Mycotoxins & Poultry Egg Quality in Southeast Asia

Tran Si Trung, PhD
EWN SEAP – Regional Technical Manager for Toxin Risk Management
1. Introduction
The global egg market is experiencing steady and robust growth, playing a vital role in food security and animal protein nutrition. According to reports from RaboResearch and the World Egg Organization, global egg production has more than doubled, rising from 46 million tonnes in 1995 to approximately 99 million tonnes in 2025. By 2035, the market is projected to expand by a further 22%, with an annual growth rate of approximately 2.0%. Asia leads with more than 64% of global output, with China and India being the largest producing countries. The global egg market value was estimated at USD 352 billion in 2025 and is expected to reach USD 585 billion by 2033, representing a CAGR of approximately 6.6%. This growth is driven by urbanization, rising incomes, demand for high-quality protein, and the widespread use of eggs in the processed food industry.
In Vietnam, the poultry industry has expanded rapidly and become one of the pillars of the agricultural sector. In 2023, poultry egg production reached approximately 19.22 billion eggs. In 2024, this figure exceeded 20 billion eggs, with chicken eggs accounting for the dominant share. Per-capita egg consumption rose from 108 eggs per year in 2017 to approximately 185–190 eggs per year in 2024, though this remains below the average of many other countries (300–350 eggs). In the near term, despite limited exports, domestic consumption is fairly stable, and the sector has the potential to achieve a value of USD 3 billion. Alongside these opportunities, the industry also faces challenges such as occasional local oversupply, price volatility, disease outbreaks, and food quality and safety issues. Among these, mycotoxins can be regarded as one of the most silent yet serious threats to egg quality and consumer health.
The principal mycotoxins include aflatoxins (AFs), ochratoxin A (OTA), zearalenone (ZEN), deoxynivalenol (DON), fumonisins (FBs), and T-2 toxin. These compounds form in feed raw materials — including maize, wheat, soybean, groundnut, and other oil seeds — under field stress conditions or during storage under inadequate conditions. In the tropical humid climates of Vietnam and much of Southeast Asia, the risk of natural contamination is particularly elevated. Mycotoxins not only reduce livestock productivity (organ damage, immune suppression, etc.) but also directly affect egg quality, nutritional value, and toxin residue levels in eggs, thereby impacting both the economic value of the product and consumer health.
2. Key Aspects of Egg Quality
Egg quality is typically assessed across multiple dimensions: external appearance (clean, intact shell, uniform shape and color), internal quality (albumen height, yolk color, Haugh unit score), nutritional value (high protein ~12–13%, lipids, vitamins A/D/E, carotenoids, choline, lutein), and food safety (freedom from microbiological contamination, antibiotic residues, mycotoxin residues, and heavy metals).
Key technical parameters include: egg weight and grade (AA, A, B); shell thickness and strength; Haugh unit (reflecting albumen freshness); yolk color (Roche scale or DSM Yolk Fan); air cell size; and absence of blood spots (meat spots). High quality ensures commercial value, shelf life, and nutritional benefit to the consumer.
Each of these quality dimensions (shell integrity, albumen height, yolk color, and residue status) is, to varying degrees, susceptible to mycotoxin insult, as the following section demonstrates.
3. Adverse Effects of Mycotoxins on Egg Quality and Value
Mycotoxins cause harm through multiple mechanisms: hepato-renal toxicity, hormonal disruption, oxidative stress, intestinal damage (reduced nutrient absorption), immune and enzyme suppression. Effects are often evident at relatively low concentrations (20–500 ppb depending on the toxin type) and are amplified when multiple mycotoxins are present simultaneously. Field surveys across Asia consistently demonstrate that co-contamination, the presence of two or more mycotoxins in a single feed ingredient or complete diet, is the norm rather than the exception, particularly in maize-based diets during wet-season harvests. Effective risk management must therefore address the full toxin spectrum rather than individual contaminants in isolation.
