Enhancing Poultry Gut Health with Novel Xylanase: A Sustainable Path to Reduced Antimicrobial Use
By Ajay Bhoyar, Senior Global Technical Manager, EW Nutrition
Gut health is pivotal to profitable poultry production, as the gastrointestinal tract (GIT) enables nutrient digestion and absorption while acting as a defense against pathogens. A healthy gut improves feed conversion, boosts immune resilience, and reduces reliance on antimicrobials—critical in the fight against antimicrobial resistance (AMR). With AMR posing significant threats to public health and animal agriculture, strategies like biosecurity, sustainable management, and effective dietary interventions are gaining traction. Feed enzymes have emerged as essential tools for managing feed costs, mitigating anti-nutritional factors, and improving nutrient utilization. Among these, feed enzymes like xylanase stand out. By breaking down xylan, a major component of non-starch polysaccharides (NSPs) in plant-based feed ingredients, xylanase reduces gut viscosity, enhances nutrient utilization, and supports optimal gut health and productivity. This article explores the innovative application of novel GH10 xylanases, such as Axxess XY, as a sustainable solution for improving feed efficiency and gut health in poultry production.
Xylanase in Poultry Nutrition
Xylanase plays a pivotal role in enhancing nutrient availability by addressing the limitations of endogenous enzyme synthesis in poultry. Xylanase enzymes belong to the carbohydrase class, catalyzing the breakdown of xylan, a major NSP in plant-based feed ingredients. They hydrolyze xylan into simple sugars like arabino-xylo-oligosaccharides (AXOs) and xylo-oligosaccharides (XOs), reducing the encapsulation of nutrients and digesta viscosity. These actions improve overall nutrient digestibility and bird performance.
Fig.1: Arabinoxylans – anti-nutrient mode of action in chicken
The primary benefit of feed xylanase lies in its ability to reduce digesta viscosity. By partially hydrolyzing NSPs in the upper digestive tract, xylanase ensures better nutrient absorption in the small intestine. Studies (Matthiesen et al., 2021; Choct & Annison, 1992) confirm that reduced viscosity enhances feed digestibility, leading to improved performance in poultry. Further, to realize the optimum benefits, it is crucial that xylanase efficiently degrades both soluble and insoluble arabinoxylans. The insoluble arabinoxylans are part of the cell wall structure of plant cells, resulting in a cage effect, entrapping nutrients like starch and protein. Effectively breaking down insoluble arabinoxylans ensures that the nutrients trapped in plant cell walls are released for growth and production.
Mechanisms Supporting Gut Health
Viscosity Reduction
High NSP content increases digesta viscosity and slows digestion and nutrient absorption. Soluble arabinoxylan is not digested in the small intestine of broilers. It produces a viscous chime, leading to the proliferation of pathogenic bacteria, intestinal inflammation, impairment of barrier function in the intestine, and severe intestinal lesions (Teirlynck et al., 2009). Xylanase mitigates this by breaking down xylans, a major component of NSPs in common feed ingredients. This results in a better flow of digesta and reduced energy losses.
Microbial Metabolites
Xylo-oligosaccharides (XOS) can also be produced in the intestine of monogastric animals to some extent when exogenous enzymes, such as xylanase, are added to the feed (Baker et al., 2021).
The XOS generated by xylanase action on arabinoxylans can act as prebiotics, fostering beneficial bacteria like Lactobacillus and Bifidobacterium, which can outcompete harmful species. XOS can positively impact the gut microbiota, enhance short-chain fatty acid (SCFA) production, stimulate immune activity in the gastrointestinal tract, and improve energy utilization.
Fig. 2. Axxess XY improved beneficial microbes and reduced the clostridial population in broilers.
Barrier Function
By lowering inflammation and irritation in the intestine, xylanase helps maintain gut integrity, reducing the risk of pathogen translocation from the intestinal lumen. In a broiler study, xylanase decreases epithelial apoptosis index, up-regulates tight junction gene expression, and inhibits mucin synthesis in the small intestine, likewise alleviating the intestinal mucosal barrier impairment from Clostridium perfringens challenge (Liu et al., 2012).
Practical Considerations for Xylanase Use
Enzyme Stability
Enzymes are proteins that tend to lose their catalytic activity at high temperatures. When exposed to excessive heat, an enzyme’s protein structure can irreversibly unfold, disrupting its active site and causing loss of function. Therefore, ensuring enzyme stability during feed processing is critical for maintaining its activity in the intestine. Intrinsically heat-stable enzymes have an inherent ability to withstand higher temperatures without the need for a protective coating and are immediately available for action upon ingestion.
Feed Composition
Xylanase efficacy is influenced by diet composition, particularly the NSP content and the presence of xylanase inhibitors in common feedstuffs. It is important to choose a xylanase that can resist the activity of xylanase inhibitors and is effective against both soluble and insoluble arabinoxylans.
The recommended energy matrix value for the xylanase enzyme should be used while formulating the feeds to create energy-deficient diets to reap the full benefits of xylanase use.
Optimal Dosage
Proper dosing is essential to maximizing the benefits of feed enzymes while avoiding unnecessary costs. It is important to follow manufacturers’ recommendations and avoid underdosing an enzyme.
GH10 Xylanases: The Superior Choice for Animal Nutrition
Most feed xylanases are classified into glycoside hydrolase families 10 (GH10) and 11 (GH11) based on their substrate specificity, catalytic action, and structural features.
Why GH10 Xylanases Are More Effective
Broader Substrate Specificity:
Unlike GH11 xylanases, GH10 xylanases can effectively hydrolyze both soluble and insoluble xylan substrates. This broader activity ensures an efficient breakdown of xylans in a wide range of feed ingredients.
Higher Catalytic Efficiency:
GH10 enzymes cleave xylan at substituted regions, yielding shorter xylo-oligosaccharides that can positively impact gut health and maximize nutrient availability.
Thermostability:
Feed processing often involves high temperatures during pelleting. Axxess XY, a GH10 family xylanase, demonstrates remarkable thermostability, maintaining over 85% activity even at 95°C for extended conditioning times. This resilience ensures consistent enzyme performance during feed manufacturing and digestion.
Fig.3: Optimum recovery of Axxess XY at elevated conditioning time and temperatures
Novel Applications of Axxess XY: A GH10 Xylanase
Axxess XY exemplifies the advantages of GH10 xylanases in poultry nutrition. Its ability to efficiently act on both soluble and insoluble arabinoxylans makes it a versatile feed enzyme. The enzyme’s high thermostability ensures efficient enzyme activity in the gut and subsequent optimum nutrient utilization under challenging processing conditions, promoting gut health and maximizing performance.
Key Benefits of Axxess XY
Enhanced Nutrient Utilization:
By unlocking nutrients trapped in NSPs, Axxess XY promotes better feed conversion ratios (FCRs).
Improved Gut Health:
Reducing the digest’s viscosity reduces gut health challenges and predisposition to gut infections. Further, the short-chain oligosaccharides released by Axxess XY support beneficial gut microbiota, improving digestive health.
Economic Efficiency:
Enabling the optimum use of high-fiber, cost-effective, locally available feed ingredients without compromising performance makes Axxess XY an asset for profitability.
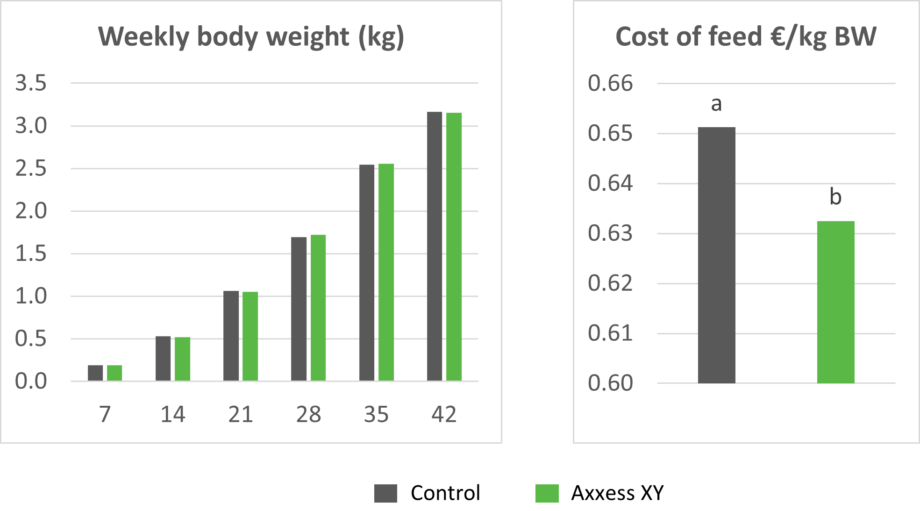
In a recently conducted 42-day trial at a commercial farm, Axxess XY maintained the average body weight of broilers with a 100 kcal/kg reduction in metabolizable energy while significantly reducing feed cost/kg body weight. The diets were based on corn, DDGS, and soybean meal.
Figures 4 and 5: Body weight and cost of feed in broilers fed a diet reduced by 100 kcal/kg in metabolizable energy compared to a standard diet without Axxess XY
Conclusion
Xylanase exemplifies how feed enzymes can transcend their traditional role in feed cost reduction to support enhanced gut health. Xylanase supports reduced antimicrobial use in poultry production by improving nutrient utilization, reducing digesta viscosity, and fostering healthy microbiota. Its integration into comprehensive gut health management strategies offers a sustainable pathway to combat AMR and ensure the long-term viability of poultry farming. By targeting NSPs, these enzymes enhance nutrient digestibility, reduce feed costs, and support sustainable production practices.
GH10 xylanases, particularly Axxess XY, stand out for their superior substrate specificity, catalytic efficiency, and thermostability. By incorporating Axxess XY into feed formulations, poultry producers can unlock the full nutritional potential of feed ingredients, ensuring optimal performance and profitability. As the poultry industry continues to evolve, adopting advanced enzyme technologies like Axxess XY represents a strategic step toward sustainable and efficient animal nutrition.
References:
Baker, J.T.; Duarte, M.E.; Holanda, D.M.; Kim, S.W. Friend or Foe? Impacts of Dietary Xylans, Xylooligosaccharides, and Xylanases on Intestinal Health and Growth Performance of Monogastric Animals. Animals 2021, 11, 609.
Choct, M., and G. Annison. “Anti‐nutritive Effect of Wheat Pentosans in Broiler Chickens: Roles of Viscosity and Gut Microflora.” British Poultry Science 33, no. 4 (September 1992): 821–34. https://doi.org/10.1080/00071669208417524.
Liu D, Guo S, Guo Y. Xylanase supplementation to a wheat-based diet alleviated the intestinal mucosal barrier impairment of broiler chickens challenged by Clostridium perfringens. Avian Pathol. 2012;41(3):291-8.
Matthiesen, Connie F., Dan Pettersson, Adam Smith, Ninfa R. Pedersen, and Adam. C. Storm. “Exogenous Xylanase Improves Broiler Production Efficiency by Increasing Proximal Small Intestine Digestion of Crude Protein and Starch in Wheat-Based Diets of Various Viscosities.” Animal Feed Science and Technology 272 (February 2021): 114739. https://doi.org/10.1016/j.anifeedsci.2020.114739.
Teirlynck, E.; Haesebrouck, F.; Pasmans, F.; Dewulf, J.; Ducatelle, R.; van Immerseel, F. The cereal type in feed influences Salmonella enteritidis colonization in broilers. Poult. Sci. 2009, 88, 2108–2112.
Sustainability will push more by-products into pig feed – Keep track of mycotoxins!
Mycotoxin Team EW Nutrition
Most grains used in feed are susceptible to mycotoxin contamination, causing severe economic losses all along feed value chains. As skyrocketing raw material prices force producers to include a higher proportion of economical cereal by-products in the feed, the risks of mycotoxin contamination likely increase. This article reviews why mycotoxins cause the damage they do – and how effective toxin-mitigating solutions prevent this damage.
Mycotoxin contamination of cereal by-products requires solutions
Cereal by-products may become more important feed ingredients as grain prices increase. However, from a sustainability point of view and considering population growth, using cereal by-products in animal feed makes much sense. Distiller’s dried grains with solubles (DDGS) are a good example of how by-products from food processing industries can become high-quality animal feed.
