Press, 2020.
Hengziao Zhai, Hong Liu, Shikui Wang, Jinlong Wu, Anna-Maria Kluenter. “Potential of essential oils for poultry and pigs.” Animal Nutrition 4 (2018): 179-186. https://doi.org/10.1016/j.aninu.2018.01.005
Katalinic V., Milos M., Kulisic T., Jukic M. “Screening of 70 medicinal plant extracts for antioxidant capacity and total phenols”. Food Chemistry (2006) 94(4):550-557. https://doi.org/10.1016/j.foodchem.2004.12.004
LeBel G., Vaillancourt K., Bercier P., Grenier D. “Antibacterial activity against porcine respiratory bacterial pathogens and in vitro biocompatibility of essential oils”. Archives of Microbiology (2019) 201:833-840; https://doi.org/10.1007/s00203-019-01655-7
Lee KG, Shibamoto T. “Antioxidant activities of volatile components isolated from Eucalyptus species”. Journal of the Science of Food and Agriculture (2001). 81:1573-1597. https://doi.org/10.1002/jsfa.980
Liu SD, Song MH, Yun W, Lee JH, Lee CH, Kwak WG Han NS, Kim HB, Cho JH. “Effects of oral administration of different dosages of carvacrol essential oils on intestinal barrier function in broilers” Journal of Animal Physiology and Animal Production (2018) https://doi.org/10.1111/jpn.12944
Murase K, Watanabe T, Arai S, Kim H, Tohya M, Ishida-Kuroki K, Vo T, Nguyen T, Nakagawa I, Osawa R, Nguyen N, Sekizaki T. “Characterization of pig saliva as the major natural habitat of Streptococcus suis by analyzing oral, fecal, vaginal, and environmental microbiota”. PLoS ONE (2019). 14(4). https://doi.org/10.1371/journal.pone.0215983
Serafino A, Vallebona PS, Adnreola F, Zonfrillo M, Mercuri L, Federici M, Rasi G, Garaci E, Pierimarchi P. “Stimulatory effect of Eucalyptus essential oil on innate cell-mediated immune response” BioMed Central (2008). 9:17 https//:doi:10.1186/1471-2172-9-17
Swildens B, Stockhofe-Zurwieden N, van der Meulen J, Wisselink HJ, Nielen M. “Intestinal translocation of Streptococcus suis type 2 EF+ in pigs”. Veterinary Microbiology (2004) 103:29-33. https://doi: 10.1016/j.vetmic.2004.06.010
Vande Maele L, Heyndrickx M, Maes D, De Pauw N, Mahu M, Verlinden M, Haesbrouck F, Martel A, Pasmans F, Boyen F. “In vitro susceptibility of Brachyspira hyodysenteriae to organic acids and essential oil components”. Journal of Veterinary Medical Science (2016). 78(2):325-328. https://doi.org/10/1292/jvms.15-0341
Zeng Z, Zhang S, Wang H, Piao X. “Essential oil and aromatic plants as feed additives in non-ruminant nutrition: a review”. Journal of Animal Science and Biotechnology (2015) 6:7. https://doi.org?10/1186/s40104-015-004-5
Respiratory Challenges: Breathing Space for Antibiotic Reduction?
Sub-therapeutic doses of antibiotic growth promoters (AGPs) were used for more than 50 years in poultry production to achieve performance targets – until growing concerns arose regarding antibiotic resistance (Kabir, 2009) and decreasing efficacy of antibiotics for medical purposes (Dibner & Richards, 2005).
Isolates of ESBL-producing E.coli from animals, farmworkers, and the environment were found to have identical multidrug resistance patterns (A. Nuangmek et al., 2018). There is also evidence that AMR strains of microorganisms spread from farm animal to animal workers and beyond. Global AMR fatalities are increasing and might reach 10 million by 2050 (Mulders et al., 2010, Trung et al., 2017, Huijbers et al., 2014).
In light of this, certain AGPs have already been banned, and there is a strong possibility of future restrictions on their use worldwide. Bans are effective: the MARAN report 2018 shows that lower antibiotics usage following the EU ban on AGPs has reduced resistant E.coli in broilers. Another positive consideration is the market opportunities that exist for antibiotic residue-free food.
However, the key element that poultry producers need to get right for antibiotic reduction to be successful is respiratory health management. This article looks at why respiratory health is a particular challenge – and how phytogenic solutions can help.
A closer look at the chickens’ respiratory system
The respiratory tract is equipped with a functional mucociliary apparatus consisting of a protective mucous layer, airway surface liquid layer, and cilia on the surface of the ciliated cells. This apparatus produces mucus, which traps the inhaled particles and pathogens and propels them out of the airways. This mechanism, called the mucociliary clearance, is the primary innate defense mechanism of the respiratory system.