| Mycotoxin | Main Feed Substrates |
Primary Mechanism(s) |
Key Effects on Egg Quality |
| Aflatoxins (AFs/AFB1) |
Maize, groundnut, soybean, cottonseed |
Hepatotoxicity; oxidative stress; Ca & Zn absorption inhibition |
↓ Laying rate; ↓ shell quality; ↓ yolk carotenoids & color; residues (AFB1, AFM1) in eggs |
| Ochratoxin A (OTA) |
Wheat, barley, maize, sorghum |
Nephrotoxicity; immunosuppression |
↓ Laying rate; ↑ cracked/thin/ misshapen shells; residues in eggs |
| Zearalenone (ZEN) |
Maize, wheat, barley |
Estrogenic receptor disruption (HPG axis) |
↓ FSH/LH/progesterone; ↓ ovarian function; ↓ fertility & hatchability in breeders (roosters + hens) |
| Deoxynivalenol (DON) |
Wheat, maize, barley, oats |
Intestinal inflammation; protein synthesis inhibition |
↓ Feed intake; ↓ shell breaking strength (10–15%); ↓ Haugh unit; ↓ yolk color |
| Fumonisins (FBs/FB1) |
Maize and maize by-products |
Sphingolipid synthesis inhibition; liver damage |
↓ Laying performance; ↓ nutrient absorption; ↓ albumen & yolk quality |
| T-2 Toxin | Cereal grains (wheat, barley, oats) |
Mucosal necrosis; immunosuppression; ribotoxic effect |
↓ Feed intake (oral lesions); ↓ Haugh unit (especially combined with DON); ↓ eggshell quality |
Table 1. Overview of principal mycotoxins, their main feed substrates, primary mechanisms of action, and key effects on poultry egg quality.
3.1. Reduced Laying Performance
AFs and OTA can reduce laying rate by 5–10% at field-relevant dietary concentrations, with greater reductions reported under conditions of more severe or prolonged contamination. ZEN, acting as a potent estrogen mimic, disrupts the hypothalamic-pituitary-gonadal (HPG) axis, reducing FSH, LH, and progesterone levels, thereby impairing follicular development and ovarian function. DON and FBs cause intestinal inflammation and reduced nutrient absorption. T-2 toxin can cause ulcerative lesions of the oral mucosa or gizzard, thereby reducing feed intake or impairing gizzard motility and feed digestion.
3.2. Impact on Eggshell Quality
Mycotoxins in general can reduce eggshell thickness and strength by inhibiting calcium absorption, vitamin D3 utilization, and carbonic anhydrase activity (zinc-dependent). In particular, AFs may induce secondary zinc deficiency through liver damage. OTA has been associated with increased incidence of cracked, thin, misshapen, and urate-spotted eggs. Experimental studies have shown that DON can reduce eggshell breaking strength by 10–15% under controlled conditions.