Figure 1: By-products are a crucial protein source (data from FEFAC Feed&Food 2021 report)
Still, research on what happens to mycotoxins during food processing shows that mycotoxins are concentrated into fractions that are commonly used as animal feed (cf. Pinotti et al., 2016; Caballero and Heinzl, 2022). To safeguard animal health and performance when feeding lower-quality cereals, monitoring mycotoxin risks through regular testing and using toxin-mitigating solutions is essentialy.
Problematic effects of mycotoxins on the intestinal epithelium
Mostmycotoxinsare absorbedin the proximal partof the gastrointestinal tract. Thisabsorption can behigh, as in the case of aflatoxins(ca. 90%), but also very limited, as in the case of fumonisins(< 1%); moreover, it depends on the species.Notably, a significant portion ofunabsorbedtoxinsremains within thelumenof the gastrointestinal tract.
Importantly, studies based on realistic mycotoxin challenges (e.g., Burel et al., 2013) show that the mycotoxin levels necessary to trigger damaging processes are lower than the levels reported as safe by EFSA, the Food Safety Agency of the European Union. The ultimate consequences range from diminished nutrient absorption to inflammatory responses and pathogenic disorders in the animal (Figure 2).
Figure 2: Mycotoxins’ impact on the GIT and consequences for monogastric animals
1. Alteration of the intestinal barrier‘s morphology and functionality
Several studies indicate that mycotoxins such as aflatoxin B1, DON, fumonisin B1, ochratoxin A, and T2, can increase the permeability of the intestinal epithelium of poultry and swine (e.g., Pinton & Oswald, 2014). This is primarily a consequence of the inhibition of protein synthesis.
As a result, there is an increase in the passage of antigens into the bloodstream (e.g., bacteria, viruses, and toxins). This increases the animal’s susceptibility to infectious enteric diseases. Moreover, the damage that mycotoxins cause to the intestinal barrier entails that they are also being absorbed at a higher rate.
2. Impaired immune function in the intestine
The intestine is a very active immune site, where several immuno-regulatory mechanisms simultaneously defend the body from harmful agents. Immune cells are affected by mycotoxins through the initiation of apoptosis, the inhibition or stimulation of cytokines, and the induction of oxidative stress.
It is known that fumonisin B1 may induce changes in the balance of sphingolipids at the cellular level, including for gastrointestinal cells. This facilitates the adhesion of pathogenic bacteria, increases in their populations, and prolongs infections, as has been shown in the case of E. coli. The colonization of the intestine of food-producing animals by pathogenic strains of E. coli and Salmonella also poses a risk to human health.
4. Interaction with bacterial toxins
When mycotoxins induce changes in the intestinal microbiota, this can increase the endotoxin concentration in the intestinal lumen. Endotoxins promote the release of several cytokines that induce an enhanced immune response, causing inflammation, thus reducing feed consumption and animal performance, damage to vital organs, sepsis, and death of the animals in some cases.
The synergy between mycotoxins and endotoxins can result in an overstimulation of the immune system. The interaction between endotoxins and estrogenic agents such as zearalenone, for example, generates chronic inflammation and autoimmune disorders because immune cells have estrogen receptors, which are stimulated by the mycotoxin.
Increased mycotoxin risks through by-products? Invest in mitigation solutions
To prevent the detrimental consequences of mycotoxins on animal health and performance, proactive solutions are needed that support the intestinal epithelium’s digestive and immune functionality and help maintain a balanced microbiome in the GIT. This becomes even more important as the current market conditions will likely engender a long-term shift towards including more cereal by-products in animal diets.
Trial data shows that EW Nutrition’s toxin-mitigating solution SOLIS MAX 2.0 provides adequate protection against feedborne mycotoxins. The synergistic combination of ingredients in SOLIS MAX 2.0 prevents mycotoxins from damaging the animals’ gastrointestinal tract and entering the bloodstream and additionally acts as antioxidant and liver-protecting:
Figure 3: Moa of Solis Max 2.0
In-vitro study shows strong mitigation effects of SOLIS MAX 2.0 against a wide range of mycotoxins
Animal feed is often contaminated with two or more mycotoxins, making it essential for an anti-mycotoxin agent to be effective against a wide range of different mycotoxins. A trial with SOLIS MAX 2.0 was conducted at an independent laboratory in Spain with an inclusion level of the product of 0.10% (equivalent to 1 kg per ton of feed). A phosphate buffer solution at pH 7 was prepared to simulate intestinal conditions in which a portion of the mycotoxins may be released from the binder (desorption). The following mycotoxins were evaluated in the test (see Table 1):
Table 1: Mycotoxin challenges
Each mycotoxin was tested separately by adding a challenge to buffer solutions, incubating for one hour at 41°C, to establish the baseline (table). At the same time, a solution with the toxin challenge and Solis Max 2.0 was prepared, incubated, and analyzed for the residual mycotoxin to find the binding efficacy. All analyses were carried out using high-performance liquid chromatography (HPLC) with standard detectors.
Figure 4: SOLIS MAX 2.0 (1 kg/t of feed) adsorption capacity against different mycotoxins (%)
The results (Figure 4) demonstrate that SOLIS MAX 2.0 is a highly effective solution against the most common mycotoxins in raw materials and animal feed.
Mycotoxin risk management for better animal feed
A healthy gastrointestinal tract is crucial to animals’ overall health: it ensures that nutrients are optimally absorbed, provides adequate protection against pathogens through its immune function, and is key to maintaining a well-balanced microflora. Even at levels considered safe by the European Union, mycotoxins can compromise different intestinal functions, resulting in lower productivity and susceptibility to disease.
The globalized feed trade, which spreads mycotoxins beyond their geographical origin, climate change, and raw material market pressures additionally escalate the problem. On top of rigorous testing, producers should mitigate unavoidable mycotoxin exposures by using solutions such as SOLIS MAX 2.0 – for stronger animal health, welfare, and productivity.
References
Antonissen, Gunther, An Martel, Frank Pasmans, Richard Ducatelle, Elin Verbrugghe, Virginie Vandenbroucke, Shaoji Li, Freddy Haesebrouck, Filip Van Immerseel, and Siska Croubels. “The Impact of Fusarium Mycotoxins on Human and Animal Host Susceptibility to Infectious Diseases.” Toxins 6, no. 2 (January 28, 2014): 430–52. https://doi.org/10.3390/toxins6020430.
Burel, Christine, Mael Tanguy, Philippe Guerre, Eric Boilletot, Roland Cariolet, Marilyne Queguiner, Gilbert Postollec, et al. “Effect of Low Dose of Fumonisins on Pig Health: Immune Status, Intestinal Microbiota and Sensitivity to Salmonella.” Toxins 5, no. 4 (April 23, 2013): 841–64. https://doi.org/10.3390/toxins5040841.
Burton, Emily J., Dawn V. Scholey, and Peter E. Williams. “Use of Cereal Crops for Food and Fuel – Characterization of a Novel Bioethanol Coproduct for Use in Meat Poultry Diets.” Food and Energy Security 2, no. 3 (September 19, 2013): 197–206. https://doi.org/10.1002/fes3.30.
Ghareeb, Khaled, Wageha A. Awad, Josef Böhm, and Qendrim Zebeli. “Impacts of the Feed Contaminant Deoxynivalenol on the Intestine of Monogastric Animals: Poultry and Swine.” Journal of Applied Toxicology 35, no. 4 (October 28, 2014): 327–37. https://doi.org/10.1002/jat.3083.
Mani, V., T. E. Weber, L. H. Baumgard, and N. K. Gabler. “Growth and Development Symposium: Endotoxin, Inflammation, and Intestinal Function in livestock1,2.” Journal of Animal Science 90, no. 5 (May 1, 2012): 1452–65. https://doi.org/10.2527/jas.2011-4627.
Obremski, K. “The Effect of in Vivo Exposure to Zearalenone on Cytokine Secretion by Th1 and Th2 Lymphocytes in Porcine Peyer’s Patches after in Vitro Stimulation with LPS.” Polish Journal of Veterinary Sciences 17, no. 4 (2014): 625–32. https://doi.org/10.2478/pjvs-2014-0093.
Oswald, I. P., C. Desautels, J. Laffitte, S. Fournout, S. Y. Peres, M. Odin, P. Le Bars, J. Le Bars, and J. M. Fairbrother. “Mycotoxin Fumonisin B1 Increases Intestinal Colonization by Pathogenic Escherichia Coli in Pigs.” Applied and Environmental Microbiology 69, no. 10 (2003): 5870–74. https://doi.org/10.1128/aem.69.10.5870-5874.2003.
Pinotti, Luciano, Matteo Ottoboni, Carlotta Giromini, Vittorio Dell’Orto, and Federica Cheli. “Mycotoxin Contamination in the EU Feed Supply Chain: A Focus on Cereal Byproducts.” Toxins 8, no. 2 (February 15, 2016): 45. https://doi.org/10.3390/toxins8020045.
Pinton, Philippe, and Isabelle Oswald. “Effect of Deoxynivalenol and Other Type B Trichothecenes on the Intestine: A Review.” Toxins 6, no. 5 (May 21, 2014): 1615–43. https://doi.org/10.3390/toxins6051615.
Mycotoxins pose a threat to the horse’s digestive system
Author: Judith Schmidt, Product Manager On Farm Solutions
Alarm in the gut! Horses have a susceptible digestive system that can quickly become unbalanced. Intestinal disorders in horses are usually associated with colic. Many factors can be responsible for intestinal issues. Have you ever thought about mycotoxins? What can horse owners do to support their horse´s gut health?
The equine stomach is not robust at all. Depending on their age and use, more than half of all horses suffer from stomach pain. Their digestive system is very sensitive and very different from that of other mammals: Horses cannot vomit and often suffer from severe abdominal pain, diarrhea, or cramps if they overeat or ingest spoiled feed.
The horse´s digestive system is complex and sensitive
The horse´s stomach has a relatively small capacity of around twelve to fifteen liters. Depending on the feed’s consistency and composition, it remains in the stomach for around one to five hours before it is pressed through the stomach outlet (pylorus) into the small intestine. The horse´s entire intestine is about ten times its body length.
Figure 1: The horse’s digestive tract
The horse´s gastrointestinal tract is a complex network, reacting extremely sensitively to changes and, therefore, highly susceptible to disorders. It essentially consists of the head intestine (lips, oral cavity, teeth, and esophagus), stomach (blind pouch, fundus, and stomach outlet), small intestine (duodenum, jejunum, and ileum), and large intestine (caecum, colon and rectum). Each section plays a crucial role in the digestive process; any disruption can lead to health issues. Understanding this structure is key to maintaining a horse’s digestive health.
Digestive disorders can have various reasons
Intestinal problems in horses can stem from diverse causes, often a complex interplay of multiple factors. By understanding these causes more deeply, horse owners can be better equipped to prevent and manage these issues. In the following, we delve into several of these causes.
1. Too long time between the feedings
Usually, a feeding break should be at most four to six hours, as, in nature, a horse is busy eating for at least 18 hours a day. In contrast to humans, who produce stomach acid only after food intake, the horse’s stomach produces gastric acid around the clock. The continuous intake of roughage, intensive chewing, and high saliva production (a horse produces 5 to 10 L of saliva per day) is, therefore, essential to protect the stomach mucosa by neutralizing excess gastric acid.
A too-long time between feedings and, therefore, no saliva production leads to an accumulation of gastric acid in the stomach. Four hours without roughage can already cause inflammation of the mucosa and probably ulcers.
2. Excessive amounts of concentrated feed
Excessive amounts of concentrates such as wheat or rye, conditioned by less chewing activity, increase gastric acid and histamine production, and the stomach lining can be attacked. Also, in this case, the development of stomach ulcers is possible.
Furthermore, the possibly resulting hyperacidity of the organism can lead to malfunctions of the organs, the skin, and the hooves.
3. Stress
Stress can also lead to a higher production of gastric acid and, therefore, to gastric ulcers. The horse is a flight animal. When it is under stress, it prepares for the impending escape, and the muscles are preferably supplied with blood, resulting in a lower blood flow to the mucous membranes. Furthermore, the rising cortisone level reduces the hydrochloric acid-suppressing prostaglandin E. As a result, more stomach acid is produced, irritating the gastric mucosa.
Stress can be triggered, e.g., by transportation, competitions, training, a change of house, a new rider, unsuitable equipment, or poor posture.
4. Dental diseases
The teeth are essential for digestion. When feed is chewed, it is broken down and mixed with saliva. Chipped teeth cannot chew well, and the feed is not sufficiently salivated or crushed, which has a detrimental effect on digestion.