High stocking density combined with stressful environmental factors can negatively influence birds’ immune systems (Heckert et al., 2002; Muniz et al., 2006), making them more susceptible to respiratory disease. When a bird suffers from respiratory disease, which is nowadays usually complicated by a co-infection or secondary bacterial infection, there is an excess production of mucus that results in ciliostasis and, therefore, in an impaired mucociliary clearance. The excess mucus in the tract obstructs the airways by forming plagues and plugs, resulting in dyspnea (hypoxia) and allowing the invasive bacteria to adhere and colonize the respiratory system.
The build-up of mucus in the respiratory tract severely reduces oxygen intake, causing breathlessness, reduced feed intake, and a drop in the birds’ energy levels, which negatively impacts weight gain and egg production. Respiratory problems can result from infection with bacteria, viruses, and fungi, or exposure to allergens. The resultant irritation and inflammation of the respiratory tract leads to sneezing, wheezing, and coughing – and, therefore, the infection rapidly spreads within the flock.
Relatively high stocking density is the norm in poultry production
Low or no antibiotics: how to manage respiratory disease?
Unsurprisingly, respiratory diseases in poultry are a major cause of mortality and economic loss in the poultry industry. For Complicated Chronic Respiratory Disease (CCRD), for instance, although the clinical manifestations are usually slow to develop, Mycoplasma gallisepticum (MG), in combination with E. coli, can cause severe airsacculitis. Beside feed and egg production reduction, these problems are of high economic significance since respiratory tract lesions can cause high morbidity, high mortality, and significant carcass condemnation and downgrading.
Producers need to pre-empt the spread of respiratory pathogens, react quickly to alleviate respiratory distress and maintain the mucociliary apparatus’ functionality. Traditionally, treatment options are based on antiviral, anti-inflammatory, and antibiotic drugs. Can the poultry industry limit losses from respiratory infections without excessive recourse to antibiotics?
Indeed, a sudden reduction in antibiotic usage comes with a risk of impaired performance, increased mortality, and impaired animal health and welfare. The impact has been quantified as a 5% loss in broiler meat production per sq. meter (Gaucher et al., 2015). Effective antibiotics reduction requires a combination of innovative products and suitable consultancy services to manage poultry gut health, nutrition, flock management, biosecurity, and, particularly, respiratory health.
Non-antibiotic alternatives to control diseases and promote broiler growth, such as organic acids (Vieira et al., 2008), probiotics (Mountzouris et al., 2010), prebiotics (Patterson & Burkholder, 2003), and essential oils (Basmacioğlu Malayoğlu et al., 2010) have been the subject of much research in recent years.
Phytogenic solutions: proven efficacy
Essential oils, which are extracted from plant parts, such as flowers, buds, seeds, leaves, twigs, bark, wood, fruits, and roots, have a particularly well-established track record of medicinal applications. Efforts have centered on phytomolecules, the biologically active secondary metabolites that account for the properties of essential oils (Hernández et al., 2004; Jafari et al., 2011).
Studying these properties is challenging: essential oils are very complex natural mixtures of compounds whose chemical compositions and concentrations are variable. For example, the concentrations of the two predominant phytogenic components of thyme essential oils, thymol and carvacrol, have been reported to range from as low as 3% to 60% of the whole essential oil (Lawrence and Reynolds, 1984).
Another well-researched example is eucalyptus oil. The essential oils of eucalyptus species show antibacterial, anti-inflammatory, diaphoretic, antiseptic, analgesic effects (Cimanga et al., 2002) and antioxidant properties (Lee and Shibamoto, 2001; Damjanović Vratnica et al., 2011). The oils are mainly composed of terpenes and terpene derivatives in addition to some other non-terpene components (Edris, 2007). The principal constituent found in eucalyptus is 1,8-cineole (eucalyptol); however, other chemotypes such as α-phellandrene, ρ-cymene, γ-terpinene, ethanone, and spathulenol, among others, have been documented (Akin et al., 2010).
Close-up of eucalyptus leaf oil glands and
the molecular structure of eucalyptol C10H18O (red = oxygen; dark grey = carbon; light grey = hydrogen)
Antimicrobial activity
In modern intensive broiler production, bacterial diseases such as salmonellosis, colibacillosis, mycoplasmosis, or clostridia pose serious problems for the respiratory system and other areas. Analyses of the antibacterial properties of essential oils have been carried out by multiple research units (Ouwehand et al., 2010; Pilau et al., 2011; Solorzano- Santos and Miranda-Novales, 2012; Mahboubi et al., 2013; Nazzaro et al., 2013; Petrova et al., 2013).
