EU Agricultural Outlook 2024-2035: Projected Trends and Challenges
by Ilinca Anghelescu, Global Director Marketing & Communications, EW Nutrition
The European Union (EU) agricultural sector is confronted with challenges and uncertainties stemming from the geopolitical risks, extreme weather events, and evolving market demand. The EU Agricultural Outlook 2024-2035, published last month, highlights the anticipated trends, challenges, and opportunities facing the sector over the medium term, given several considerations likely shaping the future.
Initial considerations for EU agricultural trends
Macroeconomic context
The EU’s real GDP growth is expected to stabilize, contributing to a stable economic environment for agriculture. Inflation rates are projected to return to the European Central Bank’s target of 2% by 2025. Exchange rates will see the Euro slightly appreciating against the US dollar, and Brent crude oil prices are anticipated to stabilize in real terms at approximately $102 per barrel by 2035.
However, despite optimistic declarations in the recent past, we have not solved world hunger. Population growth in lower-income parts of the world is leading to an unequal distribution and, after an initial dip, the number of people going to bed hungry is expected to rise again. Moreover, in the next ten years some improvements are foreseen but no massive changes are expected in the percentage of food groups and calories available per capita.
Climate change impact
Climate change is reshaping EU agriculture by affecting critical natural resources such as water and soil. Agroclimatic zones are shifting northwards, with implications for crop cultivation patterns. For example, regions traditionally suitable for wheat may increasingly shift focus to other crops better adapted to new climate conditions.
Consumer demand
Consumer awareness of sustainability is driving significant shifts in dietary preferences in the EU. The demand for plant proteins like pulses is increasing, while meat consumption, particularly beef and pork, is declining due to environmental and health concerns. Conversely, demand for fortified and functional dairy products is on the rise.
What are the projected agricultural trends in 2024-2035?
Arable crops
Land use: While the total agricultural land in the EU remains stable, a shift in crop focus is anticipated. Land allocated for cereals and rapeseed is expected to decline, making way for soya beans and pulses due to reduced feed demand and policy incentives for plant proteins.
Cereals: Production of cereals, including wheat, maize, and barley, is forecast to stabilize with minor yield increases due to advancements in precision farming and digitalization. Wheat production is set to recover after an expected dip in 2024.
Dairy Sector
Milk production: Although milk yields are projected to increase due to improved genetics and farming practices, the decline in the dairy cow herd will result in a slight overall reduction in milk production by 2035.
Dairy products: The production of cheese and whey will grow steadily, driven by domestic and international demand. Conversely, the consumption of drinking milk is expected to decline, while demand for fortified and functional dairy products grows.
Meat Sector
Beef and veal: Beef production is expected to decrease by 10%, with the EU cow herd shrinking by 3.2 million head by 2035. This decline is attributed to sustainability concerns, high production costs, and changing consumer preferences. Beef consumption is also projected to decline, driven by high prices and a preference for plant-based alternatives.
Pig meat: The sector faces a projected annual production decline of 0.9%, equating to a reduction of nearly 2 million tons compared to 2021-2023 levels. This trend is largely influenced by concerns over sustainability and a declining preference for fatty meats.
Poultry: In contrast, poultry production is forecast to increase due to its healthier image, lower cost, and minimal cultural or religious constraints. However, the growth rate will be slower than in the previous decade.
Upcoming challenges in agriculture
Climate Resilience
The increasing frequency of extreme weather events requires investments in resilient farming practices. Adoption of precision farming and crop diversification is critical to mitigate climate impacts. However, if existing policies are further implemented, greenhouse gas emissions are expected to see a significant decline.
Policy Frameworks
The Common Agricultural Policy (CAP) plays a pivotal role in steering the sector toward sustainability. However, farmers face challenges in adapting to stricter environmental regulations and securing sufficient funding for transitions. The recent Mercosur agreement has already stirred dissent in EU countries that fear unfettered competition without similar policy regulations.
Market Dynamics
Global trade tensions and competition in agricultural markets pose significant risks. While the EU remains a net exporter, dependence on imports for certain crops, such as soya beans, highlights vulnerabilities in supply chains.
In a weather-shock scenario for the EU feed supply chain, the report highlights that increased feed prices would drive up retail meat prices by 10% for poultry and pork producers, and 5% for beef and veal producers. The increase would be less abrupt for retail prices, rising by 3% for pork, and 4% for poultry meat. Producers need to be mindful of the absorbed costs of these potential shocks.
Conclusion
The EU agricultural sector must continue to balance productivity, sustainability, and consumer preferences. While advancements in technology and policy frameworks offer pathways to resilience, addressing challenges such as climate change and market dynamics will be critical to achieving long-term goals.
BioStabil Plus improves grass silage quality and cattle profitability
by Dr. Vesna Jenkins, Global Product Manager, EW Animal Care
Making silage enables the farmer to store forage, providing a cost-effective feed when required. From silage making through to feeding out, however, the challenge is to ensure that valuable dry matter, energy and protein are not lost. Any losses would require supplementation from other sources at extra cost. In the case of protein, farmers would need to purchase additional soybean meal, for example, to maintain cow productivity.
Clostridia: The Main Villain
One of the greatest challenges to making good silage is the presence of Clostridia bacteria, which can negatively impact animal health, performance and profitability. These bacteria pose a health risk to both beef and dairy cattle and can negatively influence cheese quality through the late blowing defect.
During the ensiling process, Clostridia break down protein, reducing silage nutritional value, and produce butyric acid, which decreases silage palatability and affects feed intake. Clostridia can easily enter through soil contamination and thrive in forage with low dry matter, high buffering capacity, or lower levels of soluble carbohydrates and nitrate.
Negative impacts of Clostridia
Health risk to cattle
Reduced nutritional value of silage
Declined feed intake, leading to diminished productivity
Late blowing defect in cheese
Trial results
A recent scientific trial by the Swedish University of Agricultural Sciences (SLU) tested the effect ofBioStabil Plus silage inoculant on difficult to ensile grass-clover forage (28% DM) challenged with Clostridia. The research demonstrated a clear effect of BioStabil Plus on multiple parameters.
The application of BioStabil Plus to glass-clover forage resulted in:
Improved dry matter (DM) retention (Figure 1)
Enhanced protein preservation (Figure 2)
Both outcomes contribute to feed cost savings.
Figure 1. Significantly lower dry matter loss in grass-clover silage treated with BioStabil Plus (90 days past ensiling, P<0.001). Source: Swedish University of Agricultural Sciences and EW Nutrition.
Figure 2. Less ammonia-N with BioStabil Plus, significantly higher protein preservation (90 days past ensiling, P<0.001). Source: Swedish University of Agricultural Sciences and EW Nutrition.
Benefits of BioStabil Plus
Protection Against Nutrient Loss – BioStabil Plus protects against dry matter, energy, and protein losses in the fermentation period. It contains the rapid-growing lactic-acid-producing homofermentative strain L. plantarum DSM 19457, ensuring sufficient lactic acid production for a rapid pH drop in ensiled forage (Figure 3).
Figure 3. Lower pH in grass-clover silage challenged with Clostridia and treated with BioStabil Plus compared to Clostridia challenged forage without inoculant (90 days past ensiling, P<0.001). Source: Swedish University of Agricultural Sciences and EW Nutrition.
Reduction of Clostridial Load – BioStabil Plus reduces the Clostridial load as evidenced by significantly lower butyric acid production (Figure 4). Lower butyric acid content maintains silage palatability, feed intake, and avoids final dairy product quality issues.
Figure 4. Significantly lower butyric acid with BioStabil Plus showing minimal Clostridia presence (90 days past ensiling, P<0.001). Source: Swedish University of Agricultural Sciences and EW Nutrition.
Enhanced Aerobic Stability – BioStabil Plus contains heterofermentative strains L. buchneri DSM 19455 and L. brevis DSM 23231, producing an optimal level of acetic acid for enhanced aerobic stability during the feed-out phase. An EFSA scientific opinion on L. brevis DSM 23231 specifically outlines its ability to reduce Clostridia risk.
Producing high-quality, palatable, well-preserved silage ensures that the investment in silage making is not wasted. Most importantly, the preserved energy and protein maximize profitability through higher production of milk or meat and generate feed cost savings that support producers’ margins.
Contact your local EW Nutrition representative to access valuable resources and advice on all aspects of optimized silage management.
Antimicrobial resistance in animal production workers, a serious challenge
With 73% of human-use antibiotics also used in food-animal production, antimicrobial resistance (AMR) is a pressing global health concern, particularly in contexts where humans and animals are in close proximity, such as in animal production facilities. This issue is exacerbated by the widespread use of antibiotics in livestock farming, which not only promotes resistance in bacteria but also poses direct risks to farm workers.