For this reason, an expert vet should check the horse´s teeth at least once a year.
5. Administration of painkillers/medication
As with humans, long-term medication administration can promote the formation of stomach ulcers. For this reason, it is essential to ensure that horses are fed a gentle diet on the stomach, especially when using oral pain therapy, and to add stomach protection if necessary.
6. Endotoxins
If pathogens such as E. coli or clostridia proliferate extremely or are killed by an antibiotic, endotoxins can be released. These toxins can cause transformation or inflammation of the gut mucosa. In drastic cases, whole areas of the mucosa can die off.
7. Mycotoxins – the hidden danger in horse feed
Mycotoxins in plants and horse feed are a common but often unnoticed danger to horses’ health. Mycotoxins are natural, secondary metabolites of molds that have a toxic effect on humans and animals and can trigger mycotoxicosis. Contaminated feed can severely affect the horse’s health and, in the worst case, lead to death.
Over 90 % of the world´s feed production is estimated to be contaminated with at least one mycotoxin (see also Global Mycotoxin Report 2023, EW Nutrition. The intake of mycotoxins via hay, grain, silage, or compound feed can hardly be avoided. Mycotoxins are an increasing problem for all horse owners. Scientific studies show that the mycotoxins DON and ZEA are most frequently found in horse feed and, therefore, are also frequently detected in sports horses’ urine and blood samples.
Due to the highly toxic metabolic products, feed contaminated with molds can lead to severe liver and kidney diseases in horses, affect fertility, trigger colic, and promote digestive issues (diarrhea and watery stools).
Mycotoxins Horses
Figure 2: Mycotoxins and their impact on horses
How to protect the horse from mycotoxins?
The first measure against the ingestion of mycotoxins is prevention. Correct pasture management and adequate barn and feed hygiene can contribute to preventing the ingestion of toxins.
However, despite the best prophylactic measures, it is impossible to prevent mycotoxin contamination of feed completely. As mycotoxins are not visible, analyzing the feed regarding mycotoxin contamination is recommended.
To protect your horse from mycotoxins, EW Nutrition developed MasterRisk, a tool for evaluating the risk of mycotoxins. Additionally, EW Nutrition has developed a complementary feed specifically for your horse´s needs in the form of granules. The sophisticated formulation of “Toxi-Pearls” is designed to bind mycotoxins and mitigate the adverse effects of mycotoxin contamination.
The pearls contain a mixture of mycotoxin binder, brewer’s yeast, and herbs:
The contained mycotoxin binder effectively controls the most important feed myco- and endotoxins. It additionally supports the liver and immune system and strengthens the intestinal barrier.
Brewer´s yeast supports the natural strength of the gastrointestinal tract. Due to its high natural content of beta-glucans and mannan-oligosaccharides (MOS), unique surface structure, and the associated high adsorption power, brewer´s yeast has a prebiotic effect on the intestinal microbiome.
The additional unique herbal mixture consists of the typical gastrointestinal herbs oregano, rosemary, aniseed, fennel, and cinnamon. The processed beetroot is a true all-rounder. Literature shows that it has an antioxidant effect and strengthens the immune system. It also promotes bile secretion and, therefore, supports fat digestion.
Conclusion
The horse’s digestive tract is highly sensitive and must be supported by all means. Besides failures in management, such as too long breaks between feedings or too high amounts of feed concentrate, mycotoxins present a high risk in horse nutrition. To prevent horses from intestinal issues, feed and stress management, dental care, and medication in the case of disease must be optimized. Particular attention should be paid to possible mycotoxin contamination. Effective toxin risk management, which consists of analysis, risk evaluation, and adequate toxin risk-managing products, should be implemented.
Consistency in performance: a decisive factor in choosing feed additives
by Madalina Diaconu, Global Manager Gut Health, EW Nutrition
In practical poultry production, multiple stress factors occur simultaneously: nutrition, management, environment, etc.. The effects of these factors are additive, leading to chronic stress, a condition in which animals cannot regain homeostasis and continuously deviate the use of resources to inflammation and restoring the gut barrier-function (Das et al., 2011). As a result, the gut microbiome is altered and oxidative stress ensues (Mishra et al., 2019). In this situation, health and productivity are compromised.
The feed supplied to production animals is designed to help them express their genetic potential. However, some feed components are also continuous inflammatory triggers. Anti-nutritional factors, oxidized lipids, and mycotoxins induce a low-grade inflammatory response (Cardoso Del Pont et al., 2020). Other factors that trigger gut health issues include the environment, management, and pathogens.
Feed interventions have shown to increase productivity and improve gut-related biomarkers, demonstrating a mitigation effect over the challenge factors (Deminicis et al., 2020; Latek et al., 2022).
Meta-analysis of broiler studies shows consistent results
As broilers are continuously challenged during the production period, the effects of an in-feed phytogenic (Ventar D – EW Nutrition GmbH) were extensively researched in broiler meat production. 21 trials in different locations (7 in Europe, 6 in the USA, 4 in Japan, 3 in Middle East, and 2 in India), with different production levels (grouped by EPEF) and challenges were analyzed to establish Ventar D’s benefits for the broiler production industry in terms of performance and sustainability. In all trials, the treatment group consisted of a supplementation of the basal feed with Ventar D at a dose 100 g/ton. The control groups were not supplemented with any gut health improvement feed additive.
Of these 21 trials, 14 had corn/soybean meal-based diets and 7 had high fiber diets (based on wheat and rye, which constituted a challenge as no NSP-enzymes were included). Reused litter (by 12 to 14 flocks, previous to the trial) also was used as a challenge. 18 trials were performed in research facilities and 3 in commercial farms.
Consistency in the results from Ventar D could be demonstrated as 19 out of 21 trials showed an improvement in FCR, lowering 3.4 points on average; 18 /21 trials showed higher body weight, with an average of 64 grams more; and 17 trials showed lower mortality than the control group, averaging 1.19 percentual points of reduction. The phenolic compounds included in Ventar D, such as thymol, possess antioxidant, anti-inflammatory, and antibacterial activities, which account for improving gut health and thus increasing performance in production animals.
The European Poultry Efficiency Factor (EPEF) was used to establish the performance level of each flock. This index is based on the average daily weight gain, mortality, and feed conversion, and takes in consideration the age of the flock at collection, allowing to make comparisons on performance within and between farms.
Of the 21 trials, 10 control groups had an EPEF lower than 375, and were considered of low performance level, in 8 the EPEF was between 375 and 425 and considered of medium performance, and for 3 the performance was considered high having an EPEF of 425 or more.
Ventar D increased performance at all levels (Figure 1). However, the effects were challenge-dependent:
Low performing flocks averaged an 8% increase in EPEF, and high performing flocks increased 4%, indicating that Ventar D can help broilers to overcome challenges commonly found in poultry production, and boost performance even with excellent farm and management conditions. These results concur with a meta-analysis by Valle Polycarpo and collaborators (2022), finding that a microbial challenge can influence the performance of phytogenic feed additives.
Figure 1: % of improvement in EPEF, body weight (BW) and Feed Conversion Rate (FCR) against a non-suplemented control group of IFI suplemented flocks with low (<400), mid (400 – 450) and high (>450) EPEF levels. Significant differences (p<0.05) against a control group (not shown as the improvements against it are depicted) are indicated by (*).
Overall, this analysis demonstrates that effective nutritional interventions can give consistent results and constitute effective tools to help production animals overcome stress and enhance productivity.
Organic acids can play a crucial role in zinc oxide replacement
Dr. Inge Heinzl, Editor EW Nutrition & Juan Antonio Mesonero Escuredo, GTM Swine/GPM Organic Acids EW Nutrition
The use of high levels of Zinc Oxide (ZnO) in the EU before 2022 was one of the most common methods to prevent postweaning diarrhea (PWD) in pig production. Pharmacologically high levels of ZnO (2000-3000 ppm) increase growth and reduce the incidence of enteric bacterial diseases such as post-weaning diarrhea (PWD)( Carlson et al., 1999; Hill et al., 2000; Hill et al., 2001; Poulsen & Larsen, 1995; De Mille et al., 2019).
However, ZnO showed adverse effects, such as the accumulation of heavy metal in the environment, the risk for antimicrobial resistance (AMR), and problems of mineral toxicity and adverse growth effects when feeding it longer than 28 days (Jensen et al., 2018; Cavaco et al., 2011; Vahjen, 2015; Romeo et al., 2014; Burrough et al., 2019). To replace ZnO in pig production, let us first look at its positive effects to know what we must compensate for.
ZnO has a multifactorial mode of action
ZnO shows several beneficial characteristics that positively influence gut health, the immune system, digestion, and, therefore, also overall health and growth performance.
Figure 1. Beneficial effects and ZnO mode of action in postweaning piglets
1. ZnO acts as an antimicrobial
Concerning the antimicrobial effects of ZnO, different possible modes of action are discussed:
ZnO in high dosages generates reactive oxygen species (ROS) that can damage the bacterial cell walls (Pasquet et al., 2014)
The death of the bacterial cell due to direct contact of the metallic Zn to the cell (Shearier et al., 2016)
Intrinsic antimicrobial properties of the ZnO2+ ions after dissociation. The uptake of zinc into cells is regulated by homeostasis. A concentration of the ZnO2+ ions higher than the optimal level of 10-7 to 10-5 M (depending on the microbial strain) allows the invasion of Zn2+ ions into the cell, and the zinc starts to be cytotoxic (Sugarman, 1983; Borovanský et al., 1989).
ZnO shows activity against, e.g., Staphylococcus aureus, Pseudomonas aeruginosa, E. coli,Streptococcus pyogenes, and other enterobacteria (Ann et al., 2014; Vahjen et al., 2016). However, Roselli et al. (2003) did not see a viability-decreasing effect of ZnO on ETEC.
2. ZnO modulates the immune system
Besides fighting pathogenic organisms as described in the previous chapter and supporting the immune system, ZnO is an essential trace element and has a vital role in the immune system. ZnO improves the innate immune response, increasing phagocytosis and oxidative bursts from macrophages and neutrophils. It also ameliorates the adaptative immune response by increasing the number of T lymphocytes (T cells) in general and regulatory T lymphocytes (T-regs) in particular. These cells control the immune response and inflammation (Kloubert et al., 2018). Macrophage capacity for phagocytosis (Ercan and Bor, 1991) and to kill parasites (Wirth et al., 1989), and also the killing activity of natural killer cells depends on Zn (Rolles et al., 2018). By reducing bacterial adhesion and blocking bacterial invasion, ZnO disburdens the immune system (Roselli et al., 2003).
ZnO reduces the expression of several proinflammatory cytokines induced by ETEC (Roselli et al., 2003). Several studies have also shown a modulation effect on intestinal inflammation, decreasing levels of IFN-γ, TNF-α, IL-1ß and IL-6, all pro-inflammatory, in piglets supplemented with ZnO (Zhu et al., 2017; Grilli et al., 2015).
3. ZnO improves digestion and promotes growth
Besides protecting young piglets against diarrhea, the goal is to make them grow optimally. For this target, an efficient digestion and a high absorption of nutrients is essential. Stimulating diverse pancreatic enzymes such as amylase, carboxypeptidase A, trypsin, chymotrypsin, and lipase increases digestibility (Hedemann et al., 2006; Pieper et al., 2015). However, Pieper et al. (2015) also showed that a long-term supply of very high dietary zinc triggers oxidative stress in the pancreas of piglets.
By stimulating the secretion of ghrelin at the stomach level and thereby promoting the release of insulin-like growth factor (IGF-1) and cholecystokinin (CCK), ZnO enhances muscle protein synthesis, cell proliferation, and feed intake (Yin et al., 2009; MacDonald et al., 2000)).
The result of improved digestion is increased body weight and average daily gain, which can be seen, e.g., in a study by Zhu et al. (2017).
4. ZnO protects the intestinal morphology
ZnO prevents the decrease of the trans-endothelial electrical resistance (TEER), usually occurring in the case of inflammation, by downregulating TNF-α and IFN-γ. TNF-α, as well as IFN-γ, increase the permeability of the epithelial tight junctions and, therefore, the intestinal barrier (Al-Sadi et al., 2009).