Phenols, alcohols, ketones, and aldehydes are clearly associated with antibacterial activity; the exact mechanisms of action, however, are not yet fully understood (Nazzaro et al., 2013). Essential oils’ antimicrobial activity is not attributable to a unique mechanism but instead results from a cascade of reactions involving the entire bacterial cell (Nazzaro et al., 2013). However, it is accepted that antimicrobial activity depends on the lipophilic character of the components.
The components permeate the cell membranes and mitochondria of the microorganisms and inhibit, among others, the membrane-bound electron flow and thus the energy metabolism. This leads to a collapse of the proton pump and draining of the ATP (adenosine triphosphate) pool. High concentrations may also lead to lysis of the cell membranes and denaturation of cytoplasmic proteins (Nazzaro et al., 2013; Gopi et al., 2014).
According to current knowledge, lavender, thyme, and eucalyptus oil, as well as the phytomolecules they contain, show enhanced effects when combined with other essential oils or synthetic antibiotics (Sadlon and Lamson, 2010; Bassole and Juliani, 2012; Sienkiewicz, 2012; de Rapper et al., 2013; Zengin and Baysal, 2014).
Minimum inhibitory concentration (MIC) of some essential oil components against microorganisms in vitro
Immune system boost I: improved production of antibodies
Some essential oils were found to influence the avian immune system positively, since they promote the production of immunoglobulins, enhance the lymphocytic activity, and boost interferon-γ release (Awaad et al., 2010; Faramarzi et al., 2013; Gopi et al., 2014; Krishan and Narang, 2014). Placha et al. (2014) showed that the addition of 0.5g of thyme oil per kg of feed significantly increased IgA levels.
Awaad et al. (2010) experimented on birds vaccinated with the inactivated H5N2 avian influenza vaccine. The experiment revealed that adding eucalyptus and peppermint essential oils to the water at a rate of 0.25 ml per liter resulted in an enhanced cell-mediated and humoral immune response.
Saleh et al. (2014), who applied thyme and ginger oils in quantities of 100mg and 200mg per kg of feed, respectively, observed an improvement in chickens’ immunological blood profile through increased antibody production. Rehman et al. (2013) stated that the use of herbal products containing eucalyptus oil and menthol in broilers showed consistently higher antibody titers against NDV (Newcastle disease virus), compared to untreated broilers.
Immune system boost II: better vaccine responses and anti-inflammatory effects
Essential oils are also used as immunomodulators during periods when birds are exposed to stress, acting protectively and regeneratively. Importantly, the oils alleviate the stress caused by vaccination (Barbour et al., 2011; Faramarzi et al., 2013; Gopi et al., 2014). The study by Kongkathip et al. (2010) confirmed the antiviral activity of turmeric essential oil.
In recent years studies have been carried out on the use of essential oils in conjunction with vaccination programs, including those against infectious bronchitis (IB), Newcastle disease, and Gumboro disease. The results of the experiments show that essential oils promote the production of antibodies, thus enhancing the efficacy of vaccination (Awaad et al., 2010; Barbour et al., 2010; Barbour et al., 2011; Faramarzi et al., 2013).
Essential oils contain compounds that are known to possess strong anti-inflammatory properties, mainly terpenoids, and flavonoids, which suppress the metabolism of inflammatory prostaglandins (Krishan and Narang, 2014). Also, other compounds found in essential oils have anti-inflammatory, pain-relieving, or edema-reducing properties, for example, linalool from lavender oil, or 1,8-cineole, the main component of eucalyptus oil (Peana et al., 2003).
Immune system boost III: antioxidant effects and radical scavenging
An imbalance in the rate of production of free radicals or removal by the antioxidant defense mechanisms leads to a phenomenon referred to as oxidative stress. A mixture of Oregano (carvacrol, cinnamaldehyde, and capsicum oleoresin) was found to beneficially affect the intestinal microflora, absorption, digestion, weight gain and also to have an antioxidant effect on chickens (Bassett, 2000).
Zeng et al. (2015) indicated the positive effect of essential oils on the production of digestive secretions and nutrient absorption. They reduce pathogenic stress in the gut, exert antioxidant properties, and reinforce the animal’s immune status.
Inside the cell, essential oils can serve as powerful scavenger preventing mutations and oxidation (Bakkali et al., 2008). Studies have demonstrated the concentration-dependent free radical scavenging ability of oils from eucalyptus species (Kaur et al., 2010; Marzoug et al., 2011; Olayinka et al., 2012). Some authors attribute the strong antioxidant capacity of essential oils to their phenolic constituents and synergistic effect between tannins, rutin, thymol, and carvacrol, and probably 1, 8-cineole. Moderate DPPH radical scavenging activity reported by Edris(2007), El-Moein et al. (2012), and Kaur et al. (2011).