Antimicrobial resistance in farm workers in Denmark
In Denmark, a country renowned for its robust agricultural monitoring systems, significant strides have been made in tracking AMR. A comprehensive report from 2015 emphasized the occurrence of antimicrobial-resistant bacteria, particularly in livestock-associated methicillin-resistant Staphylococcus aureus (LA-MRSA). The Danish Integrated Antimicrobial Resistance Monitoring and Research Program (DANMAP) highlighted that farm workers frequently came into contact with these resistant pathogens, which posed occupational hazards and public health challenges (Bager et al., 2015). The program found that 88% of pigs carried LA-MRSA, and farm workers had significantly elevated exposure risks, particularly in intensive swine operations (DANMAP 2015 Report).
Antimicrobial resistance in farm workers in the US
Studies in the United States have revealed even more alarming statistics. Farm workers in intensive animal farming environments were found to be 32 times more likely to develop antibiotic-resistant infections than the general population. This increased risk was attributed to prolonged exposure to resistant bacteria and antibiotic residues in animal feed and the environment (Silbergeld et al., 2008). The close interaction between humans and animals in confined spaces fosters the transfer of resistant genes, making these workers a vulnerable group.
Mechanisms of resistance spread
The spread of AMR from livestock to humans can occur through several pathways:
Direct contact: Handling animals and exposure to manure or bodily fluids.
Contaminated food: Consumption of undercooked or improperly handled meat products.
Environmental contamination: Water and soil contaminated with antibiotics or resistant bacteria.
What can be done?
Even in countries where antimicrobials reduction legislation has been in place for almost two decades, such as Germany or Sweden, new resistance cases are constantly discovered. In supermarkets around the world, meat contaminated with antibiotic-resistant superbugs is still a common occurrence. And in antibiotic resistance hot spots, “from 2000 to 2018, P50 increased from 0.15 to 0.41 in chickens—meaning that 4 of 10 antibiotics used in chickens had resistance levels higher than 50%. P50 rose from 0.13 to 0.43 in pigs and plateaued between 0.12 and 0.23 in cattle” (Dall, 2019). These hot spots are spread across the globe, from south and northeast India, northeast China, north Pakistan, Iran, and Turkey, to the south coast of Brazil, Egypt, the Red River Delta in Vietnam, and areas surrounding Mexico City, Johannesburg, and more recently Kenya and Morocco.
Globally, antimicrobial use in animals is projected to increase by 67% by 2030, especially in low- and middle-income countries where regulatory frameworks are weaker. Denmark provides a successful model for mitigating these risks. Policies such as the “Yellow Card” scheme have reduced antibiotic use in pigs by promoting alternative husbandry practices and strict monitoring. This approach has also reduced the prevalence of resistant bacteria in animal populations, offering a replicable strategy for other nations (Alban et al., 2017).
Recommendations for mitigation
Strengthening surveillance: Programs like DANMAP should be implemented globally to monitor antibiotic usage and resistance trends in animals and humans.
Reducing antibiotic use: Phasing out non-therapeutic uses of antibiotics, particularly as growth promoters, and avoiding Critically Important Antimicrobials for Human Medicine.
Protecting workers: Providing personal protective equipment (PPE) and regular health screenings for farm workers.
Public awareness: Educating communities about the risks of AMR and promoting safe food handling practices.
The evidence from Denmark and the U.S. underscores the urgent need to address AMR in animal production settings. Protecting farm workers from AMR not only safeguards their health but also prevents the spread of resistant pathogens across the wider public.
References
Bager, F., et al. (2015). DANMAP 2014: Use of antimicrobial agents and occurrence of antimicrobial resistance in bacteria from food animals, food, and humans in Denmark. Retrieved from DANMAP Report.
Silbergeld, E. K., Graham, J., & Price, L. B. (2008). Industrial food animal production, antimicrobial resistance, and human health. Annual Review of Public Health, 29(1), 151-169.
Alban, L., et al. (2017). Assessment of the risk to public health due to use of antimicrobials in pigs—An example of pleuromutilins in Denmark. Frontiers in Veterinary Science, 4, 74. DOI.
EU publishes Short-term Outlook for Agricultural Markets (Autumn 2024)
The EU’s Short-term Outlook for Agricultural Markets (Autumn 2024) reveals significant challenges in agriculture, with adverse weather, geopolitical instability, and fluctuating trade conditions impacting production. The report identifies declining cereal and oilseed outputs, particularly for soft wheat and maize. Meanwhile, milk production is expected to remain stable despite a shrinking cow herd, and the meat sector shows mixed trends, with poultry production rising but pigmeat and beef facing structural challenges.
EU’s Short-term Outlook for Autumn 2024 highlights the following key findings collectively shaping EU agricultural markets:
Weather conditions: Severe drought in Southern and Eastern Europe and excess rainfall in Northwestern regions have reduced cereal and oilseed yields.
Energy and input costs: Inflation is stabilizing but remains high, with elevated prices for inputs like fertilizers and energy.
Geopolitical tensions: The ongoing war in Ukraine and trade disputes are disrupting supply chains and impacting exports.
Global demand: While global agricultural demand is recovering, trade flows face disruptions due to regulatory changes and volatile market conditions.
Cereals
The EU cereal production in 2024/25 is projected at 260.9 million tons, approximately 7% below the 5-year average. This marks the lowest production in the past decade, driven by unfavorable weather conditions, including excessive rain in Northwestern Europe, which impacted planting, particularly for soft wheat, and drought in Southern and Eastern regions, severely affecting maize yields. Production of soft wheat and maize is expected to decline year-on-year by 9.5% and 4%, respectively. On the other hand, barley and durum wheat production are increasing by about 6% and 3%, respectively, compared to the previous year.
EU cereal exports are projected to decline by 22% year-on-year due to reduced production and quality issues. At the same time, domestic demand remains relatively stable, with animal feed consumption holding steady as livestock production stagnates. In terms of prices, cereal prices fell throughout 2024, pressuring farmers’ cash flow, which could hinder their ability to afford inputs such as fertilizers in the coming year.
Milk and Dairy Products
The EU milk market is expected to see relatively stable supply, despite a continuously shrinking cow herd. Milk yields have increased, compensating for the herd’s decline. Milk prices are forecast to stabilize after a period of volatility in the past few years, remaining above historical averages, and input costs for farmers, such as feed and energy, are showing signs of stabilizing, allowing for a potential improvement in farmer margins.
Despite the stability in milk supply, demand for dairy products continues to show mixed trends, influenced by shifts in consumer preferences and trade dynamics. The balance of milk supply and prices could provide an opportunity for dairy farmers to recover some profitability after several challenging years.
In the dairy products sector, cheese and butter continue to dominate EU production, with butter production projected to rise slightly in 2024, driven by stable milk supplies and strong domestic demand. The demand for butter in the global market remains relatively strong, although competition is rising.
Cheese production is also expected to remain stable, reflecting a balance between domestic and export markets. The cheese sector has seen steady growth over the years, supported by increasing consumer demand for premium and specialty cheeses. The demand for skimmed milk powder (SMP) and whole milk powder (WMP) is projected to remain subdued due to fluctuating global demand, particularly from key markets such as China, although some growth is expected in non-European markets.
Meat Products
The meat sector in the EU remains a mixed picture, with structural changes and external factors shaping production and trade in 2024.
Beef and Veal: Beef production continues to face structural decline due to a shrinking herd size, with the sector stabilizing but at lower levels of production. The demand for EU beef remains relatively high, and exports are increasing, but domestic production is likely to remain constrained by environmental and economic pressures. Additionally, the number of animals has been declining consistently, reflecting longer-term trends within the EU beef industry.
Pigmeat: The EU pigmeat sector is facing diverse challenges, with some countries recovering from production setbacks, while others struggle with ongoing disease outbreaks and economic issues. The overall EU pigmeat production is expected to decline slightly, and exports have become less competitive, particularly with reduced demand from key markets such as China. However, opportunities exist in other Asian countries, where EU exporters are gaining ground. Domestically, consumption is forecast to decrease slightly, reflecting shifting consumer preferences toward plant-based alternatives and poultry.
Poultry: Poultry production is expected to rise, driven by strong domestic demand and favorable export conditions. The EU poultry sector has shown resilience, with increasing production and exports, despite higher input costs. Poultry remains a preferred source of protein for consumers, especially as prices for other meats rise. The sector continues to grow in competitiveness on the international stage, with exports expected to increase in 2024 despite the challenges posed by higher EU prices.