The enterotrophic and anti-apoptotic effect of ZnO is reflected by a higher number of proliferating and PCNA-positive cells and an increased mucosa surface in the ileum (higher villi, higher villi/crypt ratio)(Grilli et al., 2015). Zhu et al. (2017) also saw an increase in villus height in the duodenum and ileum and a decrease in crypt depth in the duodenum due to the application of 3000 mg of ZnO/kg. Additionally, they could notice a significant (P<0.05) upregulation of the mRNA expression of the zonula occludens-1 and occluding in the mucosa of the jejunum of weaned piglets.
In a trial conducted by Roselli et al. (2003), the supplementation of 0.2 mmol/L ZnO prevented the disruption of the membrane integrity when human Caco-2 enterocytes were challenged with ETEC.
5. ZnO acts antioxidant
The antioxidant effect of ZnO was shown in a study conducted by Zhu et al., 2017. They could demonstrate that the concentration of malondialdehyde (MDA), a marker for lipid peroxidation, decreased on day 14 or 28, and the total concentration of superoxide dismutase (SOD), comprising enzymes that transform harmful superoxide anions into hydrogen peroxide, increased on day 14 (P<0.05). Additionally, Zn is an essential ion for the catalytic action of these enzymes.
Which positive effects of ZnO can be covered by organic acids (OAs)?
1. OAs act antimicrobial
OAs, on the one hand, lower the pH in the gastrointestinal tract. Some pathogenic bacteria are susceptible to low pH. At a pH<5, the proliferation of, e.g., Salmonella, E. coli, and Clostridium is minimized. The good thing is that some beneficial bacteria, such as lactobacilli or bifidobacteria, survive as they are acid-tolerant. The lactobacilli, on their side, can produce hydrogen peroxide, which inhibits, e.g., Staphylococcus aureus or Pseudomonas spp. (Juven and Pierson, 1996).
Besides this more indirect mode of action, a more direct one is also possible: Owing to their lipophilic character, the undissociated form of OAs can pass the bacterial membrane (Partanen and Mroz, 1999). The lower the external pH, the more undissociated acid is available for invading the microbial cells. Inside the cell, the pH is higher than outside, and the OA dissociates. The release of hydrogen ions leads to a decrease in the internal pH of the cell and to a depressed cell metabolism. To get back to “normal conditions”, the cell expels protons. However, this is an energy-consuming process; longer exposure to OAs leads to cell death. The anion remaining in the cell, when removing the protons, disturbs the cell’s metabolic processes and participates in killing the bacterium.
These theoretical effects could be shown in a practical trial by Ahmed et al. (2014). He fed citric acid (0.5 %) and a blend of acidifiers composed of formic, propionic, lactic, and phosphoric acid + SiO2 (0.4 %) and saw a reduction in fecal counts of Salmonella and E. coli for both groups.
2. OAs modulate the immune system
The immune system is essential in the pig’s life, especially around weaning. Organic acids have been shown to support or stimulate the immune system. Citric acid (0.5%), as well as the blend of acidifiers mentioned before (Ahmed et al., 2014), significantly increased the level of serum IgG. IgG is part of the humoral immune system. They mark foreign substances to be eliminated by other defense systems.
Ren et al. (2019) could demonstrate a decrease in plasma tumor necrosis factor-α that regulates the activity of diverse immune cells. He also found lower interferon-γ and interleukin (Il)-1ß values in the OA group than in the control group after the challenge with ETEC. This trial shows that inflammatory response can be mitigated through the addition of organic acids.
3. OAs improve digestion and promote growth
In piglets, the acidity in the stomach is responsible for the activation and stimulation of certain enzymes. Additionally, it keeps the feed in the stomach for a longer time. Both effects lead to better digestion of the feed.
In the stomach, the conversion of pepsinogen to pepsin, which is responsible for protein digestion, is catalyzed under acid conditions (Sanny et al., 1975)group. Pepsin works optimally at two pH levels: pH 2 and pH 3.5 (Taylor, 1959). With increasing pH, the activity decreases; at pH 6, it stops. Therefore, a high pH can lead to poor digestion and undigested protein arriving in the intestine.
These final products of pepsin protein digestion are needed in the lower parts of the GIT to stimulate the secretion of pancreatic proteolytic enzymes. If they do not arrive, the enzymes are not activated, and the inadequate protein digestion continues. Additionally, gastric acid is the primary stimulant for bicarbonate secretion in the pancreas, neutralizing gastric acid and providing an optimal pH environment for the digestive enzymes working in the duodenum.
As already mentioned, the pH in the stomach influences the transport of digesta. The amount of digesta being transferred from the stomach to the small intestine is related to the acidity of the chyme leaving the stomach and arriving in the small intestine. Emptying of the stomach can only take place when the duodenal chyme can be neutralized by pancreatic or other secretions (Pohl et al., 2008); so, acid-sensitive receptors provide feedback regulation and a higher pH in the stomach leads to a faster transport of the digesta and a worse feed digestion.
4. OAs protect the intestinal morphology
Maintaining an intact gut mucosa with a high surface area is crucial for optimal nutrient absorption. Research suggests organic acids play a significant role in improving mucosal health:
Butyric acid promotes epithelial cell proliferation, as demonstrated in an in vitro pig hindgut mucosa study (Sakata et al., 1995). Fumaric acid, serving as an energy source, may locally enhance small intestinal mucosal growth, aiding in post-weaning epithelial cells’ recovery and increasing absorptive surface and digestive capacity (Blank et al., 1999). Sodium butyrate supplementation at low doses influences gastric morphology and function, thickening the stomach mucosa and enhancing mucosal maturation and differentiation (Mazzoni et al., 2008).
Studies show that organic acids affect gut morphology, with a mixture of short-chain and mid-chain fatty acids leading to longer villi (Ferrara et al., 2016) and Na-butyrate supplementation increasing crypt depth and villi length in the distal jejunum and ileum (Kotunia et al., 2004). However, the villi length and mucosa thickness in the duodenum were reduced. Dietary sodium butyrate has been linked to increased microvilli length and cecal crypt depth in pigs (Gálfi and Bokori, 1990).
5. OAs show antioxidant activity
The last characteristic, the antioxidant effect, cannot be provided at the same level as with ZnO; however, Zhang et al. (2019) attest to OAs a certain antioxidant activity. Oxalic, citric, acetic, malic, and succinic acids, which were extracted from Camellia oleifera, also showed good antioxidant activity in a trial conducted by Zhang et al. (2020).
Organic acids are an excellent tool to compensate for the ban on ZnO
The article shows that organic acids have similar positive effects as zinc oxide. They act antimicrobial, modulate the immune system, maintain the gut morphology, fight pathogenic microbes, and also act – slightly – antioxidant. Additionally, they have a significant advantage: they are not harmful to the environment. Organic acids used in the proper pH range and combination are good tools for replacing zinc oxide.
References on request
Decoding the connection between stress, endotoxins, and poultry health
By Technical Team, EW Nutrition
Stress can be defined as any factor causing disruptions to homeostasis, which triggers a biological response to regain equilibrium. We can distinguish four major types of stressors in the poultry industry:
Technological: related with management events and conditions
Nutritional: involving nutritional disbalances, feed quality and feed management
Pathogenic: comprising health challenges.
Environmental: changes in environment conditions
In practical poultry production, multiple stress factors occur simultaneously. Their effects are also additive, leading to chronic stress. The animals are not regaining homeostasis and continuously deviate the use of resources through inflammation and the gut barrier-function, thus leading to microbiome alteration. As a consequence, welfare, health, and productivity are compromised.
What are endotoxins?
Bacterial lipopolysaccharides (LPS), also known as endotoxins, are the main components of the outer membrane of all Gram-negative bacteria and are essential for their survival. LPS have direct contact with the bacteria’s surroundings and function as a protection mechanism against the host’s immunological response and chemical attacks from bile salts, lysozymes, or other antimicrobial agents.
Gram-negative bacteria are part of animals’ microbiota; thus, there are always LPS in the intestine. Under optimal conditions, this does not affect the animals, because intestinal epithelial cells are not responsive to LPS when stimulated from the apical side. In stress situations, the intestinal barrier function is impaired, allowing the passage of endotoxins into the blood stream. When LPS are detected by the immune system either in the blood or in the basolateral side of the intestine, inflammation and changes in the gut epithelial structure and functionality occur.
The gut is critically affected by stress
Even when there is no direct injury to the gut, signals from the brain can modify different functions of the intestinal tract, including immunity. Stress can lead to functional disorders, as well as to inflammation and infections of the intestinal tract. Downstream signals act via the brain–gut axis, trigger the formation of reactive oxygen and nitrogen species as well as local inflammatory factors, and circulating cytokines, affecting intestinal homeostasis, microbiome, and barrier integrity.
Stress then results in cell injury, apoptosis, and compromised tight junctions. For this reason, luminal substances, including toxins and pathogens, leak into the bloodstream. Additionally, under stress, the gut microbiome shows and increment on Gram-negative bacteria (GNB). For instance, a study by Minghui Wang and collaborators (2020) found an increase of 24% in GNB and lower richness, in the cecum of pullets subjected to mild heat stress (increase in ambient temperature from 24 to 30°C).
Both these factors, barrier damage and alterations in the microbiome, facilitate the passage of endotoxins into the blood stream, which promotes systemic chronic inflammation.
What categories of stress factors trigger luminal endotoxins’ passage into the bloodstream?
Technological stress
Various management practices and events can be taken as stressors by the animals’ organism. One of the most common examples is stocking density, defined as the number of birds or the total live weight of birds in a fixed space. High levels are associated with stress and loss of performance.
A study from the Chung-Ang University in 2019 found that broilers with a stocking density of 30 birds/m2 presented two times more blood LPS than birds kept at half of this stocking density. Moreover, the body weight of the birds in the high-density group was 200g lower than the birds of the low-density group. The study concluded that high stocking density is a factor that can disrupt the intestinal barrier.
Nutritional stress
The feed supplied to production animals is designed to contribute to express their genetic potential, though some feed components are also continuous inflammatory triggers. Anti-nutritional factors, oxidized lipids, and mycotoxins induce a low-grade inflammatory response.
For instance, when mycotoxins are ingested and absorbed, they trigger stress and impair immunity in animals. Their effects start in gastrointestinal tract and extend from disrupting immunity to impairing the intestinal barrier function, prompting secondary infections. Mycotoxins can increase the risk of endotoxins in several ways:
By alterations in the immune response, low doses of mycotoxins, such as trichothecenes, induce the upregulation of pro-inflammatory cytokines. A possible synergy can be inferred as when they are together, the effects may be prolonged and require a lower dosage to be triggered.
A study conducted by EW Nutrition (Figure 1) shows an increase in intestinal lesions and blood endotoxins after a mycotoxin challenge of 200pbb of Aflatoxin B1 + 360ppb Ochratoxin in broilers at 21 days of age. The challenged birds show two times more lesions and blood endotoxins than the ones in the unchallenged control. The use of the right mitigation strategy, a product based on bentonite, yeast cell walls, and phytogenics (EW Nutrition GmbH) successfully prevented these effects as it not only mitigates mycotoxins, but also targets endotoxins in the gut.
Figure 1 Blood LPS and intestinal lesion score of broilers challenged with 200ppb AFB1 + 350 ppb OTA from 1 to 21 days of age without and with an anti-toxin product from EW Nutrition GmbH (adapted from Caballero et al., 2021)
Pathogenic stress
Intestinal disease induces changes in the microbiome, reducing diversity and allowing pathogens to thrive. In clinical and subclinical necrotic enteritis (NE), the intestinal populations of GNB, including Salmonella and E.coli also increases. The lesions associated with the pathogen compromise the epithelial permeability and the intestinal barrier function, resulting in translocation of bacteria and LPS (Figure 5) into the bloodstream and internal organs.
Figure 2 Increase in systemic LPS (vs a healthy control) after a NE challenge (adapted from Chen et al., 2015, Li et al., 2018 & Liu at al., 2018)
Environmental stress
Acute and chronic heat and cold stress increases gut permeability, by increasing intestinal oxidative stress and disrupting the expression of tight junction proteins. This results in the damage and destruction of intestinal cells, inflammation, and imbalance of the microbiota. An increased release and passage of endotoxins has been demonstrated in heat stress (Figure 3), as well as a higher expression of TLR-4 and inflammation.
Figure 3 Systemic LPS increase (in comparison with a non-stressed control) after different heat stress challenges in broilers: 16°C increased for 2, 5 and 10 hours (Huang et al., 2018); 9°C increased for 24 and 72 hours (Nanto-Hara et al., 2020); 10°C continuously for 3 and 10 days, and 15°C 4 hours daily for 3 and 10 days (Alhenaky et al., 2017)
Zhou and collaborators (2021) showed that 72 hours of low temperature treatment in young broilers increased intestinal inflammation and expression of tight junction proteins, while higher blood endotoxins indicate a disruption of the intestinal barrier. As a consequence, the stress decreased body gain and increased the feed conversion rate.
