Climate change in poultry production: 5 major threats and what you can do to mitigate the impact

“Every single social and global issue of our day is a business opportunity in disguise.”
Peter Drucker
By Ajay Bhoyar, Global Technical Manager, EW Nutrition
Topics covered
-
- Major areas impacted by climate change
- Feed quality
- Genetics
- Farm management
- Animal performance
- On- and off-farm logistics
The cost of doing nothing
Global livestock systems constitute an industrial asset worth over $1.4 trillion. Projections indicate that the global livestock population, now at 60+ billion, could exceed 100 billion by 2050 – more than ten times the expected human population at that time (Yitbarek 2019, Herrero 2009).
Our industry bears an enormous responsibility: to feed the growing population, sustainably and consistently, despite increasing challenges. And one of the biggest challenges is already looming large.
Animal agriculture, including poultry farming, is particularly susceptible to the adverse effects of climate change. Increased extreme weather events, farm fires facilitated by drought, thermal pressure on farmed animals, reduced availability or increased prices of water, raw materials, and electricity, and much more are already impacting the industry.
This is, in all likelihood, just the beginning. How exactly will poultry production be affected in the future – and what can you do to future-proof your operation against the coming challenges?
Major impact areas of climate change – and what to do about them
1. Feed quality
Excessive heat, droughts, or floods can reduce crop yields, decrease nutritional content, and increase the risk of pests, pathogens, and weed outbreaks.
In 2020, 75% of soil in Mexico was declared too dry to cultivate crops. In 2021, 70% of the country was impacted by crop loss and water shortages caused by drought. Corn yield decreased by 18% in five years and is expected to fall further (Carlin 2023).
Plants with a C3 photosynthetic pathway such as wheat, rice, or soybean can benefit from increased temperature more than the so-called C4 plants such as corn or sorghum (Cui 2021). NASA projections show corn crop yields are expected to decline 24% in the next 30 years (Gray 2021).
Moreover, increased temperature, shifts in rainfall patterns, and elevated surface greenhouse gas (GHG) concentrations can also lead to lower grain protein concentration (Godde 2010, Myers 2014), as well as affect mineral and vitamin concentrations in plants.
Pollinator-dependent crops like soybean or rapeseed could also see decreased yield under climactic challenges (Godde 2020).
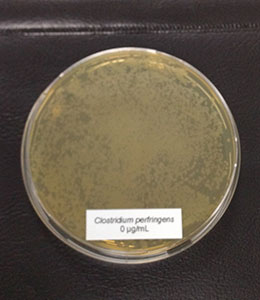
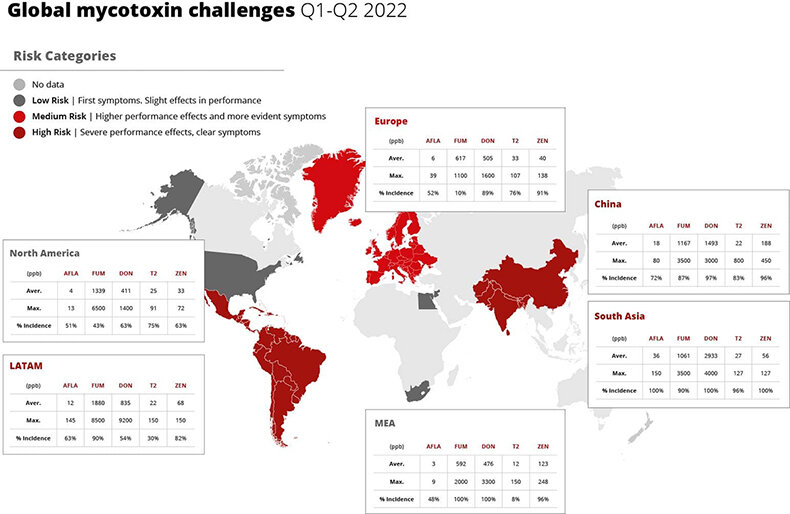
Warmer temperatures and changes in precipitation patterns can create favorable conditions for the growth of mycotoxins, leading to reduced feed quality and health problems in poultry. Especially corn and sorghum are vulnerable to aflatoxin contamination in hot and humid conditions. On top of this, storage will become more challenging as pathogen growth will further erode feed quality.