Sheep and Goat Meat: Production of sheep and goat meat continues to decline due to the structural reduction of flocks across the EU. High EU prices have made sheep and goat meat less competitive on the global market, reducing export opportunities. Domestically, consumption remains stable but at lower levels than other meat types. The ongoing structural decline in the sector highlights long-term challenges related to animal health, productivity, and market competitiveness.
Volatility and challenges persist
The report highlights the ongoing challenges faced by the cereals, dairy, and meat sectors. Weather conditions and global trade dynamics are shaping the future of EU agriculture, with many sectors grappling with production declines and shifting market demands. Despite these challenges, opportunities exist for some areas of growth, particularly in dairy and poultry, where rising consumer demand and stable supply conditions offer optimism for the future.
The art of silage: A guide to maximizing quality and nutritional value
by Vesna Jenkins, Global Product Manager, Biomin BioStabil
Silage quality directly impacts animal health and farm profitability. This guide delves into the scientific principles and practical steps necessary to produce silage of the highest caliber.
Optimal Dry Matter
The journey to exceptional silage begins with harvesting at the ideal dry matter percentage. This critical timing ensures the preservation of yield and energy content. Striking the right balance is key; harvesting too early can lead to nutrient-poor silage, while too late can compromise the forage’s structural integrity. Aim for a dry matter content of 32-38% depending on forage type for optimal results.
Wilting Wisdom
When wilting is part of the process (e.g. grass, clover or alfalfa silage), efficiency is paramount. Achieving the desired dry matter in just a few hours help to prevent spoilage and retain the forage’s nutritional value. It’s a delicate dance between removing excess moisture and maintaining the feed’s quality.
Ensiling Excellence
Post-harvest, the clock is ticking. Compacting and sealing the forage within 24 to 48 hours is vital to create an anaerobic environment. This step is crucial to ensure anaerobic conditions for optimal fermentation. Pack the silage in thin layers with heavy enough machinery such as dual wheeled heavy tractors to achieve optimal dry matter density of around 250 kg per cubic meter. Pay special attention to the edges for even compaction. Once filled, seal the clamp with high quality overlapping sheets ensuring the edges are weighted down to prevent air ingress.
Rapid Acidification
The role of silage inoculants cannot be overstated. The proven science of the silage inoculant Biomin® BioStabil accelerates the pH drop, locking in dry matter, energy, and protein. This rapid acidification is a defense mechanism against pathogenic bacteria and mycotoxin producing fungi, ensuring the silage remains safe and nutritious.
Feed-Out Finesse
Proper management of the clamp face is crucial to prevent spoilage and ensure livestock health. Cut sufficient depth fromthe clamp face daily to prevent newly exposed silage near the face from having time to spoil. Maintain a smooth and clean silage face to minimize spoilage. Spilled debris on the ground can easily go moldy, presenting hazards for animals if fed out. Use the proven silage inoculant Biomin® BioStabil for longer aerobic stability in the silage and TMR.
Silage making is both a science and an art. By following these guidelines and choosing the right inoculant, farmers can secure the nutritional integrity of their forage.
Consult with an EW Nutrition representative to select the perfect BioStabil inoculant tailored to your forage type and conditions, and elevate your silage from good to great.
Sustainable livestock farming: Progress since 1950
by Ilinca Anghelescu, Global Director Marketing Communications, EW Nutrition
Summary
Global GHG Emissions and Agriculture:
Agriculture-related emissions account for 31% of global anthropogenic emissions, with a growing share coming from food-related activities outside traditional farming, such as processing and transportation.
This represents a significant decrease from the 1950s when agriculture contributed to 58% of global emissions, a decrease largely due to the increased use of fossil fuels.
Population Growth and Emissions:
The global population has increased by 220% since 1950, leading to a threefold increase in agri-food emissions, now totaling 9-10 billion metric tons of CO2-equivalent annually.
Meat Production Growth:
Meat production has seen a 690% increase since 1950, driven by population growth, economic development, urbanization, technological advancements, and intensification of livestock production.
Technological improvements have significantly increased livestock yield, including higher carcass weights, improved feed efficiency, and greater output per animal.
Feed Conversion Ratio (FCR) as a Sustainability Metric:
FCR, which measures the efficiency of feed conversion into body mass, has improved dramatically for poultry, pigs, and cattle since the 1950s.
Improved FCR contributes to more efficient resource use, reduced environmental impact, better animal welfare, and economic viability.
Livestock Emissions and Land Use:
Livestock-related emissions have increased by 14% since 2000, with a significant portion coming from enteric fermentation in ruminants and land use changes.
Pasture and grazing land have expanded slightly, while the land for feed crops has increased substantially due to intensified livestock production.
Food Loss and Its Impact on Sustainability:
Food loss, occurring primarily before the consumer stage, remains stable at around 13%. Reducing food loss is critical for improving food security, economic efficiency, and minimizing the environmental footprint of livestock production.
Future Strategies for Sustainability:
The article emphasizes the need for ongoing investment in technology, optimization of feed efficiency, sustainable land use, and improved methods for tracking and reducing emissions to ensure the future sustainability of livestock farming.
As the global demand for animal products continues to rise, so do various claims about the impact of agriculture on greenhouse gas emissions. A study commissioned by the United Nations’ Food and Agriculture Organization (FAO) concluded that, according to the most recent data, agri-food system emissions totaled 16.5 billion metric tons of CO2 equivalent, representing 31% of global anthropogenic emissions.
Of these 31%, the most important trend highlighted by FAO was the “increasingly important role of food-related emissions generated outside of agricultural land, in pre- and post-production processes along food supply chains”. The food supply chain (food processing, packaging, transport, household consumption and waste disposal) is thus set to become the top GHG emitter, above farming and land use.
How bad is 31%?
While 31% is a large figure, even this estimate represents a significant decrease from the 1950s, when agri-food emissions constituted approximately 58% of total anthropogenic emissions: “From 1850 until around 1950, anthropogenic CO2 emissions were mainly (>50%) from land use, land-use change and forestry”, states the latest IPCC report.
Figure 1. Source: IPCC AR6 Report, 2023. LULUCF = Land Use, Land-Use Change and Forestry
As the IPCC graph in Figure 1 indicates, the percentage decrease is mostly due to the rising prevalence of oil and coal in CO2 emissions over the recent decades, as shown in Figure 2 below.
Annual greenhouse gas (GHG) emissions worldwide from 1990 to 2022, by sector (in million metric tons of carbon dioxide equivalent)
Total population and agri-food emission changes, 1950 – today
The global population increased by approximately 220%, from 2.5 billion in 1950 to 8 billion in 2023. In the meantime, estimates suggest that, in the 1950s, agri-food systems were responsible for approximately 2-3 billion metric tons of CO2-equivalent (CO2e) emissions per year. This figure includes emissions from livestock, rice paddies, fertilizer use, and land-use change (e.g., deforestation for agriculture).
Assessments generally agree that today’s agri-food systems contribute approximately 9-10 billion metric tons of CO2e annually, a threefold increase from 1950. This includes emissions from agriculture (e.g., livestock, crop production), food processing, transportation, and land-use changes.
This increase is consistent with FAO’s new findings, of food chain climbing to the top of agri-food emitters.
But where did these increased emissions come from?
A look at the graph below gives us an indication: world poverty rate decreased massively between 1950 and today. While COVID brought a setback, the historical data would clearly indicate a correlation between the increased output in agri-food systems and the decreased rate of poverty.
How did poverty rates decline so steeply? The reasons lie, to a large extent, in technological innovation, especially in genetics and farm management, and in the increased apport of plentiful and affordable meat protein to the world. The numbers below build an image of an industry that produces better, more, and cheaper.
Global meat production: 1950 vs. Present
Then…
In 1950, the estimated total meat production was of approximately 45 million metric tons.
Key Producers: The United States, Europe, and the Soviet Union were the primary producers of meat. Types of Meat: Production was largely dominated by beef and pork, with poultry being less significant.
…and now
Now, the total meat production lies somewhere around 357 million metric tons (as of recent data from FAO)., representing a 53% increase from 2000 and a staggering 690% increase from 1950.
Key Producers: Major producers include China, the United States, Brazil, and the European Union. Types of Meat: Significant increases in poultry production, with pork remaining a leading source of meat, especially in Asia. Beef production has also increased, but at a slower rate than poultry and pork.
Factors contributing to increased meat production
Population Growth: The world population has grown from approximately 2.5 billion in 1950 to over 8 billion today, driving increased demand for meat.
Economic Growth and Urbanization: Rising incomes and urbanization have led to shifts in economic power and dietary preferences, with more people consuming higher quantities of meat, especially in developing countries.
Technological Advancements: Improvements in animal breeding, feed efficiency, and production systems have increased the efficiency and output of meat production.