An experiment conducted by EW Nutrition GmbH with the objective of evaluating the ability of a toxin mitigation product to ameliorate heat-stress induced LPS. For the experiment, 1760 Cobb 500 pullets were divided into two groups, and each was placed in 11 pens of 80 hens, in a single house. One of the groups received feed containing 2kg/ton of the product from the first day. From week 8 to week 12, the temperature of the house was raised 10°C for 8 hours every day.
Throughout the heat stress period, blood LPS (Fig 4) was lower in the pullets receiving the product, which allowed lower inflammation, as evidenced by the lower expression of TLR4 (Fig. 5). Oxidative stress was also mitigated with the help of the combination of phytomolecules in the product, obtaining 8.5% improvement on serum total antioxidant capacity (TAC), supported by an increase in in superoxide dismutase (SOD glutathione peroxidase (GSH) and a decrease in malondialdehyde (MDH).
Figures 4 and 5 – Blood LPS and expression of toll-like receptor 4 (TLR4) in lymphocites of pullets before (wk 6) and during heat stress (wk 9 and 10). (*) indicates significant differences (P<0,05), and (‡) a tendency to be different against the control group (P<0,1).
In practice: there is no silver bullet
In commercial poultry production, a myriad stressors may occur at the same time and some factors trigger a chain of events that work to the detriment of animal health and productivity. Reducing the solution to the mitigation of LPS is a deceitfully simplistic approach. However, this should be part of a strategy to achieve better animal health and performance. In fact, EW Nutrition’s toxin mitigation product alone helped the pullets to achieve 3% improvement in body weight and 9 points lower cumulative feed conversion (Figure 6).
Figure 6
Keeping the animals as free of stress as possible is a true priority for poultry producers, as it promotes animal health as well as the integrity and function of the intestinal barrier. Biosecurity, good environment, nutrition and good management practices are crucial; the use of feed additives to reduce the consequences of unavoidable stress also critically supports the profitability of poultry operations.
Salmonella in pigs: a threat for humans and a challenge for pig producers
By Dr. Inge Heinzl, Editor, EW Nutrition
Salmonellosis is third among foodborne diseases leading to death (Ferrari, 2019). More than 91,000 human cases of Salmonellosis are reported by the EU each year, generating overall costs of up to €3 billion a year (EFSA, 2023), 10-20% of which are attributed to pork consumption (Soumet, 2022). The annual costs arising from the resulting human health losses in 2010 were about €90 million (FCC Consortium, 2010). Take the example of Ireland, where a high prevalence of Salmonella in lymph nodes still shows a severe issue pre-slaughter and a big challenge for slaughterhouses to stick to the process hygiene requirements (Deane, 2022).
Several governments already have monitoring programs in place, and the farms are categorized according to the salmonella contamination of their pigs. In some countries, e.g., Denmark, an economic penalty of 2% of the carcass value must be paid if the farm has level 2 (intermediate seroprevalence) and 4-8% if the level is 3. Other countries, e.g., Germany, the UK, Ireland, or the Netherlands, use quality assurance schemes. The farmers can only sell their carcasses under this label if their farm has a certain level.
Let’s take a quick look at the genus of Salmonella
Salmonellas are rod-shaped gram-negative bacteria of the family of enterobacteria that use flagella for their movement. They were named after the American vet Daniel Elmer Salmon. The genus of Salmonella consists of two species (S. bongori and S. enterica with seven subspecies) with in total more than 2500 serovars (see Figure 1). The effects of the different serovars can range from asymptomatic carriage to severe invasive systemic disease (Gal-Mor, 2014). All Salmonella serovars generally can cause disease in humans; the rosa-marked ones already showed infections.
Figure 1: the genus of Salmonella with Salmonella serovars relevant for pigs (according to Bonardi, 2017: Salmonella in the pork production chain and its impact on human health in the European Union)
Within the group of Salmonella, some serovars can only reside in one or few species, e.g., S. enterica spp. enterica Serovar Dublin (S. Dublin) in bovines (Waldron, 2018) or S. Cholerasuis in pigs (Chiu, 2004). An infection in humans with these pathogens is often invasive and life-threatening (WHO, 2018). On the contrary, serovars like S. Typhimurium and S. Enteritidis are not host-specific and can cause disease in various species.
The serotypes S. Typhi and S. Paratyphi A, B, or C are highly adapted to humans and only for them pathogenic; they are responsible for the occurrence of typhus.
Transmission of Salmonella mostly happens via contaminated food
The way of transmission to humans depends on the serovar:
Human-specific and, therefore, only in humans and higher primates residing serovars S. Typhi and Paratyphi A, B, or C (typhoidal) are excreted via feces or urine. Therefore, any food or water contaminated with the feces or urine of infected people can transmit this disease (Government of South Australia, 2023). Typhoid and paratyphoid Salmonellosis occur endemic in developing countries with the lack of clean water and, therefore, inadequate hygiene (Gal-Mor, 2014).
Serovars which can cause disease in humans and animals (non-typhoidal), can be transmitted by
– animal products such as milk, eggs, meat
– contact with infected persons/animals (pigs, cows, pets, reptiles…) or
– other feces- or urine-contaminated products such as sprouts, vegetables, fruits….
Farm animals take salmonellas from their fellows, contaminated feed or water, rodents, or pests.
Symptoms of Salmonellosis can be severe
In the case of typhoid or paratyphoid Salmonellosis, the onset of illness is gradual. People can suffer from sustained high fever, unwellness, severe headache, and decreased appetite, but also from an enlarged spleen irritating the abdomen and dry cough.
A study conducted in Thailand with children suffering from enteric fever caused by the typhoid serovars S. Typhi and Paratyphi showed a sudden onset of fever and gastrointestinal issues (diarrhea), rose spots, bronchitis, and pneumonia (Thisyakorn et al., 1987)
The non-typhoid Salmonellosis is typically characterized by an acute onset of fever, nausea, abdominal pain with diarrhea, and sometimes vomiting (WHO, 2018). However, 5% of the persons – children with underlying conditions, e.g., babies, or people who have AIDS, malignancies, inflammatory bowel disease, gastrointestinal illness caused by non-typhoid serovars, and hemolytic anemia, or receiving an immunosuppressive therapy can be susceptible to bacteremia. Additionally, serovars like S. Cholerasuis or S. Dublin are apt to develop bacteremia by entering the bloodstream with little or no involvement of the gut (Chiu, 1999). In these cases, consequences can be septic arthritis, pneumonia, peritonitis, cutaneous abscess, mycotic aneurysm, and sometimes death (Chen et al., 2007; Chiu, 2004, Wang et al., 1996).
In pigs, S. Cholerasuis causes high fever, purple discolorations of the skin, and thereinafter diarrhea. The mortality rate in pigs suffering from this type of Salmonellosis is high. Barrows orally challenged with S. Typhimurium showed elevated rectal temperature by 12h, remaining elevated until the end of the study. Feed intake decreased with a peak at 48h after the challenge and remained up to 120h after the challenge. Daily gain reduced during the following two weeks after infection. A higher plasma cortisol level and a lower IGF-I level could also be noticed. All these effects indicate significant changes in the endocrine stress and the somatotropic axis, also without significant alterations in the systemic pro-inflammatory mediators (Balaji et al., 2000)
To protect humans, Salmonella in pork must be restraint
There are three main steps to keep the contamination of pork as low as possible:
Keeping Salmonella out of the pig farm
Minimizing spreading if Salmonella is already on the farm
Minimizing contamination in the slaughterhouse
1. How to keep Salmonella out of the pig farm?
To answer this question, we must look at how the pathogen can be transported to the farm. According to the Code of Practice for the Prevention and Control of Salmonella on Pig Farms (Ministry of Agriculture, Fisheries and Food and the Scottish Executive Rural Affairs Department), there are several possibilities to infiltrate the pathogen into the farm:
Diseased pigs or pigs which are ill but don’t show any symptoms
Feeding stuff or bedding contaminated with dung
Pets, rodents, wild birds, or animals
Farm personnel or visitors
Equipment or vehicles
Caution with purchased animals!
To minimize/prevent the entry of Salmonella into the livestock, bought-in animals must come from reputable breeding farms with a salmonella monitoring system in place. As possible carrier animals are more likely to excrete Salmonella when stressed; they should be kept in isolation after purchasing. Additionally, the animals must go through a disinfectant foot bath before entering the farm.
Keep rodents, wild animals, and vermin in check!
Generally, the production site must be kept clean and as unattractive as possible for all these animals. Rests of feed must be removed, and dead animals and afterbirths must be promptly and carefully disposed of. A well-planned baiting and trapping policy should be in place to effectively control rodents.
Only selected people should enter the hog houses
In any case, the number of persons entering the hog house must be kept as low as possible. Farmworkers should be trained in the principles of hygiene. They should wear adequate clothing (waterproof boots and protective overalls) that can be easily cleaned/laundered and disinfected. The clothes/shoes should always be used only at this site. Thorough hand washing and the disinfection of the boots when entering and leaving the pig unit are a must.
If visits are necessary, the visitors should take the same measures as the farm workers. And, of course, they should not have had contact with another pig farm during the last 48 hours.
Keep pens, farm equipment, and vehicles clean!
Farm equipment should not be shared with other farms. If this cannot be avoided, it must be cleaned and disinfected before re-entering the farm. Also, the vehicles for the transport of the animals must be cleaned and disinfected as soon as possible after usage, as contaminated transporters always pose the risk of infection.
Feed should be Salmonella-free!
To get high feed quality, the feed should be purchased from feed mills/sources with a well-functioning bacterial control to guarantee the absence of Salmonella. It is essential that birds, domestic and wild animals cannot enter the feed stores.
It is also advised to keep dry feed dry as possibly contaminating Salmonella can multiply in such humid conditions. Additionally, all feed bins and delivery pipes for dry and wet feed must be consciously cleaned, and the damp feed pipes also disinfected.
The change from pellets to mash could be helpful as the pellets facilitate Salmonella colonization by stimulating the secretion of mucins (Hedemann et al., 2005).
For sanitation of the feed, we offer organic acids (Acidomix product range) or mixtures of organic acids and formaldehyde in countries where formaldehyde products are allowed (Formycine) to decrease the pathogenic load of the feed materials. In vitro trials show the effectiveness of the products:
For the in vitro trial with Formycine, autoclaved feed samples were inoculated with Salmonella enteritidis serovar Typhimurium DSM 19587 strain to reach a Salmonella contamination of 106 CFU/g of feed. After incubating at room temperature for three hours, Formycine Liquido was added to the contaminated feed samples at 0, 500, 1000, and 2000 ppm. The control and inoculated feed samples were further incubated at room temperature, and Salmonella counts (CFU/g) were carried out at 24, 48, 72 hours and on day 15. The limit of Salmonella detection was set at 100 CFU/g (102). Results are shown in figure 2.
Fig. 2: Effect of treatment time and different inclusion levels of Formycine Liquido on the Salmonella count in feed
As important as uncontaminated feed is clean water for drinking. It can be achieved by taking the water from a main or a bacteriologically controlled water borehole. Regular cleaning/disinfection of the tanks, pipes, and drinkers is essential.
Bedding should be Salmonella-free
Straw material containing feces of other animals (rodents, pets) always carries the risk of Salmonella contamination. Also, wet or moldy bedding is not recommended because it is an additional challenge for the animal. To optimize the quality of bedding, the straw should be bought from reliable and as few as possible sources. The material must be stored dry and as far as practicable from the pig buildings (Ministry of Agriculture, Fisheries and Food & Scottish Executive Rural Affairs Department, 2000).
Vaccination is a beneficial measure
For the control of Salmonella in swine herds, vaccination is an effective tool. De Ridder et al. (2013) showed that an attenuated vaccine reduced the transmission of Salmonella Typhimurium in pigs. The vaccination with an attenuated S. Typhimurium strain, followed by a booster vaccination with inactivated S. Cholerasuis, showed better effects than an inactivated S. Cholerasuis vaccine alone (Alborali et al., 2017). Bearson et al. (2017) could delimitate transmission through less shedding and protect the animals against systemic disease.