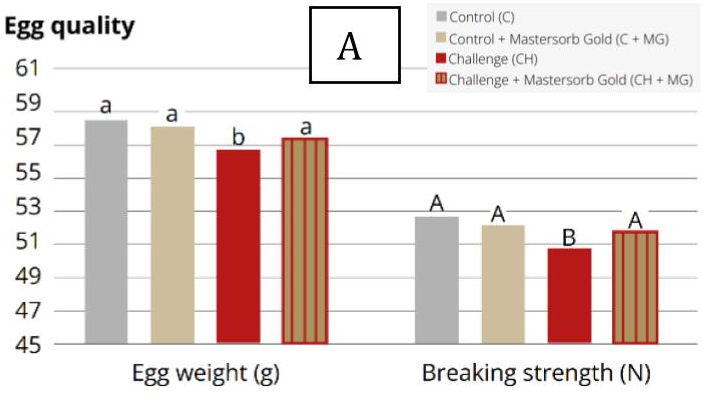
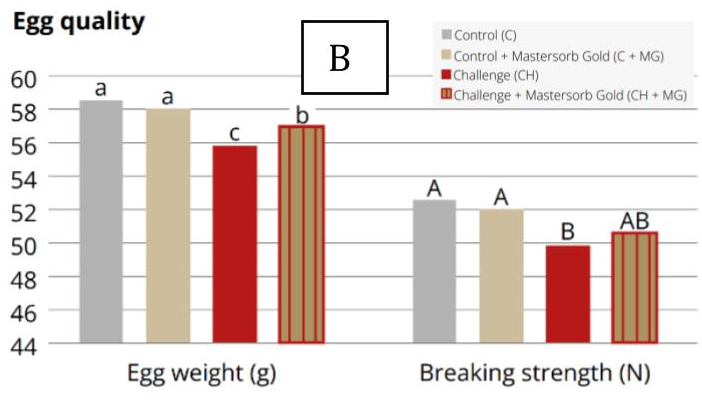
3.3. Impact on Internal Egg Quality
Research conducted at Kasetsart University (Thailand) demonstrates that DON and T-2 toxin can reduce albumen height and Haugh unit scores at relatively low dietary concentrations (Tables 2 and 3). Additionally, experimental data indicate that DON can impair yolk carotenoid content and yolk color score, diminishing both antioxidant value and visual appeal, at dietary concentrations as low as 2,500 ppb.

3.4. Residues and Food Safety
After mycotoxins are absorbed, the host begins detoxification and excretion processes, while organ damage simultaneously occurs. Detoxification is primarily carried out by the liver, and accumulation occurs mainly in the liver and kidneys. However, accumulation in other tissues, including meat and eggs, has also been documented. AFB1 and its liver-derived metabolites, including AFM1, have been detected in eggs at transfer rates of approximately 0.05% of the dietary AFB1 intake; OTA transfers at ~0.15%; T-2 at ~0.10%; while DON, FB1, and ZEN transfer at lower rates.
3.5. Economic and Indirect Impacts
While ZEN is considered to have limited impact on commercial broiler performance, the situation is markedly different for breeder flocks. Acting primarily through its active hepatic metabolite α-zearalenol (α-ZOL), which has a higher affinity for estrogen receptors than the parent compound, ZEN may reduce fertility (impaired semen quality in roosters) and hatchability (increased embryo mortality, reduced chick quality at hatch). More broadly, mycotoxins negatively affect animal health, growth performance, and egg quality, leading to increased culling and veterinary costs, as well as lower selling prices for substandard eggs. As a concrete example, with Vietnam producing more than 20 billion eggs per year, even a 1–2% reduction in productivity or egg quality could translate into losses of tens of millions of USD annually – a scale of impact applicable across every major egg-producing nation in the region.
4. Key Considerations for Mycotoxin Risk Management
Managing mycotoxin risks requires an increasingly comprehensive and integrated approach. The “3F – from Feedmill, Farm to Fork” process is an integrated management framework developed by EW Nutrition in the region to prevent, trace, and mitigate mycotoxin-related risks for poultry producers and egg manufacturers.
4.1. Prevention at Source (Feedmill)
- Upon raw material intake: conduct sensory inspection, then perform proper sampling and test for mycotoxins using rapid test strips or ELISA.
- Storage: pay close attention to ambient relative humidity and temperature in warehouses/silos, as these two factors directly influence the moisture content and water activity (Aw) of stored materials, creating favorable conditions for the growth of Aspergillus spp. and/or Penicillium spp. (mold species capable of producing mycotoxins such as AFs, OTA, citrinin, patulin, etc. during storage). As practical targets: keep grain moisture below 14% for maize and wheat (below 10% for groundnut meal); Aspergillus spp. can proliferate at Aw ≥ 0.80, while Penicillium spp. remain active at Aw ≥ 0.78; maintaining Aw below these thresholds is the single most effective storage intervention.