Intensification of Livestock Production: The shift from extensive to intensive livestock production systems has allowed for higher meat yields per animal.
Global Trade: Expansion of global trade in meat and meat products has facilitated the growth of production in countries with comparative advantages in livestock farming.
Livestock yield increase, 1950 to the present
The increase in livestock yield for cattle, pigs, and chickens between 1950 and the present has been significant due to advances in breeding, nutrition, management practices, and technology.
Beef
1950s
Average Carcass Weight: In the 1950s, the average carcass weight of beef cattle was about 200 to 250 kilograms (440 to 550 pounds).
Dressing Percentage: The dressing percentage (the proportion of live weight that becomes carcass) was typically around 50-55%.
Present Day
Average Carcass Weight: Today, the average carcass weight of beef cattle is approximately 300 to 400 kilograms (660 to 880 pounds).
Dressing Percentage: The dressing percentage has improved to about 60-65%.
Increase in Beef Cattle Yield
Increase in Carcass Weight: The average carcass weight has increased by about 100 to 150 kilograms (220 to 330 pounds) per animal.
Improved Dressing Percentage: The dressing percentage has increased by about 5-10 percentage points, meaning a greater proportion of the live weight is converted into meat.
Dairy
1950s
Average Milk Yield per Cow: Approximately 2,000 to 3,000 liters per year, depending on the region.
Present Day
Average Milk Yield per Cow: Approximately 8,000 to 10,000 liters per year globally, with some countries like the United States achieving even higher averages of 10,000 to 12,000 liters per year.
Increase in Milk Yield:: Milk yield per cow has increased about 4-5 times due to genetic selection, improved nutrition, technological advancements, and better herd management.
Chickens (Layers)
1950s
Average Egg Production per Hen: In the 1950s, a typical laying hen produced about 150 to 200 eggs per year.
Present Day
Average Egg Production per Hen: Today, a typical laying hen produces approximately 280 to 320 eggs per year, with some high-performing breeds producing even more.
Increase in Egg Yield: The average egg production per hen has increased by approximately 130 to 170 eggs per year.
Chickens (Broilers)
1950s
Average Yield per Bird: In the 1950s, broiler chickens typically reached a market weight of about 1.5 to 2 kilograms (3.3 to 4.4 pounds) over a growth period of 10 to 12 weeks.
Present Day
Average Yield per Bird: Today, broiler chickens reach a market weight of about 2.5 to 3 kilograms (5.5 to 6.6 pounds) in just 5 to 7 weeks.
Increase in Yield: The average weight of a broiler chicken has increased by approximately 1 to 1.5 kilograms (2.2 to 3.3 pounds) per bird. Additionally, the time to reach market weight has been nearly halved.
Factors contributing to yield increases
Genetic Improvement:
Selective Breeding: Focused breeding programs have developed chicken strains with rapid growth rates and high feed efficiency, significantly increasing meat yield.
Nutrition:
Optimized Feed: Advances in poultry nutrition have led to feed formulations that promote faster growth and better health, using balanced diets rich in energy, protein, and essential nutrients.
Management Practices:
Housing and Environment: Improved housing conditions, including temperature and humidity control, have reduced stress and disease, enhancing growth rates.
Technological Advancements:
Automation: Automation in feeding, watering, and waste management has improved efficiency and bird health.
Health Monitoring: Advances in health monitoring and veterinary care have reduced mortality rates and supported faster growth.
Feed Conversion Efficiency:
Improved Feed Conversion Ratios (FCR): The amount of feed required to produce a unit of meat has decreased significantly, making production more efficient.
Why Feed Conversion Ratio is a sustainability metric
Feed Conversion Ratio (FCR) is a critical metric in livestock production that measures the efficiency with which animals convert feed into body mass. It is expressed as the amount of feed required to produce a unit of meat, milk, or eggs. Advances in nutrition and precision feeding allow producers to tailor diets that optimize FCR, reducing waste and improving nutrient uptake. Also, breeding programs focused on improving FCR can lead to livestock that naturally convert feed more efficiently, supporting long-term sustainability.
Poultry (Broilers): From the 1950s, improved from approximately 4.75 kg/kg to 1.7 kg/kg.
Pigs: From the 1950s, improved from about 4.5 kg/kg to 2.75 kg/kg.
Cattle (Beef): From the 1950s, improved from around 7.5 kg/kg to 6.0 kg/kg.
Figure 4. Evolution of FCR from 1950
FCR is crucial for livestock sustainability for several reasons, as shown below.
1. Resource efficiency
– Feed Costs: Feed is one of the largest operational costs in livestock production. A lower FCR means less feed is needed to produce the same amount of animal product, reducing costs and improving profitability.
– Land Use: Efficient feed conversion reduces the demand for land needed to grow feed crops, helping to preserve natural ecosystems and decrease deforestation pressures.
– Water Use: Producing less feed per unit of animal product reduces the water needed for crop irrigation, which is crucial in regions facing water scarcity.
2. Environmental impact
– Greenhouse Gas Emissions: Livestock production is a significant source of greenhouse gases (GHGs), particularly methane from ruminants and nitrous oxide from manure management. Improved FCR means fewer animals are needed to meet production goals, reducing total emissions.
– Nutrient Runoff: Efficient feed use minimizes excess nutrients that can lead to water pollution through runoff and eutrophication of aquatic ecosystems.
3. Animal welfare
– Health and Growth: Optimizing FCR often involves improving animal health and growth rates, which can lead to better welfare outcomes. Healthy animals grow more efficiently and are less susceptible to disease.
4. Economic viability
– Competitiveness: Lowering FCR improves the economic viability of livestock operations by reducing input costs and increasing competitiveness in the global market.
– Food Security: Efficient livestock systems contribute to food security by maximizing the output of animal protein relative to the input of resources.
Improving FCR is essential for achieving sustainability in livestock production. It leads to more efficient resource use, reduced environmental impact, enhanced economic viability, and supports the well-being of animals. As global demand for animal products continues to rise, optimizing FCR will be crucial in balancing production with the need to protect and preserve natural resources.
“The eight warmest years on record since 1961 (and in fact since the beginning of observations in 1880) are all within the eight-year period of 2015–2022. Europe is the region where the temperature change has been the highest in 2022 (and also for most of the 2000–2022 period), with 2.23 °C, followed by Asia (1.75 °C), the Americas (1.05 °C), Africa (1.01 °C) and Oceania (0.8 °C). The average temperature change in the 2010s was 1.25 °C, compared to 0.96 °C in the 2000s.” FAOSTAT 2023
Livestock emissions
Livestock emissions can be direct (farm-gate) or indirect (land use). Pre- and post-production emissions are considered separately, since they refer to emissions from manufacturing, processing, packaging, transport, retail, household consumption, and waste disposal.
Global farm-gate emissions (related to the production of crops and livestock) grew by 14% between 2000 and 2021, to 7.8 Gt CO2 eq, see below. 53% come from livestock-related activities, and the emissions from enteric fermentation generated in the digestive system of ruminant livestock were alone responsible for 37 percent of agricultural emissions (FAOSTAT 2023).
Land use emissions contribute a large share to agricultural emissions overall, especially through deforestation (~74% of land-use GHG emissions). The numbers have declined in recent years, to a total of 21% reduction between 2000 and 2018.
The other side of the coin is represented by the increased land usage for livestock, either directly for grazing or indirectly for feed crops.
1. Pasture and grazing land
1950: Approximately 3.2 billion hectares (7.9 billion acres) were used as permanent pastures.
Present: The area has increased to around 3.5 billion hectares (8.6 billion acres).
Change: An increase of about 0.3 billion hectares (0.7 billion acres).
2. Land for Feed Crops
1950: The land area dedicated to growing feed crops (such as corn and soy) was significantly less than today due to lower livestock production intensities and smaller scale operations. Feed crops likely accounted for about 200-250 million hectares of the cropland, although figures are evidently difficult to estimate.
Present: Of the approx. 5 billion hectares of land globally used for agriculture, about 1.5 billion hectares are dedicated to cropland.
The increase in cropland hectares is a direct consequence of the intensification of demand for livestock production. To keep these numbers in check, it is essential that producers strive to use as little feed as possible for as much meat yield as possible – and this directly relates to a key metric of the feed additive industry: Feed Conversion Ratio, mentioned above.
The role of food loss in livestock sustainability
The Food and Agriculture Organization (FAO) of the United Nations definesfood loss as the decrease in quantity or quality of food resulting from decisions and actions by food suppliers in the chain, excluding retail, food service providers, and consumers. Food loss specifically refers to food that gets spilled, spoiled, or lost before it reaches the consumer stage, primarily taking place during production, post-harvest, processing, and distribution stages.