To achieve the best effects, the producer must understand the diversity of Salmonella serovars to choose the most promising vaccination strategy (FSIS, 2023).
2. How to minimize the spreading of Salmonella on the farm?
If there are already cases of Salmonella on the farm, infected animals must be separated from the rest of the herd. Small batch sizes are beneficial, as well as not mixing different litters after weaning. If feasible, separate units for different production phases with an all-in/all-out system could break the reinfection cycle and help reduce Salmonella contamination on the farm. And also in this case, vaccination is helpful.
Salmonella doesn’t like acid conditions
An effective tool is acidifying the feed with organic acids, as Salmonella doesn’t like acid conditions. A trial was conducted with Acidomix AFG and Acidomix AFL to show their effects against Salmonella. For the test, 105 CFU/g of Salmonella enterica ser. Typhimurium was added to feed containing 1000 ppm, 2000 ppm, and 3000 ppm of Acidomix AFG or AFL. The stomach and intestine were simulated in vitro by adjusting the pH with HCl and NaHCO3 as follows:
Stomach 2.8
Intestine 6.8-7.0
After the respective incubation, the microorganisms were recovered from feed and plated on an appropriate medium for CFU counting. The results are shown in figures 3 and 4.
Combi
Figures 3 + 4: Effects of different concentrations of Acidomix AFG and Acidomix AFL against Salmonella enterica ser. Typhimurium in feed
Phytomolecules can support pigs against Salmonella
Plant compounds or phytomolecules can also be used against Salmonella in pigs. Some examples of phytomolecules to be used are Piperine, Allicin, Eugenol, and Carvacrol. Eugenol, e.g., increases the permeability of the Salmonella membrane, disrupts the cytoplasmic membrane, and inhibits the production of bacterial virulence factors (Keita et al., 2022; Mak et al., 2019). Thymol and Carvacrol interact with the cell membrane by H bonding, also resulting in a higher permeability.
An already published in vitro trial conducted with our product Ventar D also showed excellent effects against Salmonella while sparing the beneficial gut flora. A further trial once more demonstrated the susceptibility of Salmonella to Ventar D. It showed that Ventar D controls Salmonella by suppressing their motility and, at higher concentrations, inactivating the cells (see figures 5 + 6):
Figure 5: S. enterica motility test: on the left side – control; on the right side – motility medium containing.750 µg/mL of Ventar
Fig 6 . Disk diffusion assay employing S. enterica. upper left side – disk containing 10 µL of Ventar; upper right – 5 µL; lower left – control; lower right – 1µL.
In addition to the direct Salmonella-reducing effect, essential oils / secondary plant compounds / phytomolecules improve digestive enzyme activity and digestion, leading to increased nutrient absorption and better feed conversion (Windisch et al., 2008).
3. How can the farmer keep Salmonella contamination low in the slaughterhouse?
In general, the slaughterhouse personnel is responsible for adequate hygiene management to prevent contamination of carcasses and meat. However, also the farmer can make his contribution to maintain the risk of contamination in the slaughterhouse as low as possible. A study by Vieira-Pinto (2006) revealed that one Salmonella-positive pig can contaminate several other carcasses.
According to a trial conducted by Hurd et al. (2002), infection and, therefore, “contamination” of other pigs can rapidly occur, meaning that cross-contamination is a topic during transport to the slaughterhouse and in the lairages when the pigs come together with animals from other farms. The stress to which the pigs are exposed influences physiological and biochemical processes. The microbiome and animal’s immunity are affected, leading to higher excretion of Salmonella during transport and in the lairages. So, the animals should not be stressed during loading and unloading or transportation. The trailer poses a further risk of infection if it was not cleaned and disinfected before. So, reliable people who treat the animals well and keep their trailers clean should be chosen for transportation.
Pig producers are obliged to keep Salmonella in check – phytomolecules can help
At least in the EU, pig producers have the big duty to keep Salmonella low in their herds; otherwise, they will have financial losses. They are not only responsible for their farm, but also the slaughterhouses count on them. Besides the standard strict hygiene management and vaccination, farmers can use products provided by the industry to sanitize feed but also to support their animals directly with phytomolecules acting against pathogens and supporting gut health.
All these measures together should be a solution to the immense challenge of Salmonella, to protect people and prevent economic losses.
References:
Alborali, Giovanni Loris, Jessica Ruggeri, Michele Pesciaroli, Nicola Martinelli, Barbara Chirullo, Serena Ammendola, Andrea Battistoni, Maria Cristina Ossiprandi, Attilio Corradi, and Paolo Pasquali. “Prime-Boost Vaccination with Attenuated Salmonella Typhimurium Δznuabc and Inactivated Salmonella Choleraesuis Is Protective against Salmonella Choleraesuis Challenge Infection in Piglets.” BMC Veterinary Research 13, no. 1 (2017): 284. https://doi.org/10.1186/s12917-017-1202-5.
Balaji, R, K J Wright, C M Hill, S S Dritz, E L Knoppel, and J E Minton. “Acute Phase Responses of Pigs Challenged Orally with Salmonella Typhimurium.” Journal of Animal Science 78, no. 7 (2000): 1885. https://doi.org/10.2527/2000.7871885x.
Bearson, Bradley L, Shawn M. Bearson, Brian W Brunelle, Darrell O Bayles, In Soo Lee, and Jalusa D Kich. “Salmonella Diva Vaccine Reduces Disease, Colonization, and Shedding Due to Virulent S. Typhimurium Infection in Swine.” Journal of Medical Microbiology 66, no. 5 (2017): 651–61. https://doi.org/10.1099/jmm.0.000482.
Brenner Michael, G, M Cardoso, and S Schwarz. “Molecular Analysis of Salmonella Enterica Subsp. Enterica Serovar Agona Isolated from Slaughter Pigs.” Veterinary Microbiology 112, no. 1 (2006): 43–52. https://doi.org/10.1016/j.vetmic.2005.10.011.
Chen, P.-L., C.-M. Chang, C.-J. Wu, N.-Y. Ko, N.-Y. Lee, H.-C. Lee, H.-I. Shih, C.-C. Lee, R.-R. Wang, and W.-C. Ko. “Extraintestinal Focal Infections in Adults with Non-typhoid Salmonella Bacteraemia: Predisposing Factors and Clinical Outcome.” Journal of Internal Medicine 261, no. 1 (2007): 91–100. https://doi.org/10.1111/j.1365-2796.2006.01748.x.
Chiu, Cheng-Hsun, Lin-Hui Su, and Chishih Chu. “Salmonella EntericaSerotype Choleraesuis: Epidemiology, Pathogenesis, Clinical Disease, and Treatment.” Clinical Microbiology Reviews 17, no. 2 (2004): 311–22. https://doi.org/10.1128/cmr.17.2.311-322.2004.
De Ridder, L., D. Maes, J. Dewulf, F. Pasmans, F. Boyen, F. Haesebrouck, E. Méroc, P. Butaye, and Y. Van der Stede. “Evaluation of Three Intervention Strategies to Reduce the Transmission of Salmonella Typhimurium in Pigs.” The Veterinary Journal 197, no. 3 (2013): 613–18. https://doi.org/10.1016/j.tvjl.2013.03.026.
Deane, Annette, Declan Murphy, Finola C. Leonard, William Byrne, Tracey Clegg, Gillian Madigan, Margaret Griffin, John Egan, and Deirdre M. Prendergast. “Prevalence of Salmonella spp. in Slaughter Pigs and Carcasses in Irish Abattoirs and Their Antimicrobial Resistance.” Irish Veterinary Journal 75, no. 1 (2022). https://doi.org/10.1186/s13620-022-00211-y.
Edel, W., M. Schothorst, P. A. Guinée, and E. H. Kampelmacher. “Effect of Feeding Pellets on the Prevention and Sanitation of Salmonella Infections in Fattening Pigs1.” Zentralblatt für Veterinärmedizin Reihe B 17, no. 7 (2010): 730–38. https://doi.org/10.1111/j.1439-0450.1970.tb01571.x.
EFSA. “Salmonella.” European Food Safety Authority. Accessed August 7, 2023. https://www.efsa.europa.eu/en/topics/topic/salmonella.
Elbediwi, Mohammed, Daiwei Shi, Silpak Biswas, Xuebin Xu, and Min Yue. “Changing Patterns of Salmonella Enterica Serovar Rissen from Humans, Food Animals, and Animal-Derived Foods in China, 1995–2019.” Frontiers in Microbiology 12 (2021). https://doi.org/10.3389/fmicb.2021.702909.
Elnekave, Ehud, Samuel Hong, Alison E Mather, Dave Boxrud, Angela J Taylor, Victoria Lappi, Timothy J Johnson, et al. “Salmonella Enterica Serotype 4,[5],12:I:- In Swine in the United States Midwest: An Emerging Multidrug-Resistant Clade.” Clinical Infectious Diseases 66, no. 6 (2018): 877–85. https://doi.org/10.1093/cid/cix909.
Ferrari, Rafaela G., Denes K. Rosario, Adelino Cunha-Neto, Sérgio B. Mano, Eduardo E. Figueiredo, and Carlos A. Conte-Junior. “Worldwide Epidemiology of Salmonella serovars in Animal-Based Foods: A Meta-Analysis.” Applied and Environmental Microbiology 85, no. 14 (2019). https://doi.org/10.1128/aem.00591-19.
“FSIS Guideline to Control Salmonella in Swine Slaughter and Pork Processing Establishments.” FSIS Guideline to Control Salmonella in Swine Slaughter and Pork Processing Establishments | Food Safety and Inspection Service. Accessed August 14, 2023. https://www.fsis.usda.gov/guidelines/2023-0003.
Gal-Mor, Ohad, Erin C. Boyle, and Guntram A. Grassl. “Same Species, Different Diseases: How and Why Typhoidal and Non-Typhoidal Salmonella Enterica Serovars Differ.” Frontiers in Microbiology 5 (2014). https://doi.org/10.3389/fmicb.2014.00391.
González-Santamarina, Belén, Silvia García-Soto, Helmut Hotzel, Diana Meemken, Reinhard Fries, and Herbert Tomaso. “Salmonella Derby: A Comparative Genomic Analysis of Strains from Germany.” Frontiers in Microbiology 12 (2021). https://doi.org/10.3389/fmicb.2021.591929.
Government of South Australia. Typhoid and paratyphoid – including symptoms, treatment, and prevention, April 3, 2022. https://www.sahealth.sa.gov.au/wps/wcm/connect/public+content/sa+health+internet/conditions/infectious+diseases/typhoid+and+paratyphoid/typhoid+and+paratyphoid+-+including+symptoms+treatment+and+prevention.
Hauser, Elisabeth, Erhard Tietze, Reiner Helmuth, Ernst Junker, Kathrin Blank, Rita Prager, Wolfgang Rabsch, Bernd Appel, Angelika Fruth, and Burkhard Malorny. “Pork Contaminated with SalmonellaEnterica Serovar 4,[5],12:I:−, an Emerging Health Risk for Humans.” Applied and Environmental Microbiology 76, no. 14 (2010): 4601–10. https://doi.org/10.1128/aem.02991-09.
Health and Wellbeing; address=11 Hindmarsh Square, Adelaide scheme=AGLSTERMS.AglsAgent; corporateName=Department for. “Sa Health.” Typhoid and paratyphoid – including symptoms, treatment, and prevention, April 3, 2022. https://www.sahealth.sa.gov.au/wps/wcm/connect/public+content/sa+health+internet/conditions/infectious+diseases/typhoid+and+paratyphoid/typhoid+and+paratyphoid+-+including+symptoms+treatment+and+prevention.
Hedemann, M. S., L. L. Mikkelsen, P. J. Naughton, and B. B. Jensen. “Effect of Feed Particle Size and Feed Processing on Morphological Characteristics in the Small and Large Intestine of Pigs and on Adhesion of Salmonella Enterica Serovar Typhimurium DT12 in the Ileum in Vitro1.” Journal of Animal Science 83, no. 7 (2005): 1554–62. https://doi.org/10.2527/2005.8371554x.
Hendriksen, Susan W.M., Karin Orsel, Jaap A. Wagenaar, Angelika Miko, and Engeline van Duijkeren. “Animal-to-Human Transmission ofSalmonellaTyphimurium DT104A Variant.” Emerging Infectious Diseases 10, no. 12 (2004): 2225–27. https://doi.org/10.3201/eid1012.040286.
