Environmental Stress and Mycotoxins in Breeders: Hidden Losses in Fertility, Egg Quality & Chick Output
Author: Dr. Vaibhav Gawande, Feed Safety and Toxin Control Specialist – South Asia, EW Nutrition
In Tropical countries like India, Bangladesh & Sri Lanka, high temperatures and humidity significantly increase the risk of mycotoxin contamination in animal feed. FAO surveys report mycotoxin contamination in over 70% of cereals and oilseeds used for animal feed, making them a major threat to poultry health and productivity.
The problem becomes particularly severe during the summer & monsoon when:
The new maize crop with high moisture enters the market
Drying is often inadequate
Feed mill humidity remains high
The Science of Mycotoxins & Environmental Risk
Summer temperatures (28–38°C) and high humidity promote fungal growth and mycotoxin production in feed ingredients.
Freshly harvested “new maize” often enters feed mills with unsafe moisture levels (13–15%), exceeding the safe storage limit of 11–12%.
High-moisture grain undergoes self-heating during storage, creating ideal conditions for fungal proliferation.
Poor aeration, condensation, insect damage, and humid storage environments further accelerate contamination.
Common summer mycotoxins include Aflatoxins, Ochratoxins, and T-2 toxins, which may develop during both pre- and post-harvest stages.
Although pelleting and heat treatment may destroy molds, mycotoxins are generally heat-stable and remain toxic to poultry.
Regional Case Studies: South Asia
India
Coastal Humidity Crisis – Andhra Pradesh, Telangana, Tamil Nadu, Odisha & West Bengal
Pre-monsoon humidity promotes Aspergillus flavus growth in stored maize.
Aflatoxin contamination commonly reduces shell quality and egg production.
Fatty Liver Hemorrhagic Syndrome (FLHS) is frequently observed.
Short-Storage Trap – Punjab & Haryana
Wet maize stored during summer heat retains internal moisture, favoring T-2 toxin and Ochratoxin formation.
Broilers commonly show oral lesions, feed refusal, and poor FCR.
Mixed Toxin Challenge – Maharashtra
Co-contamination with Aflatoxin and Fumonisin is common.
Combined toxicity causes immunosuppression, poor vaccine response, and increased mortality during disease outbreaks.
Bangladesh
High Humidity Feed Risk
Persistent humidity and post-flood harvesting increase moisture retention in maize and rice by-products.
Aflatoxin frequently co-occurs with Ochratoxin A.
Causes immunosuppression, uneven flock uniformity, and poor hatchability in breeders.
Sri Lanka
Tropical Storage Challenge
Tropical coastal humidity and prolonged ingredient storage favor fungal proliferation.
Aflatoxin and Fumonisin commonly develop during humid transit and storage.
Causes thin shells, liver damage, and poor FCR.
Nepal
Mountain Moisture Variability
Humid Terai grains stored in cool hill regions favor mixed mycotoxin contamination.
Aflatoxins commonly co-occur with DON and Zearalenone.
DON causes feed refusal, while Zearalenone induces prolapse and false layers.
Mycotoxins and High Temperature Humidity Index (THI): Synergistic effects on poultry health, immunity & productivity
The Immunological “Blackout”
Aflatoxins, Trichothecenes, and Ochratoxins inhibit protein synthesis, reducing the formation of antibodies and immune cells.
Mycotoxins cause atrophy of immune organs (bursa of Fabricius, thymus, and spleen)
Macrophage activity and phagocytosis are reduced, weakening bacterial clearance.
Cytokine signaling is disrupted, delaying immune activation against infections.
Enhanced oxidative stress: Mycotoxins increase the occurrence of reactive oxygen species (ROS), and heat stress weakens antioxidant defenses, resulting in severe cellular and liver damage.
Oxidative stress caused by aflatoxins and trichothecenes leads to immune cell apoptosis and tissue damage.
Gut Health & Barrier Damage
The gut is the first line of defense. Mycotoxins and heat stress act like a “chemical and physical abrasive” on the intestinal lining.
Villi Destruction: T-2 and Aflatoxins cause necrosis (cell death) of the intestinal villi. This reduces the surface area for nutrient absorption, leading to poor FCR.
The “Leaky Gut” Phenomenon: Heat stress causes blood to be diverted from the internal organs to the skin for cooling (vasodilation). As a result, the gut receives less oxygen, causing the tight junctions (the “glue” between intestinal cells) to break down and become permeable. Mycotoxins also have a direct effect, inhibiting tight junctions proteins.
Pathogen Entry: Mycotoxins further erode the protective mucus layer. With the “gates” (tight junctions) open and the “walls” (mucus) gone, bacteria can freely enter the bloodstream.
Disease Susceptibility
Because the immune system is “blind” and the gut is “leaky,” the bird becomes a target for opportunistic infections.
Secondary Bacterial Infections: Normal gut bacteria like E. coli and Salmonella transition from harmless to fatal, causing systemic septicemia.
Viral Synergism: Small viral loads, such as Inclusion Body Hepatitis (IBH) that a healthy bird would normally survive, become highly fatal.
Coccidiosis Flare-ups: Damaged gut linings are more easily colonized by Eimeria, making standard anti-coccidial programs less effective.
Vaccine Failure:
Mycotoxins suppress B-cell and T-cell maturation, reducing vaccine effectiveness.
Low immunoglobulin (IgG, IgA, IgM) production results in poor antibody titers.
Maternal toxin exposure reduces immunity transfer to chicks, increasing early-age disease vulnerability.
Figure 1: How mycotoxins and heat stress cause damage in poultry
Breeder Reproductive Dysfunction and Transgenerational Effects of Mycotoxins
In breeder operations, mycotoxins represent a catastrophic economic threat because they are vertically transmitted. Unlike commercial layers, where the loss is limited to the individual bird’s production, breeder contamination compromises the viability of the entire next generation.
1. Impact on the Reproductive Systems (Male & Female)
Mycotoxins hit both sides of the fertility equation, often exacerbated by summer heat.
Female Reproductive System
Mode of action: Mycotoxins (especially Zearalenone) mimic estrogen. This disrupts the hypothalamic-pituitary-ovarian axis.
Impact: inflammation of the oviduct, cystic ovaries, and reduced synthesis of yolk precursors in the liver, resulting in a sharp drop in egg production and poor internal egg quality.
Male Fertility
Mode of action: Toxins like T-2 and Aflatoxin induce oxidative stress that damages the phospholipid membrane of sperm cells.
Impact: Under heat stress, rooster semen quality already declines; mycotoxins accelerate this by reducing sperm motility, concentration, and increasing morphological abnormalities. This leads to a massive spike in infertility rates.
2. Hatchability & Embryonic Mortality
For breeders, mycotoxins represent a “generational loss” via vertical transmission.
Mode of action (the yolk bridge): Some mycotoxins are highly lipophilic. As the liver assembles the yolk, it deposits toxins directly into the egg.
The “three-wave” mortality:
Early (Days 1–7): Toxins interfere with mitosis (cell division), leading to early deaths often mistaken for “infertility.”
Mid-Term (Days 8–18): As the embryo begins intensive absorption of the toxic yolk, its developing liver and kidneys are compromised. This is the classic “Toxin Fingerprint.”