Vázquez et al. (2012) have demonstrated the potential of the phenolic compounds in eucalyptus bark as a source of antioxidant compounds. The study showed that eucalyptus had ferric reducing antioxidant power in the ranges 0.91 to 2.58 g gallic acid equivalent (GAE) per 100 g oven-dried bark and 4.70 to 11.96 mmol ascorbic acid equivalent (AAE) per 100 g oven-dried bark, respectively (see also Shahwar et al., 2012). Moreover, Eyles et al. (2004) were able to show superoxide dismutase (SOD)-like activity for different compounds and fractions isolated from wood extracts.
Last but not least: positive effects on the respiratory system
In poultry production houses, especially in summer, high temperatures and low humidity increase the amount of air dust. Under such conditions, respiratory tract disorders in broiler chickens, including the deposition of particulates, become more frequent and more severe.
Clinical signs of respiratory disease in chickens include coughing, sneezing, and rales
Thyme oil, thanks to the phytomolecules thymol and carvacrol, supports the treatment of respiratory disorders. These substances smooth tightened muscles and stimulate the respiratory system. An additional advantage lies in their expectorant and spasmolytic properties (Edris, 2007).
These properties are also seen in essential oils such as eucalyptus and peppermint, which contain eucalyptol and menthol. They thin out the mucus and facilitate its removal from the airways. As a result, the airways are cleared and breathing during inflammation becomes easier (Durmic and Blache, 2012).
Another positive effect of the terpenoid compounds used in commercial preparations for poultry is that they disinfect the bronchi, preventing respiratory infections (Awaad et al., 2010; Barbour et al., 2011; Mahboubi et al., 2013). Barbour and Danker (2005) reported that the essential oils of eucalyptus and peppermint improved the homogeneity of immune responses and performance in MG/H9N2-infected broilers.
Grippozon: the phytogenic solution for respiratory health
Grippozon is a liquid composition with a high content of essential oils, which are combined to systematically prevent and ease respiratory diseases. The formulation is derived from the research on essential oils’ effectiveness against respiratory pathogens that are common in animal farming. Grippozon exhibits a synergistic action of all its components to optimally support animal health. It contains a high concentration of active components; both their quantity and quality are guaranteed to deliver results.
Application of Grippozon
Grippozon application can be flexibly adapted to most common housing systems. It is fully water-soluble for use in the drinking line and it is also possible to nebulize a diluted solution in air.
The dose recommendation in drinking water usually amounts to 100ml to 200ml per 1000 liters of drinking water (Grippozon administration has not been reported to affect water consumption). The active substances in Grippozon adhere to mouth mucosa and become volatile in the breathing air later on. Therefore Grippozon can enter the respiratory system indirectly as well. The volatile compounds also spread into the whole barn air and, thus, indirectly via breathing into the respiratory system (and farmers notice the smell of essential oils when Grippozon is applied through in the waterline)
Grippozon can also be used as a spray at a rate of 200ml/10 liters of water for 2000 birds, twice daily on 2-3 days a week. This produces a very effective nebulization effect and offers faster respiratory relief to birds.
Grippozon is an impactful tool for managing respiratory problems. Thanks to its effective mucolytic and relaxant activity, Grippozon gives symptomatic relief to the birds during high-stress periods of respiratory diseases. Mucus in the trachea works as media for the proliferation of bacteria and viruses, so by thinning the mucus, Grippozon slows down the proliferation of bacteria and the spread of disease. Grippozon helps in improving air quality and air intake. It can also be used to stimulate the immune response during vaccination.
Do we have the tools to reduce antibiotics in swine production?
The global swine industry is going through unprecedented challenges. On the one hand, the threat of the African Swine Fever virus is global, despite the fact it hasn’t arrived in all markets. The virus is today alive among the wild boars in the Polish and Belgian forests. Every day it keeps gaining a few more meters to the border, threatening the German swine industry, one of the largest in the European Union.
If this happens, we might be seeing important changes to the pork supply chain on the meat market worldwide – in Europe in addition to current issues in the USA meat plants. The profitability of swine businesses depends in many ways on the export capacity of large corporations based in Germany, Spain, Denmark, etc.
On the other hand, the presence of COVID-19 in most countries is changing human behavior, meat consumption at home, and the way we look at the future. Perhaps a virus overload via the news, some “fake news” conveying wrong messages on what’s coming, and suddenly we feel the future will never be the same.
The future of the swine industry
At least for the swine industry, the future will indeed never be exactly the same. We will be facing different challenges. Some of these will be structural, such as the issue of decreased manpower and how to substitute manpower by machines, through the implementation of Precision Livestock Farming, for instance.