Keita, Kadiatou, Charles Darkoh, and Florence Okafor. “Secondary Plant Metabolites as Potent Drug Candidates against Antimicrobial-Resistant Pathogens.” SN Applied Sciences 4, no. 8 (2022). https://doi.org/10.1007/s42452-022-05084-y.
Ministry of Agriculture, Fisheries and Food, and Scottish Executive Rural Affairs Department. “Salmonella on Pig Farms – Code of Practice for the Prevention and Control Of.” ReadkonG.com, 2000. https://www.readkong.com/page/code-of-practice-for-the-prevention-and-control-of-5160969.
Morrow, W.E. Morgan, and Julie Funk. Ms. Salmonella as a Foodborne Pathogen in Pork. North Carolina State University Animal Science, n.d.
Soumet, C., A. Kerouanton, A. Bridier, N. Rose, M. Denis, I. Attig, N. Haddache, and C. Fablet. Report, Salmonella excretion level in pig farms and impact of quaternary ammonium compounds based disinfectants on Escherichia coli antibiotic resistance § (2022).
Thisyakorn, Usa. “Typhoid and Paratyphoid Fever in 192 Hospitalized Children in Thailand.” Archives of Pediatrics & Adolescent Medicine 141, no. 8 (1987): 862. https://doi.org/10.1001/archpedi.1987.04460080048025.
Ung, Aymeric, Amrish Y. Baidjoe, Dieter Van Cauteren, Nizar Fawal, Laetitia Fabre, Caroline Guerrisi, Kostas Danis, et al. “Disentangling a Complex Nationwide Salmonella Dublin Outbreak Associated with Raw-Milk Cheese Consumption, France, 2015 to 2016.” Eurosurveillance 24, no. 3 (2019). https://doi.org/10.2807/1560-7917.es.2019.24.3.1700703.
Vieira-Pinto, M, R Tenreiro, and C Martins. “Unveiling Contamination Sources and Dissemination Routes of Salmonella Sp. in Pigs at a Portuguese Slaughterhouse through Macrorestriction Profiling by Pulsed-Field Gel Electrophoresis.” International Journal of Food Microbiology 110, no. 1 (2006): 77–84. https://doi.org/10.1016/j.ijfoodmicro.2006.01.046.
Waldron, P. “Keeping Cows and Humans Safe from Salmonella Dublin.” Cornell University College of Veterinary Medicine, December 25, 2018. https://www.vet.cornell.edu/news/20181218/keeping-cows-and-humans-safe-salmonella-dublin.
Wang, J.-H., Y.-C. Liu, M.-Y. Yen, J.-H. Wang, Y.-S. Chen, S.-R. Wann, and D.-L. Cheng. “Mycotic Aneurysm Due to Non-Typhi Salmonella: Report of 16 Cases.” Clinical Infectious Diseases 23, no. 4 (1996): 743–47. https://doi.org/10.1093/clinids/23.4.743.
WHO. “Salmonella (Non-Typhoidal).” World Health Organization, February 20, 2018. https://www.who.int/news-room/fact-sheets/detail/salmonella-(non-typhoidal).
Windisch, W., K. Schedle, C. Plitzner, and A. Kroismayr. “Use of Phytogenic Products as Feed Additives for Swine and Poultry1.” Journal of Animal Science 86, no. suppl_14 (2008). https://doi.org/10.2527/jas.2007-0459.
Windisch, W., K. Schedle, C. Plitzner, and A. Kroismayr. “Use of Phytogenic Products as Feed Additives for Swine and Poultry1.” Journal of Animal Science 86, no. suppl_14 (2008). https://doi.org/10.2527/jas.2007-0459.
The future of coccidiosis control
By Madalina Diaconu, Product Manager Pretect D, EW Nutrition and
With costs of over 14 billion USD per year (Blake, 2020), coccidiosis is one of the most devastating enteric challenges in the poultry industry. With regard to costs, subclinical forms of coccidiosis account for the majority of production losses, as damage to intestinal cells results in lower body weight, higher feed conversion rates, lack of flock uniformity, and failures in skin pigmentation. This challenge can only be tackled, if we understand the basics of coccidiosis control in poultry and what options producers have to manage coccidiosis risks.
Current strategies show weak points
Good farm management, litter management, and coccidiosis control programs such as shuttle and rotation programs form the basis for preventing clinical coccidiosis. More successful strategies include disease monitoring, strategic use of coccidiostats, and increasingly coccidiosis vaccines. However, the intrinsic properties of coccidia make these parasites often frustrating to control. Acquired resistance to available coccidiostats is the most difficult and challenging factor to overcome.
Optimally, coccidiosis control programs are developed based on the farm history and the severity of infection. The coccidiostats traditionally used were chemicals and ionophores, with ionophores being polyether antibiotics. To prevent the development of resistance, the coccidiostats were used in shuttle or rotation programs, at which in the rotation program, the anticoccidial changes from flock to flock, and in the shuttle program within one production cycle (Chapman, 1997).
The control strategies, however, are not 100% effective. The reason for that is a lack of diversity in available drug molecules and the overuse of some molecules within programs. An additional lack of sufficient coccidiosis monitoring and rigorous financial optimization often leads to cost-saving but only marginally effective solutions. At first glance, they seem effective, but in reality, they promote resistance, the development of subclinical coccidiosis, expressed in a worsened feed conversion rate, and possibly also clinical coccidiosis.
Market requests and regulations drive coccidiosis control strategies
Changing coccidiosis control strategies has two main drivers: the global interest in mitigating antimicrobial resistance and the consumer’s demand for antibiotic-free meat production.
Authorities have left ionophores untouched
Already in the late 1990s, due to the fear of growing antimicrobial resistance, the EU withdrew the authorization for Avoparcin, Bacitracin zinc, Spiramycin, Virginiamycin, and Tylosin phosphate, typical growth promoters, to “help decrease resistance to antibiotics used in medical therapy”. However, ionophores, being also antibiotics, were left untouched: The regulation (EC) No 1831/2003 [13]of the European Parliament and the Council of 22 September 2003 clearly distinguished between coccidiostats and antibiotic growth promoters. Unlike the antibiotic growth promoters, whose primary action site is the gut microflora, coccidiostats only have a secondary and residual activity against the gut microflora. Furthermore, the Commission declared in 2022 that the use of coccidiostats would not presently be ruled out “even if of antibiotic origin” (MEMO/02/66, 2022) as “hygienic precautions and adaptive husbandry measures are not sufficient to keep poultry free of coccidiosis” and that “modern poultry husbandry is currently only practicable if coccidiosis can be prevented by inhibiting or killing parasites during their development”. In other words, the Commission acknowledged that ionophores were only still authorized because it believed there were no other means of controlling coccidiosis in profitable poultry production.
Consumer trends drove research on natural solutions
Due to consumers’ demand for antibiotic-reduced or, even better, antibiotic-free meat production, intensified industrial research to fight coccidiosis with natural solutions has shown success. Knowledge, research, and technological developments are now at the stage of offering solutions that can be an effective part of the coccidia control program and open up opportunities to make poultry production even more sustainable by reducing drug dependency.
Producers from other countries have already reacted. Different from the handling of ionophores regime in the EU, where they are allowed as feed additives, in the United States, coccidiostats belonging to the polyether-ionophore class are not permitted in NAE (No Antibiotics Ever) and RWE (Raised Without Antibiotics) programs. Instead of using ionophores, coccidiosis is controlled with a veterinary-led combination of live vaccines, synthetic compounds, phytomolecules, and farm management. This approach can be successful, as demonstrated by the fact that over 50% of broiler meat production in the US is NAE. Another example is Australia, where the two leading retail store chains also exclude chemical coccidiostats from broiler production. In certain European countries, e.g., Norway, the focus is increasingly on banning ionophores.
The transition to natural solutions needs knowledge and finesse
In the beginning, the transition from conventional to NAE production can be difficult. There is the possibility to leave out the ionophores and manage the control program only with chemicals of different modes of action. More effective, however, is a combination of vaccination and chemicals (bio-shuttle program) or the combination of phytomolecules with vaccination and/or chemicals (Gaydos, 2022).
Coccidiosis vaccination essentials
When it is decided that natural solutions shall be used to control coccidiosis, some things about vaccination must be known:
There are different strains of vaccines, natural ones selected from the field and attenuated strains. The formers show medium pathogenicity and enable a controlled infection of the flock. The latter, being early mature lower pathogenicity strains, usually cause only low or no post-vaccinal reactions.
A coccidiosis program that includes vaccination should cover the period from the hatchery till the end of the production cycle. Perfect application of the vaccines and effective recirculation of vaccine strains amongst the broilers are only two examples of preconditions that must be fulfilled for striking success and, therefore, early and homogenous immunity of the flock.
Perfect handling of the vaccines is of vital importance. For that purpose, the personnel conducting the vaccinations in the hatchery or on the farms must be trained. In some situations, consistent high-quality application at the farm has shown to be challenging. As a result, interest in vaccine application at the hatchery is growing.
Phytochemicals are a perfect tool to complement coccidiosis control programs
As the availability of vaccines is limited and the application costs are relatively high, the industry has been researching supportive measures or products and discovered phytochemicals as the best choice. Effective phytochemical substances have antimicrobial and antiparasitic properties and enhance protective immunity in poultry infected by coccidiosis. They can be used in rotation with vaccination, to curtail vaccination reactions of (non-attenuated) wild strain vaccines, or in combination with chemical coccidiostats in a shuttle program.
In a recent review paper (El-Shall et al., 2022), natural herbal products and their extracts have been described to effectively reduce oocyst output by inhibiting Eimeria species’ invasion, replication, and development in chicken gut tissues. Phenolic compounds in herbal extracts cause coccidia cell death and lower oocyst counts. Additionally, herbal additives offer benefits such as reducing intestinal lipid peroxidation, facilitating epithelial repair, and decreasing Eimeria-induced intestinal permeability.
Various phytochemical remedies are shown in this simplified adaptation of a table from El-Shall et al. (2022), indicating the effects exerted on poultry in connection to coccidia infection.
Bioactive compound
Effect
Saponins
Inhibition of coccidia:
By binding to membrane cholesterol, the saponins disturb the lipids in the parasite cell membrane. The impact on the enzymatic activity and metabolism leads to cell death, which then induces a toxic effect in mature enterocytes in the intestinal mucosa. As a result, sporozoite-infected cells are released before the protozoa reach the merozoite phase.Support for the chicken:
Saponins enhance non-specific immunity and increase productive performance (higher daily gain and improved FCR, lower mortality rate). They decrease fecal oocyst shedding and reduce ammonia production.
Tannins
Inhibition of coccidia:
Tannins penetrate the coccidia oocyst wall and inactivate the endogenous enzymes responsible for sporulation.Support for the chicken: Additionally, they enhance anticoccidial antibodies’ activity by increasing cellular and humoral immunity.
Flavonoids and terpenoids
Inhibition of coccidia:
They inhibit the invasion and replication of different species of coccidia.Support for the chicken: They bind to the mannose receptor on macrophages and stimulate them to produce inflammatory cytokines such as IL-1 through IL-6 and TNF. Higher weight gain and lower fecal oocyst output are an indication of suppression of coccidiosis.
Artemisinin
Inhibition of coccidia:
Its impact on calcium homeostasis compromises the oocyst wall formation and leads to a defective cell wall and, in the end, to the death of the oocyst. Enhancing the production of ROS directly inhibits sporulation and also wall formation and, therefore, affects the Eimeria life cycle.Support for the chicken: Reduction of oocyst shedding
Leaf powder of Artemisia annua
Support for the chicken: Protection from pathological symptoms and mortality associated with Eimeria tenella infection. Reduced lesion score and fecal oocyst output.
The leaf powder was more efficient than the essential oil, which could be due to a lack of Artemisinin in the oil, and to the greater antioxidant ability of A. annua leaves than the oil.
Phenols
Inhibition of coccidia:
Phenols change the cytoplasmic membrane’s permeability for cations (H+ and K+), impairing essential processes in the cell. The resulting leakage of cellular constituents leads to water unbalance, collapse of the membrane potential, inhibition of ATP synthesis, and, finally, cell death. Due to their toxic effect on the upper layer of mature enterocytes of the intestinal mucosa, they accelerate the natural renewal process, and, therefore, sporozoite-infected cells are shed before the coccidia reaches the merozoite phase.