Late (Days 19–21): Ochratoxins impair the embryo’s ability to mobilize calcium from the eggshell. As a result, the chick becomes too weak to pip and dies fully developed inside the shell (“dead-in-shell”).
3. Chick Quality, Grading, and Settability
The “Chick Quality” starts in the breeder’s gut and kidney health.
Mode of action (nutrient malabsorption): Mycotoxins reduce pancreatic lipase and bile salts. This prevents the mother from absorbing fat-soluble vitamins (A, D, E, K) and pigments. They also lead to lower intestinal adsorption due to a reduced absorption area and lower transporter efficacy.
Impact on chick quality:
“Pale Bird Syndrome”: Chicks lack vital carotenoids for early-stage defense.
Skeletal weakness: Interference with Vitamin D3 metabolism results in weak legs and “rubbery beaks” in Day-Old-Chicks (DOCs).
High first week mortality (FWM): Chicks hatch immunosuppressed, leading to high mortality during the first week.
Impact on egg grading & settability: Mycotoxins (Ochratoxin) are nephrotoxic, damaging the kidneys and disrupting the blood calcium-carbonate balance. This leads to “Sandpaper” shells, misshapen eggs, and a 5–10% drop in the number of settable eggs fit for the incubator.
Figure 2: The impact of mycotoxins on breeder production and economics
Pro tip for breeders: In summers, high-moisture new maize triggers a mycotoxin surge that synergistically destroys the breeder’s kidneys and shell gland, crippling egg settability. A 5% spike in “Dead-in-Shell” embryos during breakout analysis is a definitive indicator of feed toxicity rather than incubator failure.
Integrated Mycotoxin Mitigation Strategies for Poultry Production
To pursue an effective mycotoxin mitigation strategy, it is essential first to identify which mycotoxins are relevant to a given region before implementing measures.
Strict moisture control: Reject any maize arriving with >14% moisture.
Rapid screening: Perform rapid mycotoxin screening before unloading raw materials.
Mechanical grain driers: To maintain a safe storage moisture level (<12%)
2. Feed Plant & Storage Hygiene
First-In, First-Out (FIFO): Ensure strict inventory rotation to prevent “pockets” of old, moldy feed from contaminating new batches.
Frequently clean silos and elevators: High temperature and humidity cause moisture condensation on silo walls, leading to localized mold growth.
Antifungal Treatment: Use buffered organic acids (propionic and formic acid) to limit mold proliferation in feed.
3. Broad-Spectrum binders:
Bentonites (for Aflatoxins) and Yeast Cell Walls: These components help bind pathogenic bacteria like E. coli that capitalize on the “leaky gut” caused by toxins and heat stress.
4. Physiological & Gut Health Support
Water acidification: Lower the drinking water pH to 4.5–5.5. This prevents bacterial blooms in the water lines when birds increase water intake by 3 times during heat stress.
Liver & kidney tonics: Supplemental hepatic (milk thistle/silymarin) and renal support to help the bird metabolize and export toxins more efficiently.
Metabolite supplementation: Use 25-hydroxyvitamin D3 in breeder diets to bypass the liver/kidney damage and ensure shell quality remains intact.
Antioxidant boost: Increase levels of Vitamin E, C, and Selenium to counter the oxidative stress caused by the heat-toxin synergy.
5. Monitoring & Diagnostics
Hatchery breakout analysis: Monitor “dead-in-shell” embryos. A spike in mid-term mortality is an immediate indicator that the breeder feed toxin binder needs a dosage increase.
Frequent lab testing: Mycotoxin testing at least weekly during the new maize transition to identify the specific toxin profile.
Solutions are available to support toxin risk management
In the challenging climate where high-moisture “new maize” and summer humidity create a complex cocktail of mycotoxins, endotoxins, and pesticide residues, traditional, single-ingredient binders often fall short. Modern poultry production requires a proactive solution that does more than just “bind”; it must protect the bird’s internal integrity.
Solis Max – The effective myco- and endotoxin solution for sustained profitability
Solis Max is engineered to meet customers’ demand for an effective solution, offering a multi-pronged defense mechanism that targets the root causes of performance collapse. Solis Max uses a synergistic blend of five key components to ensure the flock’s safety.
Trials prove the effectiveness of Solis Max
Solis Max shows dose-dependent adsorbing capacity against multiple mycotoxins:
Figure 4: Mycotoxin Binding Capacity Of Solis Max
SOLIS MAX shows endotoxin adsorbing capacity – 1mg of SOLIS MAX absorbs 20 endotoxin units (EU) of E. coli endotoxin (80% adsorption rate):
Figure 5: Endotoxin Binding Capacity Of Solis Max
Solis Max demonstrates high pesticide binding efficiency across multiple compounds:
Figure 6: Pesticide Net Binding Capacity Of Solis Max (%)
Conclusion:
The convergence of a high Temperature–Humidity Index (THI) and mycotoxicosis represents a critical, multisystem challenge in poultry production, precipitating severe pathology across the hepatic, renal, and gastrointestinal systems. In breeding operations, this crisis exhibits a transgenerational impact: lipophilic mycotoxins are vertically transmitted to the yolk, inducing mid-term embryonic mortality and compromising post-hatch progeny immunity.
Mitigation demands stringent control of raw material moisture alongside advanced, broad-spectrum interventions. Utilizing an advanced multi-pronged solution like Solis Max counters this synergy by providing physicochemical adsorption and targeted organ protection. By neutralizing the concurrent threats of mycotoxins, endotoxins, and pesticides, it preserves cellular integrity, mitigates systemic pathology, and maintains optimal performance under extreme environmental stress.
References available upon request.
Shifting Consumer Preferences in Dairy: The Higher Demand for High-Protein Milk in the GLP-1 Era
Author: Valentina Mayorga and Inge Heinzl, EW Nutrition
Today, the dairy market is undergoing a remarkable transformation. Demand is no longer focused solely on volume or fat content, but rather on a specific component: protein. But why is protein suddenly at the center of consumer attention? Recent estimates indicate that approximately one in eight adults (12%) in the United States is currently using a GLP-1 medication such as Ozempic or Wegovy for weight loss or chronic disease management (Lacsamana, 2025). By significantly reducing appetite and overall caloric intake, these medications may increase the risk of muscle mass loss in the absence of adequate nutritional planning, particularly when protein intake is insufficient.
Humans and animals compete for high-protein products
For this reason, consumers are increasingly seeking high-quality protein sources, particularly those rich in whey protein, known for its high biological value and rapid digestibility. However, this shift in consumer demand also creates a new challenge for the feed industry. Whey protein, traditionally used by feed mills as a highly digestible ingredient for young animals, is increasingly being diverted to human nutrition markets, creating direct competition for this valuable protein source. Either way, dairy consumption is growing, but not uniformly across all categories. The increase is concentrated in products with lower fat content and higher protein density, such as cottage cheese, premium Greek yogurt, and whey-protein-enriched milk beverages. The protein market is accelerating, and in an industry that rewards adaptation, standing still is simply another way of moving backward.