ACTION
- Diversification of feed sources: Exploring alternative feed ingredients that are less reliant on climate-sensitive crops can help mitigate the impact of changing weather patterns on feed availability and costs.
- Mycotoxin mitigation: Not all toxin mitigation solutions are created equal. Choose standardized toxins mitigation solutions based on their efficacy instead of upfront cost. The products that are regularly tested against undesirable and harmful impurities like dioxins, dioxins-like PCBs and heavy metals.
2. Genetics

Rising temperatures may lead to reduced fertility and hatchability, affecting the overall health and reproductive performance of chickens. Extreme heat can also impact the expression of genes related to growth, feed efficiency, and resistance to diseases. As a result, poultry breeders and geneticists face the challenge of developing more heat-tolerant poultry breeds to ensure sustainable production under changing climatic conditions.
ACTION
- Genetic selection for thermotolerance: Breeding programs can focus on developing more heat-tolerant chicken breeds that exhibit improved performance and resilience in challenging climatic conditions. Producers need to pay attention to the specifics of the breed’s genetic makeup.
3. Farm Management
3.1 Solving for thermal comfort: Electricity costs
The thermal comfort of livestock is no longer a concern for tropical zones only. Temperate zones are also seeing sustained increases in ambient temperatures.
High temperatures and prolonged heat waves increase electricity consumption as farmers rely on ventilation, cooling systems, and artificial lighting to maintain optimal conditions for chickens. Consequently, energy costs will rise, impacting the profitability of poultry farms.
3.2 Solving for water availability: Resource management
Water scarcity, changing precipitation patterns, and droughts can limit the availability of water resources, affecting poultry farms’ water consumption and overall operational efficiency.
The quality of water is also an increasing concern. The UN states that “higher water temperatures and more frequent floods and droughts are projected to exacerbate many forms of water pollution – from sediments to pathogens and pesticides”. Reduced raw water quality “can decrease animal water intake, feed intake and health” (Valente-Campos 2019). Especially in Asia and Africa, which have seen massive increases in floods and droughts, respectively, water scarcity and quality will pose severe issues.
ACTION
- Improved farm management practices: Implementing energy-efficient systems, such as solar power and energy-saving technologies, can reduce electricity consumption and associated costs. Water management techniques, such as rainwater harvesting and efficient irrigation systems, can help mitigate the impact of water scarcity. As always, strict biosecurity will play a critical role.
- Enhanced ventilation and cooling systems: Upgrading ventilation systems and implementing efficient cooling mechanisms can alleviate heat stress on chickens, enhancing their overall health and productivity. Regular maintenance and sensor technologies also play an important preventive role.
3.3 Built-up and human capital risk
In high-risk areas, machinery, electricity networks, telecommunications, building infrastructure in general can be impacted by extreme weather events, rising sea levels etc. (Nardone 2010).
Labor availability and productivity might, on the other hand, be impacted in many areas. Disease outbreaks, including new strains, as well as decreased air quality, extreme events etc. might in the future contribute to labor shortages. The number of unsafe hot workdays is expected to double by 2050, which will impact especially rural India, sub-Saharan Africa, and Southeast Asia (Carlin 2023).
ACTION
- Climate-resilient infrastructure: Investing in resilient infrastructure, such as elevated coops, flood-resistant buildings, or disease surveillance technology can minimize the risk of incidents from weather events and can support early action against disease pressure. Investments in smart farming can also relieve pressure on labor and improve speed of action.
- Insurability and loan math: Any future-looking business needs to work with the likelihood of increased insurance costs and higher insurability requirements. Also, a point will come at which non-resilient infrastructure will not be financed.
4. Animal performance

Heat stress reduces productivity, impacts fertility, and increases susceptibility to disease. It can also reduce the size of eggs and thickness of eggshells (Godde 2021)
While colder areas will benefit from reduced house heating and ventilation needs, warm areas will be at increased risk. A hot environment “impairs production (growth, meat and milk yield and quality, egg yield, weight, and quality) and reproductive performance, metabolic and health status, and immune response” (Nardone 2010, Ali 2020).