Table 1: Bioactive compounds and their anticoccidial effect exerted in poultry
Consumers vote for natural – phytochemicals are the solution
Due to still rising antimicrobial resistance, consumers push for meat production without antimicrobial usage. Phytomolecules, as a natural solution, create opportunities to make poultry production more sustainable by reducing dependency on harmful drugs. With their advent, there is hope that antibiotic resistance can be held in check without affecting the profitability of poultry farming.
Toxin Mitigation 101: Essentials for Animal Production
By Monish Raj, Assistant Manager-Technical Services, EW Nutrition Inge Heinzl, Editor, EW Nutrition
Mycotoxins, toxic secondary metabolites produced by fungi, are a constant and severe threat to animal production. They can contaminate grains used for animal feed and are highly stable, invisible, and resistant to high temperatures and normal feed manufacturing processes. Mycotoxin-producing fungi can be found during plant growth and in stored grains; the prevalence of fungi species depends on environmental conditions, though in grains, we find mainly three genera: Aspergillus, Penicillium, and Fusarium. The most critical mycotoxins for poultry production and the fungi that produce them are detailed in Fig 1.
Figure 1: Fungi species and their mycotoxins of worldwide importance for poultry production (adapted from Bryden, 2012).
The effects of mycotoxins on the animal are manifold
When, usually, more than one mycotoxin enters the animal, they “cooperate” with each other, which means that they combine their effects in different ways. Also, not all mycotoxins have the same targets.
The synergistic effect: When 1+1 ≥3
Even at low concentrations, mycotoxins can display synergistic effects, which means that the toxicological consequences of two or more mycotoxins present in the same sample will be higher than the sum of the toxicological effects of the individual mycotoxins. So, disregarded mycotoxins can suddenly get important due to their additive or synergistic effect.
Table 1: Synergistic effects of mycotoxins in poultry
Synergistic interactions
DON
ZEN
T-2
DAS
FUM
*
*
*
NIV
*
*
*
AFL
*
*
Table 2: Additive effects of mycotoxins in poultry
Additive interactions
AFL
T2
DAS
MON
FUM
+
+
+
+
DON
+
+
OTA
+
+
Recognize the effects of mycotoxins in animals is not easy
The mode of action of mycotoxins in animals is complex and has many implications. Research so far could identify the main target organs and effects of high levels of individual mycotoxins. However, the impact of low contamination levels and interactions are not entirely understood, as they are subtle, and their identification requires diverse analytical methods and closer observation.
With regard to the gastrointestinal tract, mycotoxins can inhibit the absorption of nutrients vital for maintaining health, growth, productivity, and reproduction. The nutrients affected include amino acids, lipid-soluble vitamins (vitamins A, D, E, and K), and minerals, especially Ca and P (Devegowda and Murthy, 2005). As a result of improper absorption of nutrients, egg production, eggshell formation, fertility, and hatchability are also negatively influenced.
Most mycotoxins also have a negative impact on the immune system, causing a higher susceptibility to disease and compromising the success of vaccinations. Besides that, organs like kidneys, the liver, and lungs, but also reproduction, endocrine, and nervous systems get battered.
Mycotoxins have specific targets
Aflatoxins, fumonisins, and ochratoxin impair the liver and thus the physiological processes modulated and performed by it:
lipid and carbohydrate metabolism and storage
synthesis of functional proteins such as hormones, enzymes, and nutrient transporters
metabolism of proteins, vitamins, and minerals.
For trichothecenes, the gastrointestinal tract is the main target. There, they hamper digestion, absorption, and intestinal integrity. T-2 can even produce necrosis in the oral cavity and esophagus.
Figure 2: Main target organs of important mycotoxins
How to reduce mycotoxicosis?
There are two main paths of action, depending on whether you are placed along the crop production, feed production, or animal production cycle. Essentially, you can either prevent the formation of mycotoxins on the plant on the field during harvest and storage or, if placed at a further point along the chain, mitigate their impact.
Preventing mycotoxin production means preventing mold growth
To minimize the production of mycotoxins, the development of molds must be inhibited already during the cultivation of the plants and later on throughout storage. For this purpose, different measures can be taken:
Selection of the suitable crop variety, good practices, and optimal harvesting conditions are half of the battle
Already before and during the production of the grains, actions can be taken to minimize mold growth as far as possible:
Choose varieties of grain that are area-specific and resistant to insects and fungal attacks.
Practice crop rotation
Harvest proper and timely
Avoid damage to kernels by maintaining the proper condition of harvesting equipment.
Optimal moisture of the grains and the best hygienic conditions are essential
The next step is storage. Here too, try to provide the best conditions.
Dry properly: grains should be stored at <13% of moisture
Control moisture: minimize chances of moisture to increase due to condensation, and rain-water leakage
Biosecurity: clean the bins and silos routinely.
Prevent mold growth: organic acids can help prevent mold growth and increase storage life.
Mold production does not mean that the war is lost
Even if molds and, therefore, mycotoxins occur, there is still the possibility to change tack with several actions. There are measures to improve feed and support the animal when it has already ingested the contaminated feed.
1. Feed can sometimes be decontaminated
If a high level of mycotoxin contamination is detected, removing, replacing, or diluting contaminated raw materials is possible. However, this is not very practical, economically costly, and not always very effective, as many molds cannot be seen. Also, heat treatment does not have the desired effect, as mycotoxins are highly heat stable.
2. Effects of mycotoxins can be mitigated
Even when mycotoxins are already present in raw materials or finished feed, you still can act. Adding products adsorbing the mycotoxins or mitigating the effects of mycotoxins in the organism has been considered a highly-effective measure to protect the animals (Galvano et al., 2001).
This type of mycotoxin mitigation happens at the animal production stage and consists of suppressing or reducing the absorption of mycotoxins in the animal. Suppose the mycotoxins get absorbed in the animal to a certain degree. In that case, mycotoxin mitigation agents help by promoting the excretion of mycotoxins, modifying their mode of action, or reducing their effects. As toxin-mitigating agents, the following are very common:
Aluminosilicates: inorganic compounds widely found in nature that are the most common agents used to mitigate the impact of mycotoxins in animals. Their layered (phyllosilicates) or porous (tectosilicates) structure helps “trap” mycotoxins and adsorbs them.
Bentonite / Montmorillonite: classified as phyllosilicate, originated from volcanic ash. This absorbent clay is known to bind multiple toxins in vivo. Incidentally, its name derives from the Benton Shale in the USA, where large formations were discovered 150 years ago.
Bentonite mainly consists of smectite minerals, especially montmorillonite (a layered silicate with a larger surface area and laminar structure).
Zeolites: porous crystalline tectosilicates, consisting of aluminum, oxygen, and silicon. They have a framework structure with channels that fit cations and small molecules. The name “zeolite” means “boiling stone” in Greek, alluding to the steam this type of mineral can give off in the heat). The large pores of this material help to trap toxins.
Activated charcoal: the charcoal is “activated” when heated at very high temperatures together with gas. Afterward, it is submitted to chemical processes to remove impurities and expand the surface area. This porous, powdered, non-soluble organic compound is sometimes used as a binder, including in cases of treating acute poisoning with certain substances.
Yeast cell wall: derived from Saccharomyces cerevisiae. Yeast cell walls are widely used as adsorbing agents. Esterified glucomannan polymer extracted from the yeast cell wall was shown to bind to aflatoxin, ochratoxin, and T-2 toxin, individually and combined (Raju and Devegowda 2000).
Bacteria: In some studies, Lactic Acid Bacteria (LAB), particularly Lactobacillus rhamnosus, were found to have the ability to reduce mycotoxin contamination.
Which characteristics are crucial for an effective toxin-mitigating solution
If you are looking for an effective solution to mitigate the adverse effects of mycotoxins, you should keep some essential requirements:
The product must be safe to use:
safe for the feed-mill workers.
does not have any adverse effect on the animal
does not leave residues in the animal
does not bind with nutrients in the feed.
It must show the following effects:
effectively adsorbs the toxins relevant to your operation.
helps the animals to cope with the consequences of non-bound toxins.
It must be practical to use:
cost-effective
easy to store and add to the feed.
Depending on
the challenge (one mycotoxin or several, aflatoxin or another mycotoxin),
the animals (short-cycle or long-living animals), and
the economical resources that can be invested,
different solutions are available on the market. The more cost-effective solutions mainly contain clay to adsorb the toxins. Higher-in-price products often additionally contain substances such as phytogenics supporting the animal to cope with the consequences of non-bound mycotoxins.
Solis – the cost-effective solution
In the case of contamination with only aflatoxin, the cost-effective solution Solis is recommended. Solis consists of well-selected superior silicates with high surface area due to its layered structure. Solis shows high adsorption of aflatoxin B1, which was proven in a trial:
Figure 3: Binding capacity of Solis for Aflatoxin
Even at a low inclusion rate, Solis effectively binds the tested mycotoxin at a very high rate of nearly 100%. It is a high-efficient, cost-effective solution for aflatoxin contamination.
Solis Max 2.0: The effective mycotoxin solution for sustainable profitability
Solis Max 2.0 has a synergistic combination of ingredients that acts by chemi- and physisorption to prevent toxic fungal metabolites from damaging the animal’s gastrointestinal tract and entering the bloodstream.
Figure 4: Composition and effects of Solis Max 2.0
Solis Max 2.0 is suitable for more complex challenges and longer-living animals: in addition to the pure mycotoxin adsorption, Solis Max 2.0 also effectively supports the liver and, thus, the animal in its fight against mycotoxins.
In an in vitro trial, the adsorption capacity of Solis Max 2.0 for the most relevant mycotoxins was tested. For the test, the concentrations of Solis Max 2.0 in the test solutions equated to 1kg/t and 2kg/t of feed.
Figure 5: Efficacy of Solis Max 2.0 against different mycotoxins relevant in poultry production
The test showed a high adsorption capacity: between 80% and 90% for Aflatoxin B1, T-2 Toxin (2kg/t), and Fumonisin B1. For OTA, DON, and Zearalenone, adsorption rates between 40% and 80% could be achieved at both concentrations (Figure 5). This test demonstrated that Solis Max 2.0 could be considered a valuable tool to mitigate the effects of mycotoxins in poultry.
Broiler trial shows improved performance in broilers
Protected and, therefore, healthier animals can use their resources for growing/laying eggs. A trial showed improved liver health and performance in broilers challenged with two different mycotoxins but supported with Solis Max 2.0.
For the trial, 480 Ross-308 broilers were divided into three groups of 160 birds each. Each group was placed in 8 pens of 20 birds in a single house. Nutrition and management were the same for all groups. If the birds were challenged, they received feed contaminated with 30 ppb of Aflatoxin B1 (AFB1) and 500 ppb of Ochratoxin Alpha (OTA).
Negative control:
no challenge
no mycotoxin-mitigating product
Challenged group:
challenge
no mycotoxin-mitigating product
Challenge + Solis Max 2.0
challenge
Solis Max 2.0, 1kg/t
The body weight and FCR performance parameters were measured, as well as the blood parameters of alanine aminotransferase and aspartate aminotransferase, both related to liver damage when increased.
Concerning performance as well as liver health, the trial showed partly even better results for the challenged group fed with Solis Max 2.0 than for the negative, unchallenged control (Figures 6 and 7):
6% higher body weight than the negative control and 18.5% higher body weight than the challenged group
12 points and 49 points better FCR than the negative control and the challenged group, respectively
Lower levels of AST and ALT compared to the challenged group, showing a better liver health
The values for body weight, FCR, and AST, even better than the negative control, may be owed to the content of different gut and liver health-supporting phytomolecules.
Figure 6: Better performance data due to the addition of Solis Max 2.0
Figure 7: Healthier liver shown by lower values of AST and ALT
Effective toxin risk management: staying power is required
Mycotoxin mitigation requires many different approaches. Mycotoxin mitigation starts with sewing the appropriate plants and continues up to the post-ingestion moment. From various studies and field experience, we find that besides the right decisions about grain crops, storage management, and hygiene, the use of effective products which mitigate the adverse effects of mycotoxins is the most practical and effective way to maintain animals healthy and well-performing. According to Eskola and co-workers (2020), the worldwide contamination of crops with mycotoxins can be up to 80% due to the impact of climate change and the availability of sensitive technologies for analysis and detection. Using a proper mycotoxin mitigation program as a precautionary measure is, therefore, always recommended in animal production.
EW Nutrition’s Toxin Risk Management Program supports farmers by offering a tool (MasterRisk) that helps identify and evaluate the risk and gives recommendations concerning using toxin solutions.