This shift in what consumers care about raises a key question for dairy farmers: how can they increase milk protein content through farm nutrition practices? Improving milk protein content isn’t just about putting more protein into the diet. It needs a balanced nutritional approach that supports rumen function, promotes microbial protein synthesis, and maintains overall metabolic function. With the right feed mix, farms can better meet consumers’ changing tastes.
Increase milk protein with higher energy intake
Milk protein synthesis is primarily driven by energy intake, particularly fermentable energy. When cows consume more metabolizable energy (ME), rumen microbial activity increases, leading to greater microbial protein synthesis. Since microbial protein represents the main source of metabolizable amino acids absorbed in the small intestine, improving rumen efficiency directly supports higher milk protein production. Research has shown that increasing concentrate intake is associated with increases in milk protein concentration, with a response of approximately +0.06 percentage units per additional 10 MJ of ME intake per day. This response occurs because higher energy intake increases dry matter intake, improves nitrogen utilization, enhances microbial growth, and ultimately increases the supply of metabolizable protein to the mammary gland. Importantly, the source of energy matters. Energy derived from fermentable carbohydrates, particularly starch and sugars, is far more effective at stimulating microbial protein synthesis than energy derived from fat.
Starch plays a crucial role
Among fermentable carbohydrates, starch plays a central role in increasing milk protein concentration. When starch in the diet increases, rumen fermentation produces more propionate. Propionate is absorbed and converted in the liver into glucose through gluconeogenesis. Glucose is essential for lactose synthesis in the mammary gland, and lactose regulates milk volume through osmotic pressure. At the same time, improved energy status enhances microbial protein synthesis, increasing the availability of amino acids for casein production. This makes increasing dietary starch one of the most influential nutritional strategies for enhancing milk protein concentration.
Replacing grass silage with forages higher in starch and sugars, such as maize silage or fodder beet, can increase total energy intake, milk yield, and milk protein concentration. However, starch must be carefully balanced with adequate fiber. Excessively low fiber levels can reduce rumen pH, leading to acidosis, decreased feed intake, milk fat depression, and compromised animal health. Therefore, the objective is not simply high starch inclusion but rather high fermentable energy within a stable rumen environment, supported by sufficient physically effective fiber.
Effective protein strategies coordinate the supply of fermentable energy and degradable protein
Feeding more crude protein alone does not increase milk protein concentration. If degradable protein exceeds the availability of fermentable energy, excess nitrogen is converted into urea, reducing nitrogen efficiency and increasing milk urea nitrogen (MUN) without improving milk protein yield. Instead, effective protein strategies involve synchronizing rumen-degradable protein (RDP) with fermentable carbohydrates to maximize microbial growth, while also providing adequate rumen-undegradable protein (RUP) to supply metabolizable amino acids directly to the intestine. Precision supplementation of limiting amino acids, particularly methionine between 2.4-2.5% and lysine between 7.2-7.5% of metabolizable protein (MP), ensures a crucial 3:1 ratio, supporting casein synthesis in the mammary gland and improving true milk protein yield.
Feeding is one thing, genetics is another
Under optimized nutritional management, realistic improvements in milk protein concentration can be achieved. In Holstein cows, which typically average around 3.1% protein, levels can increase to approximately 3.3–3.5%. In Jersey cows, which average around 3.9%, protein concentration may increase to approximately 4.1–4.3% with well-balanced diets and excellent management. Increases beyond these ranges generally require genetic selection in addition to nutritional adjustments.
Higher protein production is possible…up to a certain degree
High-starch diets often increase milk protein while potentially lowering milk fat percentage. This occurs because increased propionate production is associated with reduced acetate formation, and acetate is the primary precursor for milk fat synthesis. For consumers seeking dairy products with higher protein and lower fat content (particularly individuals aiming to preserve muscle mass while reducing caloric intake), this shift in milk composition may align with emerging market demands. However, excessive starch without adequate fiber can negatively impact rumen health, emphasizing the importance of nutritional balance.
References
Chamberlain, A. T., and J. M. Wilkinson. Feeding the dairy cow. Mountwood House: Chalcombe Publications, 2011.
The influence of moisture on salmonella control in feed processing
by Ivan Ilić, Application Manager EW Nutrition GmbH
Choosing the right strategy
During global client visits, I frequently observe that the primary objective of a process is disconnected from the subsequent steps and final actions. Choosing a strategy is sometimes done paradoxically – like putting worn-out winter tires on a vehicle just because they are cheap and available in your garage, and then attempting to race in the Paris-Dakar rally. To succeed, you must choose the right race or use the proper equipment; anything else is a waste of time and energy without meaningful results. Let’s examine heat treatment and Salmonella control in feed processing as a prime example.
Moisture is not merely a percentage point in the final product; it is a fundamental component of high-quality feed. While much has been written about its influence on pellet quality, energy efficiency, and starch gelatinization, its role extends much further. Moisture is one of the most critical parameters influencing the effectiveness of Salmonella control in feed manufacturing. Its impact is observed across multiple stages, including thermal treatment, chemical control using organic acids, and post-processing stability during storage and handling.
Thermal processing and microbial resistance
From a thermal processing perspective, moisture directly affects the heat resistance of Salmonella. In low-moisture environments, such as dry feed (10–11% moisture), Salmonella cells exhibit significantly increased thermal resistance. This is primarily because reduced moisture stabilizes cellular structures and limits heat-induced damage. As demonstrated by Gautam et al. (2020), decreasing moisture leads to increased survival of Salmonella during heat exposure. Consequently, higher temperatures or longer retention times are required to achieve equivalent microbial reduction in dry feed.
In contrast, the presence of moisture – especially in the form of steam during conditioning – enhances heat transfer and increases microbial susceptibility. Coe et al. (2022) showed that effective reductions (>6 log₁₀) of Salmonella in feed could be achieved under hydrothermal conditions, confirming that temperature, moisture, and time must be considered together. Moisture facilitates protein denaturation within bacterial cells and disrupts membrane integrity, significantly improving the lethality of heat treatment.
The role of organic acids
Moisture also plays a key role in the efficacy of organic acids used for Salmonella control. Organic acids act primarily through their undissociated form, which penetrates bacterial cell membranes. This mechanism is highly dependent on the presence of water. Liquid acids, already in an aqueous phase, are immediately active and capable of rapid antimicrobial action. Powder acids, on the other hand, require moisture for dissolution, diffusion, and activation. Under dry conditions, their antimicrobial effect is delayed or reduced; however, in conditioned feed, they can approach the efficacy of liquid acids.
When comparing powder versus liquid acids, it is important to distinguish between immediate efficacy in feed hygiene and biological efficacy in the bird. Liquid acids are typically more effective for rapid feed decontamination because they distribute more readily and do not require the same degree of moisture activation. Powder acids and salts may be less aggressive, easier to handle, and more stable during storage, providing a longer-lasting effect against recontamination. However, their performance depends heavily on feed moisture, conditioning, and release characteristics.
In the bird, protected or coated acids may outperform free liquid acids in later gut segments because they are designed to survive the upper digestive tract. Therefore, the definition of ‘better’ depends on the target: surface/feed kill, residual feed hygiene, or gut modulation. Direct comparative evidence remains limited, so this distinction should be viewed as a mechanistic interpretation rather than a universal ranking.