Food loss is currently estimated to be relatively stable over the last decades, at around 13%.
Key aspects of food loss
Stages of Food Loss:
Production: Losses that occur during agricultural production, including damage by pests or diseases and inefficiencies in harvesting techniques.
Post-Harvest Handling and Storage: Losses that happen due to inadequate storage facilities, poor handling practices, and lack of proper cooling or processing facilities.
Processing: Losses during the processing stage, which may include inefficient processing techniques, contamination, or mechanical damage.
Distribution: Losses that occur during transportation and distribution due to poor infrastructure, inadequate packaging, and logistical inefficiencies.
Quality and Quantity:
Quality Loss: Refers to the reduction in the quality of food, affecting its nutritional value, taste, or safety, which may not necessarily reduce its quantity.
Quantity Loss: Refers to the actual reduction in the amount of food available for consumption due to physical losses.
Exclusions:
Retail and Consumer Level: Food loss does not include food waste at the retail or consumer levels, which is categorized as food waste. Food waste refers to the discarding of food that is still fit for consumption by retailers or consumers.
Importance of reducing food loss
Every step along the production chain, each action taken to preserve feed, increase yield, ensure stable and high meat quality, can contribute to reducing food loss and ensuring that animal protein production stays sustainable and feeds the world more efficiently.
Food Security: Reducing food loss can help improve food availability and access, particularly in regions where food scarcity is a concern. Where we thought we were on our way to eradicate world hunger, recent upticks in several regions show us that progress is not a given.
Economic Efficiency: Minimizing food loss can improve the efficiency and profitability of food supply chains by maximizing the utilization of resources.
Environmental Impact: Reducing food loss helps to decrease the environmental footprint of food production by lowering greenhouse gas emissions and minimizing land and water use. This is all the more important in regions where world hunger shows signs of going up. Perhaps not by coincidence are these regions some of the most affected by climate change.
By understanding and addressing the causes of food loss, stakeholders across the food supply chain can work towards more sustainable and efficient food systems.
What’s next?
Improving production practices and technology
Investment in research and development of new technologies that enhance livestock production efficiency and reduce environmental impact is vital for the future sustainability of the sector.
India is a good illustration of room to grow. If we look at cow milk alone, India, with a headcount of approximately 61 million animals, has a total milk production that is neck-and-neck with the United States, whose dairy cow headcount is in the neighborhood of 9.3 million. India’s milk yield sits around 1,600 liters/animal/year, compared to the US’s average of 10,700 liters.
Continued focus on improving FCR through genetic selection, optimized nutrition, and advanced management practices will be crucial for reducing the environmental footprint of livestock production.
Promoting Sustainable Land Use
Strategies to balance the need for increased livestock production with sustainable land use practices are essential. This includes adopting agroecological approaches and improving the efficiency of feed crop production.
Reducing Food Loss
Stakeholders across the food supply chain must prioritize reducing food loss through improved storage, transportation, and processing technologies. This will help ensure that livestock production contributes effectively to global food security.
Enhancing Emission Tracking and Reporting
There is a need for standardized methods for collecting and reporting data on GHG emissions in agriculture. This will enable more accurate assessments and the development of targeted strategies for emission reductions.
References
Bell, D. D. (2002). Laying hens in the U.S. market: An appraisal of trends from the beginning of the 20th century to present. Poultry Science, 81(5), 485-490. https://doi.org/10.1093/ps/81.5.485
Crippa, M., Solazzo, E., Guizzardi, D., Monforti-Ferrario, F., Tubiello, F. N., Leip, A., … & Janssens-Maenhout, G. (2022). Greenhouse gas emissions from food systems: building the global food system emissions database (GFED). Earth System Science Data, 14(4), 1795-1821. https://essd.copernicus.org/articles/14/1795/2022/essd-14-1795-2022.pdf
Goldewijk, K. K., & Verburg, P. H. (2013). Per-capita estimations of long-term historical land use and the consequences for global change research. Global Environmental Change, 23(4), 1166-1175. https://doi.org/10.1016/j.gloenvcha.2013.04.001
Kusuma, A. B., Laga, W. R., & Purnomo, H. (2022). Climate Change and Livestock Farming: Strategies for Mitigation and Adaptation. MDPI, 12(10), 1554. https://www.mdpi.com/2077-0472/12/10/1554
Roche, J. R., Friggens, N. C., Kay, J. K., Fisher, M. W., Stafford, K. J., & Berry, D. P. (2013). Invited review: Body condition score and its association with dairy cow productivity, health, and welfare. Animal Frontiers, 3(4), 23-29. https://doi.org/10.2527/af.2013-0032
Thornton, P. K., & Herrero, M. (2015). Impacts of climate change on the livestock food supply chain; a review of the evidence. Frontiers in Veterinary Science, 2, 93. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4686767/
Zuidhof, M. J., Schneider, B. L., Carney, V. L., Korver, D. R., & Robinson, F. E. (2014). Evolution of the modern broiler and feed efficiency. Annual Review of Animal Biosciences, 2(1), 47-71. https://doi.org/10.1146/annurev-animal-022513-114132
Global antimicrobial use in livestock farming: A revised estimate
Antimicrobial resistance (AMR) poses a significant threat to global health, driven by the overuse and misuse of antibiotics in both human medicine and livestock farming. In livestock farming, antimicrobials are still used extensively for therapeutic and non-therapeutic purposes. However, estimates of the quantities used per species are notoriously hard to derive from fragmented, incomplete, or unstandardized data around the world.
A recent article (“Global antimicrobial use in livestock farming: an estimate for cattle, chickens, and pigs”, Animal, 18(2), 2024) attempts to update the figures by estimating global biomass at treatment of cattle, pigs, and chickens, considering distinct weight categories for each species in biomass calculation, and using the European Medicines Agency’s weight standards for the animal categories. With these more refined calculations, authors Zahra Ardakani, Maurizio Aragrande, and Massino Canali aim to provide a more accurate estimate of global antimicrobial use (AMU) in cattle, chickens, and pigs. Understanding these patterns is crucial for addressing AMR and developing strategies for sustainable livestock management.
Key Findings
The study estimates that the global annual AMU for cattle, chickens, and pigs amounts to 76,060 tons of antimicrobial active ingredients. This is a significant revision from previous estimates due to a more detailed evaluation of animal weights and categories:
1. Cattle: 40,697 tons (53.5% of total AMU)
2. Pigs: 31,120 tons (40.9% of total AMU)
3. Chickens: 4,243 tons (5.6% of total AMU)
Figure 1: Distribution of global antimicrobial use among cattle, pigs, and chickens.
Methodology
The study utilizes the concept of Population Correction Units (PCU) to estimate antimicrobial usage, taking into account the weight and category of livestock at the time of treatment. This method differs from previous approaches that relied on live weight at slaughter, providing a more accurate representation of AMU.
The PCU is calculated by multiplying the number of animals by their average weight during treatment. This approach allows for differentiation by age and sex, which is particularly important for species like cattle and pigs.
Figure 2: (a) Changes in global PCU (million tonnes), (b) changes in global antibiotic use in mg per PCU, and (c) changes in global AMU (thousand tonnes) for cattle, chickens, and pigs; between 2010 and 2020. Abbreviations: PCU = Population Correction Unit; AMU = Antibiotic Use.
Figure 2: (a) Changes in global PCU (million tonnes), (b) changes in global antibiotic use in mg per PCU, and (c) changes in global AMU (thousand tonnes) for cattle, chickens, and pigs; between 2010 and 2020. Abbreviations: PCU = Population Correction Unit; AMU = Antibiotic Use.
Study shows lower AMU than previous estimates
The study highlights a significant shift in AMU patterns, with chickens showing a remarkable decrease in antimicrobial use despite increased production. This is indicative of improved management and more responsible use of antibiotics in the poultry industry.
The lower AMU in cattle and pigs, compared to previous estimates, underscores the importance of considering animal age and weight at treatment. These findings align closely with World Organization for Animal Health (WOAH) estimates, validating the methodology.
However, the study also acknowledges limitations, including reliance on European standards for average weight at treatment, which may not reflect global variations. Additionally, the lack of comprehensive global data on veterinary antibiotics presents challenges in creating fully accurate estimates.
Corrected estimate highlights improved production advances
This study provides a revised and potentially more accurate estimate of global antimicrobial use in livestock. By accounting for the weight and treatment categories of animals, it offers insights that could guide policy and management practices to mitigate the spread of antimicrobial resistance.
The article also indicates that the industry may have over-estimated antimicrobial usage in livestock and, just as importantly, that antimicrobial use has been kept in check or even reduced, despite increases in farmed animal headcounts. The lower usage is likely due to regulatory oversight and improvements in alternative methods to control and mitigate health challenges.