The proliferation of pathogens in warm environments will pose further challenges. Antibiotic resistance from attempts to control these issues will only compound the problem.
Additionally, as mentioned before, changes in weather patterns can impact crop yields, including the availability and affordability of feed ingredients for chickens. Producers will have to reformulate often to match availability, cost, and nutritional value.
ACTION
- Stress and pathogenic impact mitigation solutions: Phytogenic feed additives can support poultry gut health and strengthen the immune response when confronted with stress factors, including heat stress, humid environments, pen density, and pathogen pressure. With the added benefit of reducing dependence on antibiotics and other medication, they can naturally stimulate or support a healthy response to challenges.
5. On- and off-farm logistics
Transportation is also affected all along the supply chain, from bringing feed or young stock to the farm to moving livestock to processing facilities and further distribution along the chain. Extreme weather events, such as hurricanes, floods, or heavy snowfall, can lead to power outages and/or disrupt transportation routes and infrastructure, hindering the timely delivery of chicks, feed, and other essential supplies to poultry farms.
In addition to the challenge of transportation, packaging will soon fall under regulatory scrutiny. Sustainability requirements may be national, but compliance will have to follow across borders for any producers eyeing international markets.
ACTION
- Data is your friend: Transportation and logistics data can helps improve efficiency and reduce your environmental impact. Start tracking fuel consumption, carbon emissions, transportation costs, and other relevant metrics to identify areas for optimization.
- Think globally: ESG (Environmental, Social and Governance) guidance will become a standard in many important markets, including Europe and the US. Keep an eye on international regulations, especially for your target markets. Their ESG requirements are your ESG requirements.
The world needs more meat
The bad news is that climate change is coming at us fast. Animal agriculture will be among the most heavily impacted. Major adjustments will be needed to mitigate the effects and to embrace the long view.
1.5% annual growth in livestock and fish production will result from improvements in per-animal productivity. Poultry will account for over 50% of meat production growth, due to sustained profitability and favorable meat-to-feed price ratio (OECD FAO 2022).
The good news is that livestock systems remain critical to our growing population. The world population is projected to grow to 9.8 billion by 2050 (UNDESA, 2017). Livestock products (meat, milk and eggs) account for about 30% of the population’s protein supply, with large regional variations (FAOSTAT, 2022; Godde et al, 2021).
To answer this growing demand, world meat production is expected to increase by 14% by the end of the decade, compared to current figures (Carlin 2023). The increase in meat demand might be as high as 76% compared to 2005/2007 (Alexandratos 2012).
The cost of doing nothing
We must look at the challenges of climate change, in the words of Peter Drucker, as a business opportunity. As always, those who act early will reap important rewards – not just through market differentiation but through economic resilience.
What awaits those who do not take action?
The United Nations Environment Programme warns of some foreseeable consequences of inaction, most of which can be grouped under three categories:
- Rising costs: Cost of decreased performance, increased cost of doing business, carbon taxes
- Policy restrictions: Once a few major markets have implemented restrictive labeling, packaging, or production regulations, anyone who wants to operate in these markets is subject to the same restrictions.
- Reputational risk / Market and investor preferences: The risk of falling behind or not taking action, in other words the opportunity cost, is hard to quantify until it’s too late. Banks and investors may give up on unsustainable financing as soon as consumers and/or regulators show signs of concern. Acting ahead of the curve is also a market positioning win as well as economic win. The market rewards first movers.
The impact of climate change on genetics, farm management, animal performance, farm logistics, and transportation necessitate proactive adaptation and mitigation strategies, in coordination with local and global expertise. Responses will vary depending on geography, production type, and more – but doing nothing is no longer an option. By implementing sustainable practices across the board and investing in resilient infrastructure, poultry producers can maintain a robust, high-performing, sustainable production system.