We are also facing important health challenges to our animals: not just ASF, but also new and more aggressive PRRS strains, among other pathogens. Our sows´ production capacity is increasing annually, yet in some cases 25% of the new-born piglets are lost from birth to market. Increasingly, we may start to see increased levels of mortality not only in the nursery but in fattening pigs and sows as well.
It is becoming clearer all the time: the future of the global swine industry lies in producing more pigs with reduced antibiotics. To stay the course, we need to take further action and implement corrective measures.
Why we should remove antibiotics in pig production
Pressure from stakeholders and regulators
There is, and there will be, increasing pressure from many stakeholders worldwide to work toward pig production with reduced or no antibiotics. Meat suppliers, slaughterhouses and processors, governments at different levels, and, of course, the European Union – all are demanding reductions in the level of antibiotics in livestock production.
There is also an increasing awareness at the global societal level regarding antimicrobial resistance related to antibiotic usage in farming production. Consumer pressure will grow exponentially as the terrible COVID-19 experience will be “digested” by the global population.
Pressure to accede to the pork market
There is yet another important reason to start working in that direction: the global swine meat market. Today, China’s pork meat shortage is opening the market. Now any producer could potentially sell meat, either to China or to any other country. We are starting to see moves from companies in the USA or Brazil banning the use of Ractopamine in their operations because they want to get access to the ractopamine-free market (Europe & Asia, over 70% of the global population).
According to M. Pierdon (AASV 2020 Proceedings), there will be two types of markets: the “Niche ABFree” and the “Commodity ABFree”. Companies will have to analyse what their future is on the meat market. Not all the producers may be willing to enter this new phase, but for sure many will try.
Strategies for antibiotic reduction
In Europe, the time has arrived. Zinc oxide will be banned in June 2021 and there is now more than a trend in production with less or no antibiotic use. In some cases, there is a need; in others, this is simply profitable.
Challenges to antibiotic reduction
Producing pigs completely without antibiotics is not easy, and not affordable for all. Initially we may have to give up some performance parameters in order to achieve the balance between what we want and what we can achieve in animal performance. But the time will arrive when these two objectives will converge; there is no alternative.
To that end, we will have to include in our pig production strategy all the available tools and technologies: genetic selection, immunization against some key pathogens, environmental control (mandatory but often forgotten), early detection of diseases, etc.
In this new era we are entering, nutrition and feed additives will play a key role. It will be crucial to find solutions targeting the microbiome’s stabilization and diversification, creating and maintaining healthy farms and achieving all the performance parameters.
Do we have the tools for antibiotic reduction?
Even today there are companies able to produce completely antibiotic-free pigs – proof that yes, the tools are already in place.
Nevertheless, for most producers, the answer to – Can we produce without antibiotics? is most likely “probably not”. This will require a holistic approach, given the specific case of piglets.
The microbiome of the piglet is strongly influenced by birth and the subsequent weeks. What, then, are the elements that will be part of this new game that comprises a new approach?
The colostrum intake & the management of the piglets
Antibiotic usage and its influence on the gut
The piglets’ microbiome and its evolution during the periweaning period
The weaning process, appetite, and water intake
Zinc oxide removal and its influence on the microbiome
The immune system and the relationship with the GIT status
Inflammation and its modulation at the gut level
The health status and the effect on the concomitant infections: which ones are key and which ones are secondary pathogens
The all-important biosecurity, management, and hygiene
To summarize: there is no one tool, but rather a holistic approach to face this new challenge that the swine industry is facing nowadays. The answer is not a silver bullet, but a journey that we all must undertake.
A complex battlefield: mycotoxins in the gastrointestinal tract
Most grains used as feed raw materials are susceptible to mycotoxin contamination. These toxic secondary metabolites are produced by fungi before or after harvest and cause severe economic losses all along agricultural value chains. For livestock, negative consequences include acute effects such as impaired liver and kidney function, vomiting, or anorexia, as well as chronic effects such as immunosuppression, growth retardation, and reproductive problems. Mycotoxin management is, therefore, of the utmost priority for animal producers worldwide.
But how is it that mycotoxins cause such damage in the first place? This article delves into the complex processes that take place when mycotoxins come into contact with the gastrointestinal tract (GIT). The intestinal epithelium is the first tissue to be exposed to mycotoxins, and often at higher concentrations than other tissues. A deeper understanding of how mycotoxins affect the GIT allows us to appreciate the cascading effects on animal health and performance, why such damage already occurs at contamination levels well below official safety thresholds – and what we can do about it.
The intestinal epithelium: the busy triage site for nutrients and harmful substances
When mycotoxins are ingested, they encounter the GIT’s intestinal epithelium (Figure 1). This single layer of cells lining the intestinal lumen serves two conflicting functions: firstly, it must be permeable enough to allow the absorption of nutrients. On the other hand, it constitutes the primary physiological barrier against harmful agents such as viruses, microorganisms, and toxins.