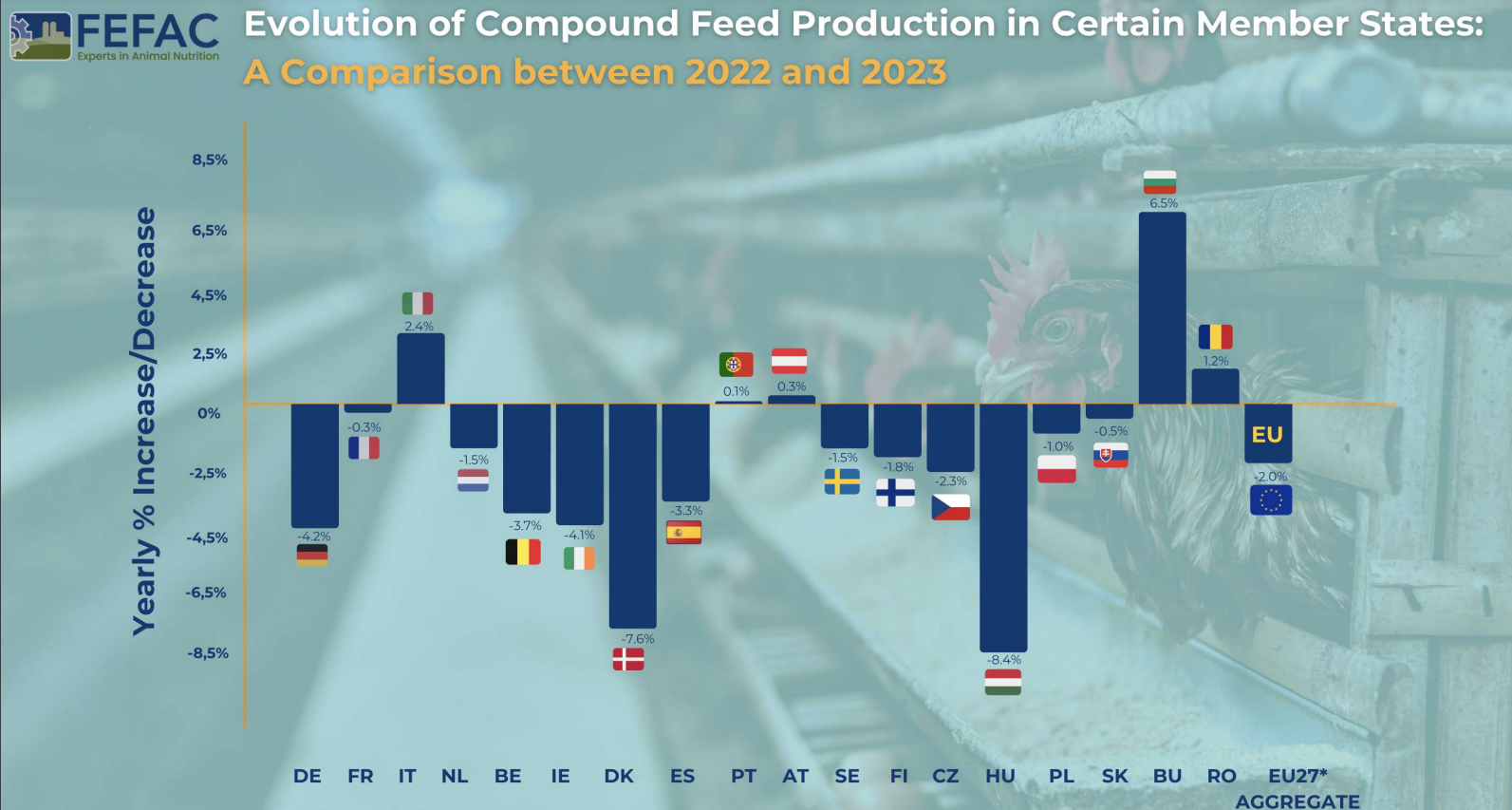
FEFAC: Quick Overview of 2023 EU Compound Feed Production
Total Production 2023: 144.3 million metric tons for farmed animals
Change from 2022: 2% decrease
Factors Influencing Decrease
Political and Market Pressures: Addressing crises and the shift towards sustainable feed.
Climate and Diseases: Effects of droughts, floods, Avian Influenza (AI), and African Swine Fever (ASF) on raw material supply and animal production.
National Policies: Initiatives for greenhouse gas and nitrate emission reduction.
Production Variability: Different trends across EU Member States, with notable decreases in countries like Germany, Ireland, Denmark, and Hungary, and slight increases in Austria, Bulgaria, Italy, and Romania.
Sector-Specific Trends
By Species
Pig Feed: Major decline of nearly 2.5 million tons. Key challenges included:
Loss of export markets, particularly in Asia
Negative media impact in Germany
Significant production drop in Denmark (-13.6%) and Spain (loss of 800,000 metric tons)
Italy’s ongoing struggle with ASF
Poultry Feed: Increase by 0.9 million tons, yet still 700,000 metric tons below 2021 levels. Challenges included declines in Hungary and Czechia due to reduced broiler production.
Cattle Feed: Decrease of 0.8 million tons from 2022.
“Green and animal welfare” policies affecting local production
Summary
The EU’s compound feed production in 2023 faced numerous challenges, leading to an overall decrease. The pig feed sector was most severely hit, while poultry feed showed some recovery. The influence of environmental, economic, and policy factors played a significant role in shaping these trends. Despite the price of feed cereals falling back to the levels seen before Russia’s invasion of Ukraine, these challenges will continue to be felt in 2024.
Ketosis: the most critical metabolic disease in dairy cows
Judith Schmidt, Product Manager On-Farm Solutions
Improvements in genetics, nutrition, and management continue to enhance dairy cows’ performance. However, being high-performance athletes comes at a cost, putting an extremely high burden on the animals’ energy metabolism. Especially around calving and during the first eight weeks of lactation, dairy cows can experience many stress factors: subclinical hypocalcemia, abomasum displacements, herd composition changes, or lameness. The more stress factors put the cows’ organism under pressure, the more likely they will become sick. A common consequence of stress is the occurrence of metabolic diseases, especially ketosis.
Both in terms of animal health and economic aspects, ketosis is probably the most critical dairy cow disease when also considering the correlated diseases. In this article, we explore the causes and consequences of ketosis and highlight prevention strategies that keep this issue under control.
Ketosis: causes and consequences
How ketosis develops
A restricted feed intake capacity and/or reduced energy concentration in the ration lead to a deficit in the animal’s energy balance. This situation occurs, for instance, at calving when the mother animal focuses her resources on the calf and its care. To compensate for the energy deficit, body fat is broken down for energy production. This process creates free fatty acids that accumulate in the liver and are partially converted into ketone bodies. These ketone bodies are a “transport medium” for energy, which various organs can use as an alternative energy source.
The problem arises when the deficiency lasts too long: more and more body fat is broken down, more and more fatty acids reach the liver, which leads to a fatty liver, and too high an amount of ketone bodies is formed and released into the blood. The ketone bodies in the blood inhibit appetite, resulting in less feed consumption and an energy deficit – the vicious cycle of ketosis begins.
Subclinical ketosis
Subclinical ketosis is defined as the stage of the disease at which an increased level of ketone bodies can be detected in the blood, urine, and milk. Furthermore, signs of hypoglycemia, increased levels of non-esterified fatty acid, and decreased hepatic gluconeogenesis can be seen in the blood. These conditions are typically not detected because there are no clinical signs.
Subclinical ketosis is a problem as it does not cause visible symptoms but leads to an increased incidence of subsequent diseases such as lab stomach displacement, clinical ketosis, and uterine inflammation. In addition, there may be loss of milk and fertility problems. Subclinically ill animals cannot be identified by the farmer by observation alone. Therefore, subclinical ketosis must be detected at an early stage to be able to act at the right time: prophylaxis instead of therapy.
There are several test possibilities to find out if an animal suffers from ketosis:
Milk: Milk test for ketosis detection has been available for many years. The results are to be obtained based on a color gamut. In contrast to blood analysis, the milk test does not evaluate exact values but shows a color change of the contained indicator. However, an increased milk cell content of the feeding of poorly fermented silages with a high butyric acid content significantly influences the result. The test often does not adequately reflect the actual conditions.
Urine: Another possibility is the examination of urine samples. Urine can be obtained spontaneously or with the help of a catheter. The results can also be read on a color scale of the urine test stripes. Like the milk test, the urine test only distinguishes different concentration ranges, but these are more finely graded than in the milk tests.