References
Alexandratos, N. and Jelle Bruinsma. “World agriculture towards 2030/2050: the 2012 revision”. ESA Working Paper No. 12-03, June 2012. https://www.fao.org/3/ap106e/ap106e.pdf
Ali, Zulfekar et al. “Impact of global climate change on livestock health: Bangladesh perspective”. Open Veterinary Journal. 2020 Apr-Jun; 10(2): 178–188. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7419064/
Bernabucci, Umberto. “Climate change: impact on livestock and how can we adapt”. Animal Frontiers, Volume 9, Issue 1, January 2019, Pages 3–5, https://doi.org/10.1093/af/vfy039
Cheng, M. et al. Climate Change and Livestock Production: A Literature Review. Atmosphere 2022, 13(1), 140; https://doi.org/10.3390/atmos13010140
Carlin, David et al. Climate Risks in the Agriculture Sector. UN Environment Programme, March 2023. https://www.unepfi.org/wordpress/wp-content/uploads/2023/03/Agriculture-Sector-Risks-Briefing.pdf
Cui, Hongchang. “Challenges and Approaches to Crop Improvement Through C3-to-C4 Engineering.” Frontiers in Plant Science, 14 September 2021, Volume 12 – 2021. https://doi.org/10.3389/fpls.2021.715391
FAO Statistics. Statistical yearbook world food and agriculture. 2022. https://www.fao.org/3/cc2211en/cc2211en.pdf
Godde, C.M. et al. “Impacts of climate change on the livestock food supply chain; a review of the evidence”. Global Food Security, 2021 Mar; 28: 100488. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7938222/
Gray, Ellen. “Global Climate Change Impact on Crops Expected Within 10 Years, NASA Study Finds”. NASA Global Climate Change. November 2, 2021. https://climate.nasa.gov/news/3124/global-climate-change-impact-on-crops-expected-within-10-years-nasa-study-finds/
Herrero, Mario et al. “Livestock, livelihoods and the environment: understanding the trade-offs. Current Opinion in Environmental Sustainability Volume 1, Issue 2, December 2009, Pages 111-120. https://doi.org/10.1016/j.cosust.2009.10.003
Nardone, A. et al. “Effects of climate changes on animal production and sustainability of livestock systems”. Livestock Science, Volume 130, Issues 1–3, May 2010, Pages 57-69. https://www.sciencedirect.com/science/article/abs/pii/S1871141310000740
OECD FAO. Agricultural Outlook 2022-2031. https://www.oecd.org/development/oecd-fao-agricultural-outlook-19991142.htm
United Nations Climate Action. Water – at the center of the climate crisis. Retrieved 20 June 2023. https://www.un.org/en/climatechange/science/climate-issues/water#:~:text=Water%20quality%20is%20also%20affected,pathogens%20and%20pesticides%20(IPCC).
United Nations Department of Economic and Social Affairs (UNDESA). “World population projected to reach 9.8 billion in 2050, and 11.2 billion in 2100”. 2017 Revision of World Population Prospects, 21 June 2017. https://www.un.org/development/desa/en/news/population/world-population-prospects-2017.html#:~:text=News-,World%20population%20projected%20to%20reach%209.8%20billion%20in,and%2011.2%20billion%20in%202100&text=The%20current%20world%20population%20of,Nations%20report%20being%20launched%20today.
USDA. Climate Change and Agriculture in the United States: Effects and Adaptation. Technical Bulletin 1935, February 2013. Retrieved June 2023. https://www.climatehubs.usda.gov/animal-agriculture-changing-climate#:~:text=Breadcrumb&text=Climate%20change%20may%20affect%20animal,and%20disease%20and%20pest%20distributions.
Valente-Campos S., et al. “Critical issues and alternatives for the establishment of chemical water quality criteria for livestock”. Regul. Toxicol. Pharmacol. 2019;104:108–114. doi: 10.1016/j.yrtph.2019.03.003
Yitbarek, Melkamu Bezabih. “Livestock and livestock product trends by 2050: Review”. International Journal of Animal Research, 2019; 4:30. https://www.researchgate.net/publication/344188926_Livestock_and_Livestock_products_by_2050