Balancing hygiene and nutritional quality
The interaction between heat treatment and organic acids also affects broiler performance. Research by Goodarzi Boroojeni et al. indicates that thermal processing severity changes nutrient digestibility. Their work shows that harsh conditioning can reduce ileal nutrient digestibility, while organic acid inclusion can improve early feed efficiency and help maintain performance. This is a vital practical point: the most aggressive hygienization strategy is not necessarily the best biological strategy. A feed mill can reduce microbial risk but may lose nutritional value if the thermal load is excessive.
Additionally, moisture improves the distribution and penetration of acids into microenvironments where bacteria may be protected, such as within dust particles or organic matrices. However, excessive moisture can dilute acids and reduce their local concentration. As in many aspects of processing, balance is the key.
Post-process hygiene and recontamination
Reviews of Salmonella in feed manufacturing emphasize that even heat-treated feed may become contaminated again via dust, coolers, conveyors, or storage. While moisture and heat determine the success of the initial ‘kill step,’ post-process hygiene determines whether those gains are maintained. This is why chemical control measures are usually discussed as complements to – not replacements for – hydrothermal processing and mill hygiene.
Practical conclusions
Moisture acts as both an enabler and a risk factor. It enhances heat and acid efficacy during processing but can increase microbial risk if not properly managed after production. Effective Salmonella control requires an integrated approach. The research supports three practical conclusions:
Moisture significantly enhances the effectiveness of heat treatment; dry feed protects Salmonella and increases its thermal resistance.
Moisture influences acid efficacy, with powder forms being more moisture-dependent than liquid forms for rapid action.
Organic acids can support animal performance, particularly body weight gain and feed efficiency.
With products like Surf-Ace, we can achieve increased pellet output, improved conditioning, enhanced durability of the pelleted feed, reduced fines formation, and improved overall quality of the final feed product. However, the best feed hygiene strategy is not to rely on one tool alone, but to also integrate controlled moisture, appropriate thermal treatment, organic acid application (such as Acidomix, whose strong antimicrobial effects help improve feed hygiene and help prevent / control salmonella), and strict post-pellet hygiene into a single cohesive system. We just need to select the right tools to achieve the results we want.
References
Abd El-Ghany, W. A. (2024). Applications of organic acids in poultry production: An updated and comprehensive review. Agriculture, 14(10), 1756. https://doi.org/10.3390/agriculture14101756
Coe, N., Wei, S., Little, C., & Shen, C. (2022). Thermal inactivation of Salmonella surrogate, Enterococcus faecium, in mash broiler feed pelleted in a university pilot feed mill. Poultry Science, 104(5), 104998. https://doi.org/10.1016/j.psj.2025.104998
Gautam, M., Lian, K., Jin, Y., Steinbrunner, P., & Tang, J. (2020). Water activity influence on the thermal resistance of Salmonella in soy protein powder at elevated temperatures. Food Control, 113, 107160. https://doi.org/10.1016/j.foodcont.2020.107160
Goodarzi Boroojeni, F., Mader, A., Knorr, F., Vahjen, W., & Zentek, J. (2014). The effect of different thermal processing methods and carbohydrate sources on performance, nutrient digestibility and the intestinal microbiota of broiler chickens. Poultry Science, 93(5), 1152–1162. https://doi.org/10.3382/ps.2013-03632
Polycarpo, G. V., Burbarelli, M. F., Carão, A. C., Merseguel, C. E., Dadalt, J. C., Magalhães, R., … & Albuquerque, R. (2017). Effects of organic acids, probiotics and antibiotics on performance, gastrointestinal pH, and intestinal morphology of broiler chickens. Poultry Science, 96(1), 127–134. https://doi.org/10.3382/ps/pew270
Tomičić, Z., Čabarkapa, I., Čolović, R., Đuragić, O., & Tomičić, R. (2019). Salmonella in the feed industry: Problems and potential solutions. Journal of Agronomy, Technology and Engineering Management, 2(1), 130–139.
Van Immerseel, F., Russell, J. B., Flythe, M. D., Gantois, I., Timbermont, L., Pasmans, F., … & Ducatelle, R. (2006). The use of organic acids to combat Salmonella in poultry: A mechanistic explanation of the efficacy. Avian Pathology, 35(3), 182–188. https://doi.org/10.1080/03079450600711045
The 7 pillars of poultry health: A holistic strategy for disease control
by Madalina Diaconu, Business Development Manager, EW Nutrition
Modern poultry production is currently battling a perfect storm of respiratory, enteric, and bacterial pressures. These overlapping challenges do more than just make birds sick; they actively erode performance, lead to higher condemnation rates at the plant, and squeeze already tight profit margins. To stay ahead, any practical health program must move beyond quick fixes and instead align interventions across everything from gut integrity and immunity to farm management and data collection.
Despite significant technological leaps in biosecurity and disease control, many “old” enemies remain stubbornly persistent:
Coccidiosis: This remains the single largest financial drain on the industry, costing an estimated EUR 10.4 billion globally due to losses in weight gain and increased mortality. (Blake et al., 2020)
Necrotic Enteritis (NE): Often triggered by coccidiosis, NE ranges from “silent” subclinical performance loss to sudden, fatal outbreaks. (Hargis, 2024; Skinner et al., 2010)
Histomoniasis: In turkeys, this disease (often called Blackhead) frequently results in 80-100% mortality, made worse by the fact that there are currently no approved treatments in major markets. (Beer et al., 2022; Merck, 2024)
APEC/Colibacillosis: This is a major driver of bird loss and processing plant condemnations, complicated by a high prevalence of multi-drug resistance. (Apostolakos et al., 2021; Joseph et al., 2023; Kazimierczak et al., 2025)
Salmonella: This pathogen persists at critical production nodes, with varying strains moving through the production pyramid from breeders to the final product. (Siceloff et al., 2022)
Why a pillar-based approach?
In the real world, economic impact rarely comes from just one source. It is usually a “multi-factorial” mess where pathogens, poor environment, and weak biosecurity interact to cause a crash. For example, respiratory and enteric issues often amplify one another, such as when Mycoplasma gallisepticum (MG) meets a viral infection and APEC. Because of this, “siloed” interventions – that only look at one problem in isolation usually underperform. Specialists are increasingly calling for integrated prevention. In what follows, we propose a strategy built on seven core pillars, which touches on all critical points of poultry production and ensures reduced use of antibiotics and chemicals through the consistent use of phytogenics.