- Finished feed samples from each batch must be properly collected and analyzed for multiple mycotoxins using ELISA or chromatographic methods (HPLC, LC-MS/MS, etc.). Retained samples should be stored under cool, dry conditions for a minimum of two weeks to enable analysis and traceability in the event of a subsequent incident.
- Periodically inspect hygiene of storage facilities and equipment (e.g., mixer, cooler, feed transport trucks from feedmill to farm).
- Develop preventive strategies against the adverse effects of mycotoxins on the health and performance of commercial laying hens, including supplementation with broad-spectrum solutions (Mastersorb Gold, etc.) adsorbing a diverse range of toxins and providing antioxidant support to mitigate oxidative stress.
4.2. Prevention at Farm Level
- Establish a routine hygiene monitoring program for housing facilities, particularly feed storage areas/silos and associated equipment (e.g., automated feeders, egg and manure conveyors, etc.).
- Feed samples from each batch at the farm level should also be properly collected and retained (under cool, dry conditions) for a minimum of two weeks for analysis and traceability should any subsequent issue arise.
- When animal health or performance issues arise and mycotoxicosis is suspected, in complement to analyzing retained feed samples, the analysis of mycotoxin residues in Dried Blood Spots (DBS), a technique developed by EW Nutrition and its partner, can be a valuable complementary measurement to make a diagnosis.
4.3. Food Safety at the Table (Fork)
Vietnamese Standard TCVN 1858:2018 stipulates that commercial chicken eggs must be clean, uncracked, undistorted, and free from spoilage; air cell depth must not exceed 6 mm (depending on grade); yolk must not be visibly off-center; and no off-odors are permitted. Eggs are graded based on both external and internal quality criteria. Additionally, Circular 34/2012/TT-BNNPTNT and food safety and hygiene regulations require traceability, Salmonella control, and monitoring of specific residues. The national technical regulation QCVN 01-190:2020/BNNPTNT on animal feed sets maximum limits for mycotoxins in feed raw materials. While Vietnam serves as a concrete example, analogous frameworks are in place across Southeast and South Asia, with many producers also referencing the Codex Alimentarius maximum levels for aflatoxins in food (4 µg/kg total AFs; 2 µg/kg AFB1) and EU feed maximum limits as de facto benchmarks for export-oriented operations.
According to the International Agency for Research on Cancer (IARC), AFB1 is classified as a Group 1 carcinogen (carcinogenic to humans), while OTA, AFM1, and FB1 are classified as Group 2B (possibly carcinogenic to humans). It is of particular concern that these mycotoxins are also highly heat-stable, they are not destroyed by cooking or standard food processing temperatures. Consequently, their residues in eggs represent an important aspect to be monitored and controlled before eggs reach the consumer’s table.
5. Conclusion
Mycotoxins are a critical factor affecting egg quality across all dimensions – external appearance, internal quality, food safety, and economic value. In the context of Vietnam’s egg industry, and the broader dynamic growth of egg production across Southeast and South Asia, moving toward modernization and export competitiveness, mycotoxin control is not merely a loss-reduction measure, but a strategy for sustainable competitive advantage. The sector requires close collaboration among feed manufacturers, poultry and egg producers, regulatory authorities, and scientific researchers to turn these challenges into opportunities for development.
In line with this broader direction, EW Nutrition has developed and is actively supporting the implementation of the integrated 3F Management Process (from Feedmill, Farm to Fork), grounded in scientific evidence and technology, to help protect poultry flocks, enhance egg quality, and ensure consumer safety across the region.
About the Author
Dr. Tran Si Trung holds PhD degrees in Food Safety & Quality in France and serves as Regional Technical Manager for Toxin Risk Management at EW Nutrition, Southeast Asia and Pacific. He specializes in mycotoxin risk assessment and feed quality management. For further information or technical inquiries, please contact EW Nutrition Vietnam.