Within the intestinal epithelium, several types of highly specialized cells are involved in epithelial regeneration, nutrient absorption, innate defense, transport of immunoglobulins, and immune surveillance. The selective barrier function is maintained due to the formation of complex networks of proteins that link adjacent cells and seal the intercellular space. Besides, the intestinal epithelium is covered with mucus produced by goblet cells, which isolates its surface, preventing the adhesion of pathogens to the enterocytes (intestinal absorptive cells).
Due to its dual involvement in digestive and immune processes, the intestinal epithelium plays a pivotal role in the animal’s overall health. Importantly, the epithelium is directly exposed to the entire load of ingested mycotoxins. Hence their effects can be problematic even at low concentrations.
Figure 1: The intestinal epithelium
Problematic effects of mycotoxins on the intestinal epithelium
Most mycotoxins are absorbed in the proximal part of the gastrointestinal tract (Table 1). This absorption can be high, as in the case of aflatoxins (~90%), but also very limited, as in the case of fumonisins (<1%); moreover, it depends on the species. Importantly, a significant portion of unabsorbed toxins remains within the lumen of the gastrointestinal tract.
Some of the mycotoxins that enter the intestinal lumen can be bio-transformed into less toxic compounds by the action of certain bacteria. This action, however, predominantly happens in the large intestine – therefore, no detoxification takes place before absorption in the upper parts of the GIT. Part of the absorbed mycotoxins can also re-enter the intestine, reaching the cells from the basolateral side via the bloodstream. Furthermore, they re-enter through enterohepatic circulation (the circulation of substances between the liver and small intestine). Both actions increase the gastrointestinal tract’s overall exposure to the toxins.
Table 1: Rate and absorption sites of different mycotoxins
The damaging impact of mycotoxins on the intestinal epithelium initially occurs through:
A decrease in protein synthesis, which reduces barrier and immune function (Van de Walle et al., 2010)
Increased oxidative stress at the cellular level, which leads to lipid peroxidation, affecting cell membranes (Da Silva et al., 2018)
Changes in gene expression and the production of chemical messengers (cytokines), with effects on the immune system and cellular growth and differentiation (Ghareeb et al., 2015)
The induction of programmed cell death (apoptosis), affecting the reposition of immune and absorptive cells (Obremski & Poniatowska-Broniek, 2015)
Importantly, studies based on realistic mycotoxin challenges (e.g., Burel et al., 2013) show that the mycotoxin levels necessary to trigger these processes are lower than the levels reported as safe by EFSA, the Food Safety Agency of the European Union. The ultimate consequences range from diminished nutrient absorption to inflammatory responses and pathogenic disorders in the animal (Figure 2).
Figure 2: Mycotoxins’ impact on the GIT and consequences for monogastric animals
1. Alteration of the intestinal barrier‘s morphology and functionality
The mycotoxins DON, fumonisin, and T2 induce a reduction in the rate of epithelial cell proliferation and differentiation. This causes a decrease in the height and the surface of the intestinal villi, which in turn leads to a reduction in nutrient absorption. Additionally, some nutrient transporters are inhibited by the action of mycotoxins such as DON and T2, for example, negatively affecting the transport of glucose.
Several studies indicate that mycotoxins such as aflatoxin B1, DON, fumonisin B1, ochratoxin A, and T2, can increase the permeability of the intestinal epithelium of poultry and swine (e.g. Pinton & Oswald, 2014). This is mostly a consequence of the inhibition of protein synthesis. As a result, there is an increase in the passage of antigens into the bloodstream (e.g., bacteria, viruses, and toxins). This increases the animal’s susceptibility to infectious enteric diseases. Moreover, the damage that mycotoxins cause to the intestinal barrier entails that they are also being absorbed at a higher rate.
2. Impaired immune function in the intestine
The intestine is a very active immune site, where several immuno-regulatory mechanisms simultaneously defend the body from harmful agents. Immune cells are affected by mycotoxins through the initiation of apoptosis, the inhibition or stimulation of cytokines, and the induction of oxidative stress. Studies demonstrate that aflatoxin, DON, fumonisin, T2, and zearalenone interact with the intestinal immune system in such a manner that the animal’s susceptibility to viral and bacterial infections increases (e.g., Burel et al., 2013). Moreover, by increasing their fecal elimination, the horizontal transmission of pathogens is extended.
For poultry production, one of the most severe enteric problems of bacterial origin is necrotic enteritis, which is caused by Clostridium perfringens toxins. Any agent capable of disrupting the gastrointestinal epithelium – e.g. mycotoxins such as DON, T2, and ochratoxin – promotes the development of necrotic enteritis. The inhibition of the intestinal immune system caused by mycotoxins such as aflatoxin, DON, and T2 also collaborates with the development of this disease.