Pillar 1: Pathogen pressure & epidemiology
Respiratory pathogens like IBV or NDV often show up as mixed infections, leading to high morbidity and more condemnations. MG and MS amplify these chronic issues. (Liu et al., 2025; El-Gazzar, 2025; CFSPH) Enteric pathogens like Eimeria (coccidiosis) create the groundwork for Clostridium perfringens (NE) to thrive. (Blake et al., 2020; Hargis, 2024; Skinner et al., 2010)
The Phytogenic Lever: Essential oils and plant polyphenols can disrupt the membranes of bacteria like Salmonella and E. coli, lowering the overall intestinal load and reducing environmental shedding. (Gentile et al., 2025; Wickramasuriya et al., 2022)
Pillar 2: Immunity & Vaccination
Successful vaccination isn’t just about the bottle; it requires precise strain selection, prime/boost design, and correct application. This is especially true for managing AIV (Avian Influenza) under global risk-based strategies. (FAO/WOAH, 2025)
The Phytogenic Lever: Certain plant-based additives act as immunomodulators, boosting macrophage activity and helping birds maintain resilience even when stressed by high stocking densities or heat. (Wickramasuriya et al., 2022)
Pillar 3: Microbiome & Gut Integrity
“Dysbacteriosis” is essentially a microbiome out of balance, which ruins nutrient absorption and weakens the gut barrier. (Aruwa et al., 2021; Aruwa & Sabiu, 2024) Protecting the gut is essential because clinical NE can kill birds quickly, while subclinical NE silently ruins efficiency. (Hargis, 2024; Skinner et al., 2010)
The Phytogenic Lever: These additives support “good” bacteria like Lactobacilli while suppressing opportunists and strengthening the “tight junctions” in the gut lining. (Wickramasuriya et al., 2022) Multiple trials show reduced NE pressure when phytogenics accompany coccidiosis programs. (Wickramasuriya et al., 2022)
Pillar 4: Environment & Management
The environment plays a massive role; for instance, recycling litter beyond six cycles significantly increases the risk of Salmonella detection. (Machado et al., 2020) Proper ventilation is also key to preventing thermal stress, which can trigger gut dysbiosis. (Liu et al., 2025; Aruwa et al., 2021)
The Phytogenic Lever: By stabilizing digestion and the microbiota, these additives can reduce wet litter and ammonia release, indirectly improving respiratory comfort. (Wickramasuriya et al., 2022; Aruwa et al., 2021)
Pillar 5: Biosecurity & Movement Control
Disease spreads through networks. Prioritizing biosecurity at “high-centrality” nodes – like hatcheries and common service routes – is more effective than a blanket approach. (Sequeira et al., 2025)
The Phytogenic Lever: Reducing the amount of pathogens a flock sheds helps support structural biosecurity barriers by lowering the overall transmission risk within houses. (Gentile et al., 2025; Wickramasuriya et al., 2022)
Pillar 6: Water, Feed & Processing Interface
Water hygiene is a vital tool for microbiome stability, especially during the vulnerable brooding phase. (Wickramasuriya et al., 2022) At the processing plant, PAA chillers remain the most effective chemical intervention to reduce contamination. (Thames et al., 2022)
The Phytogenic Lever: Using phytogenics in feed or water helps stabilize the upper-GI tract during feed transitions and can lower carcass pathogen loads. (Gentile et al., 2025; Wickramasuriya et al., 2022)
Pillar 7: Diagnostics, Genomics & Data Systems
Modern tools like Whole Genome Sequencing (WGS) and RT-PCR panels allow for much faster detection of APEC or respiratory viruses, enabling “precision” interventions. (Kazimierczak et al., 2025; El-Gazzar, 2025; Liu et al., 2025)
The Phytogenic Lever: When data shows rising pathogen pressure, phytogenics offer a flexible, rapid-response alternative that helps maintain antibiotic stewardship. (Kazimierczak et al., 2025; Gentile et al., 2025)
A 12-Month Roadmap for Implementation
Q1: Baseline & Risk Map: Map pathogen pressure using targeted PCR/WGS panels and review movement networks to prioritize high-centrality nodes. (Kazimierczak et al., 2025; El-Gazzar, 2025; Liu et al., 2025; Siceloff et al., 2022; Sequeira et al., 2025)
Q2: Program Design: Update vaccine strains and set up co-management plans for coccidiosis and NE, including microbiome supports with clear targets. (Liu et al., 2025; El-Gazzar, 2025; Blake et al., 2020; Hargis, 2024; Wickramasuriya et al., 2022)
Q3: Execution & Plant Linkage: Solidify water/feed hygiene SOPs and link farm Salmonella trends to plant PAA chiller performance. (Siceloff et al., 2022; Thames et al., 2022; Sequeira et al., 2025)
Q4: Review & Scale: Audit how well the team followed the diagnostic-driven actions and refine the playbooks for the next cycle. (Kazimierczak et al., 2025)
The Integrated View
Phytogenic feed additives aren’t “silver bullets,” but they contribute across all seven pillars. Their multi-target mode of action – acting as anti-inflammatories, antioxidants, and antimicrobials – complements traditional tools like vaccines and biosecurity to build a more resilient bird. (Wickramasuriya et al., 2022; Gentile et al., 2025; Aruwa et al., 2021)
Gentile N, et al. Emerging challenges in Salmonella control: innovative, sustainable disinfection strategies in poultry farming. Pathogens (2025). https://www.mdpi.com/2076-0817/14/9/912
Siceloff AT, Waltman D, Shariat NW. Regional Salmonella differences in U.S. broiler production (2016–2020). Applied and Environmental Microbiology (2022). https://journals.asm.org/doi/10.1128/aem.00204-22
Skinner JT, et al. An economic analysis of subclinical necrotic enteritis in broilers. Avian Diseases (2010). [suspicious link removed]
Wickramasuriya SS, et al. Role of physiology, immunity, microbiota and infectious diseases in poultry gut health. Vaccines (2022). https://www.mdpi.com/2076-393X/10/2/172
The lessons of 2025 for poultry and feed producers
by Ilinca Anghelescu, Global Director Marketing & Communications, EW Nutrition
2025 was a year defined by four converging forces for the global feed and animal production industry: an unprecedented HPAI crisis that cost American consumers alone $14.5 billion extra in egg expenditures; historic record corn production driving feed ingredient prices lower; a highly disruptive US tariff regime that reshuffled global trade flows for soybeans, corn, chicken, and pork; and accelerating regulatory pressure on antimicrobial use across Europe and globally.
The strategic imperatives from 2025 are clear: biosecurity investment is no longer optional, ingredient price volatility demands agile procurement strategies, trade compliance is a weekly operational concern, and antibiotic-free production transitions require credible, phased plans now.
KEY METRIC: Global chicken meat production reached approximately 105 million MT in 2025 (+2%), even as egg production suffered severely. The global feed market is valued at $542 billion in 2025, growing at 3.3% CAGR. Corn hit record production of 17 billion bushels in the US alone – the highest since 1936 in terms of harvested area.
The H5N1 clade 2.3.4.4b strain of Highly Pathogenic Avian Influenza (HPAI) continued to dominate animal health headlines in 2025. Since its reemergence in February 2022, the US outbreak alone has resulted in the confirmed loss of over 175 million birds across 1,700+ flocks – the costliest poultry disease event in recorded history.