Blood: The most accurate but also most complex and expensive method is a blood test. It has the advantage that not only ketone bodies but also other parameters such as free fatty acids, minerals, and liver enzymes can be analyzed. In addition, the blood analysis results are evaluated in numbers and are more comparable than the color changes of test stripes. A good alternative is a rapid test by using a rapid test device, which is also used for measuring human blood sugar. A result is displayed with a drop of blood on a test strip within a few seconds.
Clinical ketosis
Depending on why there are elevated ketone body levels in the blood, we distinguish between primary and secondary clinical ketosis. For the primary form of clinical ketosis, the energy deficit itself (due to high performance and/or incorrect feeding) causes the condition. This form mainly occurs in susceptible, high-yielding dairy cows between the second and seventh weeks of lactation (Vicente et al., 2014). Secondary ketosis is caused indirectly by other diseases disease. A cow suffering from, for example, a claw disease might no longer consume a performance-based feed ration, leading to an energy deficit.
Typical symptoms
Typical of metabolic diseases, ketosis leads to a broad spectrum of symptoms. The classic symptoms at the beginning of the disease are a loss of appetite and decreased milk performance. As the disease develops, motor skills may be affected, and the excrement’s consistency becomes firmer and darker in color. The respiratory rate of sick animals increases, and they show dyspnea. Dyspnea is the medical description for breathing difficulties. Affected animals suffer from air shortage, which can occur in different situations. Due to the excretion of ketone bodies via the mucous membranes, the animals’ breath smells more or less strongly of acetone (Robinson and Williamson, 1977).
In addition, the animals undergo rapid and severe weight loss, and their general body conditions deteriorate noticeably. Furthermore, cows suffering from ketosis show increased milk fat content or an increased milk fat/protein quotient. Clinical symptoms include reduced general well-being, apathy, blindness, staggering, persistent “absent-minded” licking of the environment or overexcitability, muscle tremors, and aggressiveness (Andersson, 1984).
Effects on animal health and performance
Even in its subclinical form – if untreated – ketosis will engender health risks and reduced performance, negatively impacting milk yield and cows’ fertility. For clinical cases, typical effects include infertility, udder and hoof problems, and a fatty liver. Ketosis during early lactation is usually associated with fatty liver disease. In severe cases, the liver becomes enlarged and more fragile. It then no longer performs its detoxification function, toxic compounds increase, and the central nervous system is damaged. Anorexia or even a total loss of consciousness, the so-called hepatic coma, might ensue, ending in a complete liver function failure.
Direct economic costs range from high veterinary costs to the total loss of the dairy cow, i.e., approximately € 600 to € 1.000 per cow. Moreover, producers face indirect costs from secondary diseases such as fatty liver disease, increased postpartum behavior such as uterine infections, abomasum dislocations, or claw diseases.
Ketosis prevention: feeding and targeted supplementation
Feeding strategy
As part of the preparatory feeding, both dry and pregnant cows should receive rations that lead to an optimal (and not maximum) body condition at the time of calving. Animals with a poorer nutritional status do not have enough body fat reserves to compensate for lack of energy in the first phase of lactation. In more cases, animals have a too high BCS, leading to a risk of difficult births, and the cows have too little appetite at the beginning of lactation. These cows tend to show an excessive mobilization of fat reserves and develop a fatty liver. So prevention of ketosis of the current lactation starts with preventing a too-high BCS in the middle of the previous lactation.
The aim of feeding measures is to keep the lactating cow’s discrepancy between nutrient requirements and nutrient uptake as low as possible when the genetically determined performance potential is exhausted. For this reason, the ration must have a certain minimum energy density (high-quality forage and appropriate concentrate supplements). Also, anything that prevents the cows from ingesting the maximum amount of dry matter should be avoided.
Ket-o-Vital bolus for metabolic support
Another important preventive measure is the specific support of the calving cow’s liver, rumen, and immune system. EW Nutrition’s Ket-o-Vital Bolus was explicitly designed to reduce the risk of ketosis. It contains fast-available glucogenic substances, positively influencing the cow’s energy metabolism. Another advantage the bolus offers is the slow release of the contained cobalt, selenium, niacin, and active yeast:
Cobalt is a trace element important to form cobalamin, the so-called vitamin B12. It is essential for blood formation and the functioning of the nervous system.
Selenium protects cells from oxidative damage and ensures an intact immune defense;
Niacin is a B vitamin that intervenes in energy metabolism and prevents fatty liver syndrome;
And active yeast supports rumen health, preventing rumen acidosis and increasing feed intake.
The application of the Ket-o-Vital Bolus is profitable and straightforward. Only one bolus per application is required.
Ketosis control: be one step ahead
High-performance dairy cows are at risk of ketosis, which results in involuntary culling, poor health, and performance losses. Advanced feed management practices combined with the targeted use of the Ket-o-Vital bolus offer a solution for preventing this debilitating disease. The bolus protects the cows from clinical and subclinical ketosis, reduces metabolic disorders, increases appetite, and improves health – leading to a quick recovery and ensuring profitable production.
References
Vicente, Fernando, María Luisa Rodríguez, Adela Martínez-Fernández, Ana Soldado, Alejandro Argamentería, Mario Peláez, and Begoña de la Roza-Delgado. “Subclinical ketosis on dairy cows in transition period in farms with contrasting butyric acid contents in silages.” The Scientific World Journal 2014 (November 25, 2014): 1–4. https://doi.org/10.1155/2014/279614.
Andersson, L. “Concentrations of blood and milk ketone bodies, blood isopropanol and plasma glucose in dairy cows in relation to the degree of hyperketonaemia and clinical signs*.” Zentralblatt für Veterinärmedizin Reihe A 31, no. 1-10 (1984): 683–93. https://doi.org/10.1111/j.1439-0442.1984.tb01327.x.
Robinson, A. M., and D. H. Williamson. “Effects of acetoacetate administration on glucose metabolism in mammary gland of fed lactating rats.” Biochemical Journal 164, no. 3 (1977): 749–52. https://doi.org/10.1042/bj1640749.
Masked mycotoxins – particularly dangerous for dairy cows
By Si-Trung Tran, SEAP Regional Technical Manager, EW Nutrition
Technical Team and Dr. Inge Heinzl, Editor, EW Nutrition
Mycotoxins are secondary metabolites of fungi, commonly found as contaminants in agricultural products. In some cases, these compounds are used in medicine or industry, such as penicillin and patulin. In most cases, however, they are considered xenobiotics that are toxic to animals and humans, causing the disease collectively known as mycotoxicosis. The adverse effects of mycotoxins on human and animal health have been documented in many publications. Aflatoxins (AFs) and deoxynivalenol (DON, vomitoxin) are amongst the most critical mycotoxins affecting milk production and -quality.
Aflatoxins do not only affect cows
Aflatoxins (AFs) are highly oxygenated, heterocyclic difuranocoumarin compounds produced by Aspergillus flavus and Aspergillus parasiticus. They colonize crops, including many staple foods and feed ingredients. Within a group of over 20 AFs and derivatives, aflatoxin B1 (AFB1), B2, G1, and G2 are the most important naturally occurring compounds.
Among the aflatoxins, AFB1 is the most widespread and most toxic to humans and animals. Concern about mycotoxin contamination in dairy products began in the 1960s with the first reported cases of contamination by aflatoxin M1 (AFM1), a metabolite of AFB1 formed in the liver of animals and excreted in the milk.
There is ample evidence that lactating cows exhibit a significant reduction in feed efficiency and milk yield within a few days of consuming aflatoxin-contaminated feed. At the cellular level, aflatoxins cause degranulation of endoplasmic membranes, loss of ribosomes from the endoplasmic reticulum, loss of nuclear chromatin material, and altered nuclear shapes. The liver, as the organ mainly dealing with the decontamination of the organism, gets damaged, and performance drops. Immune cells are also affected, reducing immune competence and vaccination success (Arnold and Gaskill, 2023).
DON reduces cows’ performance
Another mycotoxin that can also reduce milk quality and affect metabolic parameters, as well as the immune function of dairy cows, is DON. DON is produced by different fungi of the Fusarium genus that infect plants. DON synthesis is associated with rainy weather from crop flowering to harvest. Whitlow and co-workers (1994) reported the association between DON and poor performance in dairy herds and showed decreased milk production in dairy cows fed 2.5 mg DON/kg. However, in cows fed 6 to 12 mg DON/kg dry matter for 10 weeks, no DON or its metabolite DOM-1 residues were detected in milk.