3. Alteration of the intestinal microflora
The gastrointestinal tract is home to a diverse community of bacteria, fungi, protozoa, and viruses, which lines the walls of the distal part of the intestine. This microbiota prevents the growth of pathogenic bacteria through competitive exclusion and the secretion of natural antimicrobial compounds, volatile fatty acids, and organic acids.
Recent studies on the effect of various mycotoxins on the intestinal microbiota show that DON and other trichothecenes favor the colonization of coliform bacteria in pigs. DON and ochratoxin A also induce a greater invasion of Salmonella and their translocation to the bloodstream and vital organs in birds and pigs – even at non-cytotoxic concentrations. It is known that fumonisin B1 may induce changes in the balance of sphingolipids at the cellular level, including for gastrointestinal cells. This facilitates the adhesion of pathogenic bacteria, increases in their populations, and prolongs infections, as has been shown for the case of E. coli.
From the perspective of human health, the colonization of the intestine of food-producing animals by pathogenic strains of E. coli and Salmonella is of particular concern. Mycotoxin exposure may well increase the transmission of these pathogens, posing a risk for human health.
4. Interaction with bacterial toxins
When mycotoxins induce changes in the intestinal microbiota, this can lead to an increase in the endotoxin concentration in the intestinal lumen. Endotoxins or lipopolysaccharides (LPS) are fragments of Gram-negative bacteria’s cell walls. They are released during bacterial cell death, growth, and division. Hence endotoxins are always present in the intestine, even in healthy animals. Endotoxins promote the release of several cytokines that induce an enhanced immune response, causing inflammation, thus reducing feed consumption and animal performance, damage to vital organs, sepsis, and death of the animals in some cases.
The synergy between mycotoxins and endotoxins can result in an overstimulation of the immune system. The interaction between endotoxins and estrogenic agents such as zearalenone, for example, generates chronic inflammation and autoimmune disorders because immune cells have estrogen receptors, which are stimulated by the mycotoxin. The combination of DON at low concentrations and endotoxins in the intestine, on the other hand, has been shown to engender a decrease in transepithelial resistance and to alter the balance of the microbiota.
What to do? Proactive toxin risk management
To prevent the detrimental consequences of mycotoxins on animal health and performance, proactive solutions are needed that support the intestinal epithelium’s digestive and immune functionality and help maintain a balanced microbiome in the GIT. Moreover, it is crucial for any anti-mycotoxin product to feature both anti-mycotoxin and anti-bacterial toxin properties and that it supports the organs most targeted by mycotoxins, e.g., the liver. EW Nutrition’s Mastersorb Gold premix is based on the synergistic combination of natural clay minerals, yeast cell walls, and phytomolecules. Its efficacy has been extensively tested, including as a means for dealing with E. coli endotoxins.
Mastersorb Gold: anti-mycotoxin activity stabilizes performance and strengthens liver health
A field trial conducted in Germany on male Ross 308 broilers showed that for broilers receiving a diet contaminated with DON and zearalenone, adding 1kg of Mastersorb Gold per ton of feed to their diet led to significant performance enhancements. Not only did they recuperate the mycotoxin-induced weight loss (6% increase relative to the group receiving only the challenge), but they gained weight relative to the control group (which received neither the challenge nor Mastersorb Gold). Feed conversion also improved by 3% relative to the group challenged with mycotoxins.
A scientific study of crossbred female pigs showed that Mastersorb Gold significantly reduced the deleterious effects of fumonisin contamination in the feed. The decrease in weight gain and the decline of feed conversion could be mitigated by 6.7% and 13 FCR points, respectively (Figure 3). Also, the sphinganine/sphingosine (Sa/So) ratio, a biomarker for fumonisin presence in the blood serum, could be decreased by 22.5%.
Figure 3: Mastersorb Gold boosts performance for pigs fed a fumonisin-contaminated diet
Another study of crossbred female piglets, carried out at a German university, investigated whether Mastersorb Gold could support performance as well as liver health under a naturally occurring challenge of ZEA (~ 370ppb) and DON (~ 5000ppb). Mastersorb Gold significantly improved weight gain and feed conversion in piglets receiving the mycotoxin-contaminated diet: daily body weight gain was 75g higher than that of a group receiving only the challenge, and the FCR improved by 24% (1.7 vs. 2.25 for the group without Mastersorb Gold). Moreover, Mastersorb Gold significantly improved the liver weight (total and relative) and the piglets’ AST levels (aspartate aminotransferase, an enzyme indicating liver damage). A tendency to improve spleen weight and GGT levels (gamma-glutamyl transferase, another enzyme indicative of liver issues) was also evident, all of which indicate that Mastersorb Gold effectively counteracts the harmful impact of mycotoxin contamination on liver functionality.