Metric
Data Point
Source
Total US birds affected (2022–2025)
175+ million
USDA APHIS, May 2025
US flocks confirmed positive
1,704+
USDA APHIS, May 2025
Proportion of affected birds: layers
75%
USDA / Congressional Research Service
US egg layer flock deficit vs. 2022
–8% fewer birds
CoBank / USDA
Consumer egg overspend (May 2024–Apr 2025)
$14.5 billion extra
Innovate Animal Ag analysis
Peak US retail egg price
$6.23/dozen (March 2025)
BLS / USDA
HPAI-related US taxpayer response costs
$1.8 billion+
Innovate Animal Ag
Global HPAI mammal outbreaks (2024)
1,022 (vs. 459 in 2023)
WOAH 2025
Countries self-declaring HPAI freedom (May 2025)
25
WOAH
1.2 2025-Specific Developments
United States: Early-Year Severity, Policy Response
The first six weeks of 2025 saw 28 million layers depopulated – the worst start to any calendar year on record. Ohio, Indiana, and Missouri bore the brunt. The USDA launched a five-pronged approach in February 2025 including:
Gold-standard biosecurity assessments (948 completed Jan 20–June 26)
Indemnity increase from $7 to $17 per lost layer hen
Importation of 26+ million dozen shell eggs from Brazil, Honduras, Mexico, Turkey, and South Korea
Removal of select regulatory burdens to accelerate flock repopulation
$793 million in HPAI research proposals received in response to USDA Innovation Grand Challenge
⚠ Price Manipulation Investigation: In April 2025, the DOJ Antitrust Division launched an investigation into the largest US egg producer after it reported a 247% increase in quarterly net income. Egg producers and retailers face ongoing scrutiny over whether crisis pricing exceeded what supply constraints warranted.
Brazil: First Commercial HPAI Outbreak – May 2025
On May 15, 2025, Brazil – the world’s largest poultry exporter, responsible for nearly 30% of global exports – confirmed its first-ever commercial HPAI case at a breeder facility in Montenegro, Rio Grande do Sul (17,000 birds). This was a watershed event for global poultry trade.
Consequence
Detail
China (#1 buyer of Brazilian chicken) suspended imports
Trade suspended as of May 2025; Chinese delegation visited RS in Sept 2025 to assess resumption
Brazil’s monthly poultry exports declined
Exports fell 12.9% to $655 million; volume down 14.4% to 363,100 MT (May)
UAE replaced China as Brazil’s top buyer
First time China dropped from #1 buyer since 2019
WOAH new 10-year global HPAI strategy launched
Prevention and Control of HPAI (2024–2033), February 2025
Regionalized trade bans helped contain damage
Bans limited to affected regions, not all of Brazil
Europe: Persistent Pressure
HPAI continued to circulate widely in European poultry and wild bird populations. Key 2025 events include recurrence in Australia (February), ongoing outbreaks in Germany, Hungary, Netherlands, UK, and France, and the first confirmed domestic cat HPAI death in the Netherlands (H5N1, November 2025).
CRITICAL RISK: HPAI is now classified as enzootic (endemic) in wild birds across North America by the CDC. The virus circulates year-round in wildlife reservoirs, making seasonal recurrence in commercial flocks a structural, not episodic, risk. US egg producers are 8% below their 2022 flock baseline.
EU-wide – statistically significant increase trend 2020–2024 per EFSA/ECDC joint report, March 2025
AMR pressure in broilers and layers; genomic surveillance being mandated by EU
Newcastle Disease (NCD)
Brazil – outbreak July 2024, RS state
First commercial NCD in Brazil since 2006; adds biosecurity burden on top of HPAI protocols
H5N1 in Dairy Cattle (USA)
Ongoing – cross-species spread to 50+ US states
Cattle-to-poultry transmission confirmed; biosecurity interfaces between dairy and poultry operations must be reviewed
HPAI – Antarctica
First confirmed case March 2024 (South Polar Skua)
Indicates virus reached every continent; unprecedented in poultry disease history
CHAPTER 2: GLOBAL POULTRY PRODUCTION
2.1 Global Output – 2025 Performance
Despite HPAI disruptions, global chicken meat production grew approximately 2% in 2025 to around 105 million MT (ready-to-cook), driven by demand resilience and lower feed costs for broiler production. Total global poultry meat (including turkey, duck, and others) is forecast to exceed 152 million MT for 2025, per FAO Food Outlook June 2025.
Country / Region
2025 Production Forecast (MT)
Year-on-Year Change
Key Driver
USA – Broilers
21.7 million MT
+1.4% vs. 2024
Strong hatchery data; lower feed costs; HPAI minimal in broilers
Affordability vs. beef; consumer demand in developing markets
OECD-FAO 10-Year Outlook (2025–2034)
The OECD-FAO Agricultural Outlook 2025–2034, released in July 2025, projects global poultry meat production will grow by over 19% to 173.4 million MT by 2034 compared to the 2022–24 average. Poultry will account for the majority of additional meat consumption globally, driven by:
Affordability relative to beef and pork, especially in price-sensitive emerging markets
Population and income growth in Southeast Asia, South Asia, and Sub-Saharan Africa
Rapid urbanization and expansion of Quick Service Restaurant (QSR) chains
Superior feed conversion ratio (FCR) and lower greenhouse gas emissions per kg of protein
STRATEGIC NOTE: In high-income countries, per capita poultry consumption growth is flattening as consumers focus increasingly on welfare, environment, and health attributes. Growth opportunity is almost entirely in middle-income markets. Product premiumization (antibiotic-free, cage-free, organic) is the North American and European story.
2.2 Egg Production – Crisis Sector
Egg production was the sector hardest hit by HPAI globally. In the US, 75% of all HPAI-affected birds were table-egg layers, despite layers comprising less than 4% of the total poultry population. This structural vulnerability reflects longer flock lifespans and, increasingly, cage-free housing adoption.
Indicator
2025 Data
US retail egg price peak
$6.23/dozen (March 2025)
US retail egg price decline from peak
–27% by June 2025 (wholesale –64%)
US retail egg price (January 2025)
$4.95/dozen – 96% higher than January 2024
USDA full-year 2025 egg price forecast
+41.1% vs. 2024 average
% of US laying flock in cage-free systems
~40% (120+ million birds)
Global hen egg production (2023 baseline)
91 million tonnes (~1.7 trillion eggs)
Global egg trade volume (2024)
Nearly doubled from prior years
⚠ Cage-Free Transition & Disease Vulnerability: Some analysts link cage-free housing to higher HPAI susceptibility. Regardless of epidemiological debate, the US cage-free market is now structurally undersupplied relative to corporate commitments made in 2014–2017. Producers face a squeeze: comply with welfare commitments while managing disease risk.
CHAPTER 3: FEED INGREDIENT MARKETS
3.1 Grain & Oilseed Prices – 2025 Summary
From a feed cost perspective, 2025 was broadly favorable for livestock and poultry producers. Record US corn production and generally adequate global grain and oilseed supplies put downward pressure on the major feed commodities, offering partial relief from the margin pressure of recent years.