Masked mycotoxins hide themselves during analysis
Plants suffering from fungal infestations and thus confronted with mycotoxins convert the harmful forms of mycotoxins into less harmful or harmless ones for themselves by conjugation to sulfates, organic acids, or sugars. Conjugated mycotoxins cannot always be detected by standard analytical methods. However, in animals, these forms can be released and transformed into parent compounds by enzymes and microorganisms in the gastrointestinal tract. Thus, the feed may show a concentration of mycotoxins that is still below the limit value, but in the animal, this concentration is suddenly much higher. In dairy cows, the release of free mycotoxins from conjugates during digestion may play an important role in understanding the silent effects of mycotoxins.
Fusarium toxins, in particular, frequently occur in this “masked form”. They represent a serious health risk for animals and humans.
Aflatoxins first show up in the milk
Masked aflatoxins may also play a role in total aflatoxin contamination of feed materials. Research has harvested little information on masked aflatoxins that may be present in TMR ingredients. So far, metabolites such as Aflatoxin M2 have been identified (Righetti, 2021), which may reappear later in milk as AFM1.
DON-related symptoms without DON?
Sometimes, animals show DON-related symptoms, with low levels detected in the feed or raw materials. Besides sampling errors, this enigma could be due to conjugated or masked DON, which is structurally altered DON bound to various compounds such as glucose, fatty acids, and amino acids. These compounds escape conventional feed analysis techniques because of their modified chemical properties but can be released as their toxic precursors after acid hydrolysis.
Masked DON was first described in 1984 by Young and co-workers, who found that the DON content of yeast-fermented foods was higher than that of the contaminated wheat flour used in their production. The most plausible reason for this apparent increase was that the toxin from the wheat had been converted to a compound other than DON, which could be converted back to DON under certain conditions. Since this report, there has been much interest in conjugated or masked DON.
Silage: masked DON is a challenge for dairy producers
Silage is an essential feed for dairy cows, supporting milk production. Most silage is made from corn and other grains. The whole green plant is used, which can be infected by fungi. Since infection of corn with Fusarium spp. and subsequent DON contamination is usually a major problem in the field worldwide, a relatively high occurrence of this toxin in silage must be expected. The ensiling process may reduce the amount of Fusarium fungi, but the DON formed before ensiling is very stable.
Silage samples show DON levels of concern
It is reasonable to assume that the DON biosynthesized by the fungi was metabolized by the plants to a new compound and thus masked DON. Under ensiling conditions, masked DON can be hydrolyzed, producing free DON again. Therefore, the level of free DON in the silage may not reflect the concentration measured in the plants before ensiling.
A study analyzed 50 silage samples from different farms in Ontario, Canada. Free DON was found in all samples, with levels ranging from 0.38 to 1.72 µg/g silage (unpublished data). Eighty-six percent of the samples contained DON at concentrations higher than 0.5 µg/g. Together with masked DON, it poses a potential threat to dairy cattle.
Specific hydrolysis conditions allow detection
However, in the natural ensiling process, the conditions for hydrolysis of masked DON are not optimal. The conditions that allow improved analysis of masked DON were recently described. This method detected masked DON in 32 of 50 silage samples (64%) along with free DON, increasing DON concentration by 23% in some cases (unpublished data).
Mycotoxins impact humans and animals
Aflatoxins, as well as DON, have adverse effects. In the case of DON, the impact on the animal is significant; in the case of aflatoxin, the possible long-term effects on humans are of higher relevance.
DON has more adverse effects on the animal and its performance
Unlike AFs, DON may be found in milk at low or trace concentrations. It is more associated with negative effects in the animal, altered rumen fermentation, and reduced flow of usable protein into the duodenum. For example, milk fat content was significantly reduced when cows were fed 6 µg DON/kg. However, the presence of DON also indicates that the feed probably contains other mycotoxins, such as zearalenone (ZEA) (estrogenic mycotoxin) and fusaric acid (pharmacologically active compound). All these mycotoxins may interact to cause symptoms that are different or more severe than expected, considering their individual effects. DON and related compounds also have immunosuppressive effects, resulting in increased somatic cell counts in milk. The U.S. FDA has established an action level for DON in wheat and wheat-derived products intended for cows, which is 5µg DON/g feed and the contaminated ingredient must not exceed 40% of the ration.
Aflatoxins decrease milk quality and pose a risk to humans
Aflatoxins are poorly degraded in the rumen, with aflatoxicol being the main metabolite that can be reconverted to AFB1. Most AFs are absorbed and extensively metabolized/hydrolyzed by enzymes found mainly in the liver. This results in the formation of AFM1, a part of which is conjugated to glucuronic acid and subsequently excreted in the bile. The other part enters the systemic circulation. It is either excreted in urine or milk. AFM1 appears within 12-48 hours after ingestion in cow’s milk. The excreted amount of AFM1 in milk from dairy cows usually ranges from 0.17% to 3% of the ingested AFB1. However, this carryover rate may vary from day to day and from one milking to the next in individual animals, as it is influenced by various factors, such as feeding regime, health status, individual biotransformation capacity, and, of course, by actual milk production. Carryover rates of up to 6.2% have been reported in high-yielding dairy cows producing up to 40 liters of milk per day.
In various experiments, AFM1 showed both carcinogenic and immunosuppressive effects. Accordingly, the International Agency for Research on Cancer (IARC) classified AFM1 as being in Group 2B and, thus, possibly carcinogenic in humans. The action level of 0.50 ppb and 0.05 ppb for AFM1 in milk is strictly adhered to by the U.S. Food and Drug Administration (FDA) and the European Food Safety Authority (EFSA), respectively.
Trials show the high adsorption capacity of Solis Max
A trial was conducted at an independent laboratory located in Spain. The evaluation of the performance of Solis Max was executed with the following inclusion levels:
0.10% equivalent to 1.0 kg of Solis Max per ton of feed
0.20% equivalent to 2.0 kg of Solis Max per ton of feed
A phosphate buffer solution at pH 7 was prepared for the trial to simulate rumen conditions. Each mycotoxin was tested separately, preparing solutions with known contamination (final concentration described in the table below). The contaminated solutions were divided into 3 parts: A positive control, 0.10% Solis Max and 0.20% Solis Max. All samples were incubated at 41°C for 1 hour, centrifuged, and the supernatant was analyzed for the mycotoxin added to determine the binding efficacy. All analyses were carried out by high-performance liquid chromatography (HPLC) with standard detectors.
Mycotoxin
Contamination Level (ppb)
Aflatoxin B1
800
DON
800
Fumonisin B1
2000
ZEA
1200
Results: The higher concentration of Solis max showed a higher adsorption rate for most mycotoxins. The high dose of Solis Max adsorbed 99% of the AFB1 contamination. In the case of DON, more than 70% was bound. For fumonisin B1 and zearalenone, Solis max showed excellent binding rates of 87.7% and 78.9%, respectively (Figure 1).
Figure 1: Solis Max showed a high binding capacity for the most relevant mycotoxins
Another trial was conducted at an independent laboratory serving the food and feed industry and located in Valladolid, Spain.
All tests were carried out as duplicates and using a standard liquid chromatography/mass spectrometry (LC/MS/MS) quantification. Interpretation and data analysis were carried out with the corresponding software. The used pH was 3.0, toxin concentrations and anti-mycotoxin agent application rates were set as follows (Table 1):
Table 1: Trial set-up testing the binding capacity of Solis Plus 2.0 for several mycotoxins in different contamination levels
Results:
Under acidic conditions (pH3), Solis Plus 2.0 effectively adsorbs the three tested mycotoxins at low and high levels. 100% binding of aflatoxin was achieved at a level of 150ppb and 98% at 1500ppb.In the case of fumonisin, 87% adsorption could be reached at 500ppb and 86 for a challenge with 5000ppb. 43% ochratoxin was adsorbed at the contamination level of 150ppb and 52% at 1500ppb.
Figure 2: The adsorption capacity of Solis Plus 2.0 for three different mycotoxins at two challenge levels
Mycotoxins – Effective risk management is of paramount importance
Although the rumen microflora may be responsible for conferring some mycotoxin resistance to ruminants compared to monogastric animals, there are still effects of mycotoxins on rumen fermentation and milk quality. In addition, masked mycotoxins in feed present an additional challenge for dairy farms because they are not readily detectable by standard analyses.
Feeding dairy cows with feed contaminated with mycotoxins can lead to a reduction in milk production. Milk quality may also deteriorate due to an adverse change in milk composition and mycotoxin residues, threatening the innocuousness of dairy products. Dairy farmers should therefore have feed tested regularly, consider masked mycotoxins, and take action. EW Nutrition’s MasterRisk tool provides a risk evaluation and corresponding recommendations for the use of products that mitigate the effects of mycotoxin contamination and, in the end, guarantee the safety of all of us.



 .
.