In-vitro studies demonstrate Mastersorb Gold’s effectiveness against mycotoxins as well as bacterial toxins
Animal feed is often contaminated with two or more mycotoxins, making it important for an anti-mycotoxin agent to be effective against a wide range of different mycotoxins. Besides, to prevent mycotoxins damaging the GIT, an effective product should ideally adsorb most mycotoxins in the first part of the animal’s intestine (under acidic conditions). In-vitro experiments at an independent research facility in Brazil showed that an application of 0.2% Mastersorb Gold binds all tested mycotoxins at rates from 95 to 97% at a pH level of 3, using realistic challenges of 1000ppb (Aflatoxin B1 and ZEA) and 2500ppb (Fumonisin B1 and DON). The binding results achieved for Fumonisin and DON, which are often considered outright “nonbinding,” under challenging close to neutral conditions (pH 6), are particularly encouraging.
Figure 4: Mastersorb Gold binding capacity against different mycotoxins (%)
Concerning its efficacy against endotoxins, an in vitro study conducted at Utrecht University, among other studies, has shown Mastersorb Gold to be a strong tool against the LPS released by E. coli. For the test, four premium mycotoxin binders were suspended in a phosphate buffer solution to concentrations of 0.25% and 1%. E. coli LPS were suspended to a final concentration in each sample of 50ng/ml. Against this particularly high challenge, Mastersorb Gold achieved a binding rate of 75% at an inclusion rate of 1%: clearly outperforming competing products, which at best showed a binding rate of 10%.
Conclusion
A healthy gastrointestinal tract is crucial to animals’ overall health: it ensures that nutrients are optimally absorbed, it provides effective protection against pathogens through its immune function, and it is key to maintaining a well-balanced microflora. Even at levels considered safe by the European Union, mycotoxins can compromise different intestinal functions such as absorption, permeability, immunity, and microbiota balance, resulting in lower productivity and susceptibility to disease.
To safeguard animal performance, it is important to continually strive for low levels of contamination in feed raw materials – and to stop the unavoidable mycotoxin loads from damaging the intestinal epithelium through the use of an effective anti-mycotoxin agent, which also supports animals against endotoxins and boosts liver function. Research shows that Mastersorb Gold is a powerful tool for proactive producers seeking stronger animal health, welfare, and productivity.
By Technical Team, EW Nutrition
Poultry health and welfare: Phytomolecules for poultry diets
The large majority of poultry specialists in Europe consider phytomolecules as one of the key elements in diets for broilers, broiler breeders, and layers when birds are raised without antibiotics. A quick glance at the market will reveal more commercial products than can possibly be imagined. There are three basic elements you should bear in mind when making your choice:
Most phytomolecules are volatile. As such, unprotected products will soon evaporate if left exposed to the open air – as it happens, for instance, with feed prepared in commercial farms. Microencapsulation is therefore essential.
There are countless phytomolecules. Consequently, finding the right mix for the task required is essential, as not all mixtures will get you the desired result. When designing a phytomolecule mix, the manufacturer must have the necessary knowledge and experience to achieve the desired result.
Phytomolecules are powerful. This is to say that you cannot just keep adding higher quantities to achieve a better result. Finding the exact inclusion rates for the right purpose is a difficult balancing exercise.
In fact, the right protection, the right mix and the right inclusion rates must be combined to ensure that the animals do not refuse the feed (worst case scenario) or just fail to benefit from the inclusion of phytomolecules.
Among the feed additives, phytomolecules (or secondary plant compounds) stand out as a class of active ingredients that may help to improve gut health and thereby reduce the use of antibiotics. Synthesized by plants as a defense mechanism against pathogens, phytomolecules promote the digestion of feed ingredients (Zhai et al. 2018), prevent loss of gut integrity during enteric challenges (Liu et al. 2018), and have antimicrobial properties that hinder the growth of potential pathogens (Chowdhury, 2018). Phytomolecules can prevent the overgrowth of opportunistic pathogens, thereby reducing the frequency of occurrence of diseases such as necrotic enteritis and dysbacteriosis and thus improve performance data such as daily weight gain and feed efficiency.
Beyond the phytomolecules’ proven effects, what works best in supporting the health and welfare of your animals is, in fact, a holistic program (such as those offered by EW Nutrition) that consists of an effective combination of innovative products and consultancy services in the fields of gut health, nutrition, AMR monitoring, and biosecurity management.

 Figure 1: Reduction of oocysts in the droppings by Pretect D
Figure 1: Reduction of oocysts in the droppings by Pretect D