Commodity
2025 Price Direction
Key 2025 Data
Implication for Feed
Corn (US)
DOWN –3.9% (3rd consecutive annual decline)
Record US crop: 17.0 billion bu; yield 186.5 bu/acre – record; harvested area highest since 1936
Favorable for poultry/swine FCR cost; season avg ~$4.15/bu projected
Soybean Meal
DOWN –4.3% (3rd consecutive decline)
Prices at lowest since early 2016 at one point; large South American supply weighing on markets
Significant reduction in diet protein cost; amino acid supplementation cost-competitive
Soybeans
UP slightly +3.3%
After 22.9% collapse in 2024; still well below historical peaks; US acreage declining
Bean oil +20.8% (energy diet component); meal-to-bean ratio remains attractive for crushers
Wheat (Chicago)
DOWN –4.3% (4th consecutive year)
Abundant global supply; Russia/Argentina record crops; increased feed use
Wheat competing with corn in feed formulations globally – inclusion rising in EU/Asia diets
Soybean Oil
UP +20.8%
Driven by biofuel demand (US 45Z renewable fuel credits)
Energy ingredient cost pressure; may affect fat inclusion rates in formulations
PROCUREMENT SIGNAL: The US/China trade tensions created windows of soybean buying opportunity as prices swung on trade deal news. China agreed to purchase US soybeans in late 2025 as part of a limited trade deal, causing a price uptick. Procurement teams should monitor US-China negotiations as a lead indicator for soybean pricing in 2026.
3.2 Global Feed Market Overview
Metric
2025 Data
Global animal feed market value
$542.36 billion
CAGR (2026–2034)
3.3%
Largest feed segment by additive type
Amino acids (33.6% share)
Largest feed segment by species
Poultry (dominant share)
Asia Pacific regional status
Dominant region (largest market)
Top feed ingredient challenge
Fluctuating prices for corn, SBM – still key risk for margin management
3.3 Key Ingredient Trends to Watch
Fertilizer Cost Relief
Fertilizer prices have declined significantly from their 2022 peak. A basket of N, P, and K fertilizers averaged $437/tonne in May 2025, down from the $815/tonne peak in April 2022, per FAO Food Outlook. This benefits grain production economics and should support adequate grain supplies into 2026.
Soybean Oil Competition: Biodiesel vs. Feed
US soybean oil demand from renewable fuel programs (the 45Z credit) competed directly with feed-grade fat supplies, pushing soy oil prices up 20.8% in 2025. Feed mills formulating with added fats should evaluate alternative lipid sources. Poultry fat and palm olein remain cost-competitive in some markets.
Alternative Proteins: Insect Meal, DDGS, Algae
While adoption remains limited in volume, regulatory acceptance of insect meal in EU poultry diets continues to expand. Dried Distillers Grains with Solubles (DDGS) remain a strategically important co-product, particularly in the US and EU. Feed formulators should have up-to-date matrix values and be prepared to use them when corn prices favor inclusions.
⚠ Tariff Risk for Feed Inputs: US feed manufacturers faced effective tariff rates averaging 12%+ on key agricultural inputs from China and other countries in 2025, including herbicides, pesticides, and some micro-ingredient precursors. Amino acid supplies (predominantly Chinese-origin lysine, methionine, threonine) faced added cost and supply uncertainty.
CHAPTER 4: TRADE POLICY DISRUPTIONS
4.1 The 2025 US Tariff Regime – Agricultural Impact
The Trump administration’s tariff policies beginning January 20, 2025, represented the most significant disruption to global agricultural trade in decades. The three largest US agricultural export markets – Mexico ($30.3B in 2024), Canada ($28.3B), and China ($24.7B) – were all targeted, triggering retaliatory measures that hit feed, grain, poultry, and pork exports.
Country
US Tariff (2025)
Retaliation on US Agriculture
Key Products Impacted for Feed/Poultry Industry
China
Reached 145% (paused to 30% via May 2025 truce)
15% on chicken, corn, wheat; 10% on soybeans, sorghum, pork – applied from March 2025
Chinese poultry buyers shifted away from US; US corn/soy export disruption; amino acid supply chain uncertainty
Canada
25–35% (escalated to 35% in Aug)
25% on US dairy, poultry, meat products ($21B)
Canada imports ~45% of US poultry exports; feed grain flows affected
Mexico
25–30% (USMCA-compliant goods largely exempted)
Retaliatory tariffs threatened on agricultural goods
Mexico is #1 market for US turkey exports; ongoing uncertainty
EU
14% (paused under negotiations)
Planned retaliation announced April 2025
Potential impact on US soy meal exports; EU feed ingredient costs
CHINA TRADE DEAL (MAY 2025): A 90-day tariff truce agreed May 12, 2025 reduced US tariffs on Chinese goods from 145% to 30%, and China’s tariffs on US products from 125% to 10%. China agreed to purchase US soybeans. No permanent deal was signed. The limited agreement provided short-term stability but medium-term uncertainty remains.
4.2 Impact on US Agricultural Trade Flows
Product
Trade Flow Change (2025)
Implication
Corn exports
UP >20% YoY
Record US production driving export competitiveness despite tariff uncertainty
Soybean exports
DOWN – China shifted to South America
Brazil and Argentina taking larger share of Chinese soy imports
US chicken exports
Maintained overall (6.8B USD)
Despite China restrictions, other markets (Middle East, Mexico) absorbed volume
US turkey exports
At risk – 10% of production exported; Mexico = 65% of turkey exports
HPAI + AMPV supply squeeze threatened export volumes at peak holiday season
Brazil chicken exports
Down 12.9% month of May impact; year-end positive
HPAI disruption in May/June; recovery in H2 2025 after regionalization
US egg imports (temporary)
26M dozen shell eggs imported
Emergency imports from Brazil, Honduras, Turkey, South Korea, Mexico to fill supply gap
4.3 Strategic Trade Lessons
Supply chain diversification is no longer a luxury: concentration of US soy exports to China created a single-point-of-failure vulnerability that became fully exposed in 2025.
Regionalized disease zoning is a trade-preserving tool: Brazil’s rapid implementation of regionalized HPAI bans (rather than country-wide) preserved most of its export access; this is the model the industry should support with regulators globally.
USMCA dependency is real: 70% of US corn, 60% of soybeans, 45% of poultry exports go to Mexico, Canada, China – the same three countries targeted by 2025 tariffs.
US government announced $12B in emergency farm compensation in 2025, repeating the pattern from Trump’s first term – indicating persistent trade disruption risk.
In 2025, the European Commission proposed a package to streamline EU food and feed safety legislation while maintaining high health standards. The initiative, announced mid-2025, is intended to boost competitiveness of EU producers by reducing regulatory complexity – a direct response to competitive concerns vs. non-EU producers.
5.2 EFSA 2025 Guidance on Microorganisms
On September 24, 2025, EFSA’s Scientific Committee adopted new harmonized guidance on the characterization of microorganisms in the food chain. This is a landmark shift with major implications for feed additive manufacturers, probiotics suppliers, and novel food applicants.
Key Element
Operational Implication
Whole Genome Sequencing (WGS) now mandatory for strain-level ID of all bacteria, yeasts, fungi, viruses in applications
All existing microbial feed additive dossiers must be reviewed; WGS data cannot be more than 2 years old at time of submission
Genomics-first approach to AMR assessment
Any AMR gene hit in curated databases triggers mandatory case-by-case assessment; significantly raises the regulatory bar for probiotics and fermentation products
Replaces multiple previous guidance documents
Companies must align R&D, QC, and regulatory documentation to new unified standard immediately
GM microorganisms: clearer differentiation
Products ‘produced by GMO’ now distinguished from ‘GMO active agents’ – critical for enzyme and probiotic positioning
Non-compliance = application rejection risk
Early non-alignment causes ‘clock-stops’ or formal rejection at EFSA suitability check stage
AMR remains the defining long-term regulatory risk for the animal feed and production industry. Key 2025 actions:
EFSA/ECDC Joint Report (March 2025): Highlighted persistently high resistance to critical antimicrobials in poultry, especially Campylobacter and Salmonella, with ‘statistically significant increasing trend 2020–2024.’ This directly fuels EU legislative pressure.
EU Regulation 2019/6 (Veterinary Medicines) – Article 118: Banning import of animal products containing antimicrobials used for growth promotion. Application delayed to 2026, raising questions about enforcement timelines – and competitive fairness regarding imports from countries still allowing AGPs.
EU AMR Implementation Decision 2023: New harmonized monitoring requirements for AMR in zoonotic and indicator bacteria from food-producing animals – effective January 1, 2025. All EU Member States now required to collect and report standardized AMR surveillance data.
WOAH 10-Year HPAI Strategy (2024–2033): Promotes surveillance, vaccination programs, and timely reporting as cornerstones of international HPAI management.
BOTTOM LINE ON AMR: The regulatory trajectory is clear and irreversible – sub-therapeutic antibiotic use for growth promotion is being eliminated globally. The timeline varies by region (already banned in EU since 2006; US voluntary approach from 2017; global WHO action plan). Companies that have already invested in transition are ahead; those that have not face increasing compliance risk and market access restrictions.
$793M in proposals received (417 submissions); awards expected by fall 2025; covers prevention, vaccines, therapeutics
DOJ Antitrust Investigation – Egg Producers
Launched April 2025; examining price-fixing allegations amid 247% profit increase by largest producer
Meat & Poultry Special Investigator Act (S.1312)
Proposed creation of Office of Special Investigator for Competition Matters within USDA – pending
Food Security & Farm Protection Act (S.1326)
Would prohibit states from imposing certain standards on preharvest agricultural production sold in interstate commerce – relevant to cage-free mandates
CHAPTER 6: FEED ADDITIVE & NUTRITION STRATEGIES
PRECISION NUTRITION SIGNAL: The industry’s shift to reduced crude protein (CP) diets, precisely supplemented with industrial amino acids (L-Lys, DL-Met, L-Thr, L-Trp, L-Val) remained the dominant reformulation strategy in 2025. Lower CP diets reduce feed cost, lower N excretion (environmental benefit), and reduce substrate for pathogenic bacteria. With amino acid prices remaining favorable, there are few economic arguments for maintaining high CP diets.
6.1 The Post-AGP Transition: Where the Industry Stands
The antibiotic-free (ABF) production movement accelerated further in 2025. With the EU ban on AGPs in place since 2006 and the US moving toward voluntary phase-out, the entire industry is in active transition. The key challenge: AGP removal creates enteric health gaps that must be addressed with alternative tools. Without effective management, removal of AGPs leads to increased necrotic enteritis, Campylobacter colonization, and poorer FCR.
6.2 Heat Stress – A Growing Production Challenge
Climate-related heat stress was a highlighted research and production topic in 2025. Modern high-performance broiler genetics have been selectively bred for rapid growth under thermoneutral conditions. Heat stress impairs feed intake, FCR, immunity, meat quality, and reproduction. Management strategies:
Vitamin C and E supplementation at heat stress periods
Betaine inclusion as an osmolyte; reduces supplemental methionine requirement under heat stress
Feed schedule adjustment (limit feeding during hottest hours; early morning/evening feeding)
Housing design investment: tunnel ventilation, evaporative cooling, adequate air velocity
6.3 In Ovo Technology
In ovo vaccination and nutrition delivery continued to advance in 2025. Key developments include high-throughput systems (3,000 eggs/hour at 99% accuracy) for in ovo vaccination and nutritional interventions. Early-life gut programming through in ovo delivery of probiotics, nutrients, and vaccine antigens is becoming an increasingly important hatchery-level biosecurity and performance tool.
CHAPTER 7: MARKET TRENDS & CONSUMER SHIFTS
7.1 Poultry Gaining Share vs. Other Proteins
Elevated beef prices throughout 2025 – driven by tight US cattle supply (herd at decades-long lows) and high demand – continued to push consumers toward poultry as a cost-effective protein. This dynamic is a structural tailwind for the broiler industry globally.
Market Dynamic
Detail
US broiler net cash farm income 2025
+27% YoY – livestock sector outperforms crop side
Global poultry market value (2025)
$316.77 billion; projected $433.98B by 2034 (CAGR 3.56%)
Global poultry export growth 2025
+1.8% to 16.9 million MT
Supermarkets poultry market share
42.1% of poultry distribution (2024)
Online poultry retail growth rate
CAGR 11.4% (fastest growing channel)
Italy – poultry share of total meat consumed
>44% in 2025
FAO Meat Price Index – poultry
Decreased in 2025 from mid-2024 high (broiler ample supply)
7.2 Cage-Free & Animal Welfare Commitments
The cage-free transition is structurally undersupplied in the US. Corporate commitments made in 2014–2017 implied a need for 220 million cage-free layers by 2025–26. Current production is well below that target. This creates both a market opportunity (premium pricing) and a risk (HPAI vulnerability concerns in cage-free systems). Producers must balance welfare compliance with biosecurity protocols.
7.3 Antibiotic-Free, Organic, and Specialty Products
Consumer and corporate buyer demand for ABF, No Antibiotics Ever (NAE), organic, and pasture-raised products continued to grow in premium markets in 2025. The pasture-raised egg segment reported 30% annual growth rates despite high price points. For integrated producers, this requires dedicated production lines with separate management protocols, supply chain segregation, and robust documentation systems.
7.4 Sustainability Pressure
Feed manufacturers and integrators are under growing pressure from retail and foodservice customers, NGOs, and regulators to demonstrate reduced environmental footprint. Key metrics under scrutiny:
GHG emissions per kg of chicken meat produced (Scope 1, 2, and 3)
Deforestation-free supply chains for soy (EU Deforestation Regulation – EUDR)
Feed conversion ratio improvement as a sustainability lever
EUDR NOTE: The EU Deforestation Regulation requires companies to ensure that soy used in feed does not originate from recently deforested land. Implementation deadlines have been debated, but traceability requirements for soy origin – particularly from Brazil – are operationally significant for EU feed manufacturers and importers.
CHAPTER 8: STRATEGIC LESSONS & ACTION PRIORITIES
8.1 Summary: Top 10 Lessons of 2025
#
Lesson
Key Data Point
1
HPAI is now a permanent structural risk, not a cyclical one. Biosecurity investment must be treated as core capital expenditure.
CDC: H5N1 now enzootic in North American wild birds; US flock 8% below 2022 baseline
2
Egg production is structurally more vulnerable than broiler production – different biosecurity and business continuity protocols are required.
75% of HPAI losses = layers; broilers grew 1.4% in 2025
3
Vaccination for HPAI is the central unresolved debate of the decade – expect DIVA strategies to become standard within 3–5 years as industry and regulators align.
417 vaccine/research proposals submitted to USDA Grand Challenge
4
Trade concentration is a strategic vulnerability. Diversify export markets actively; do not allow 70%+ of any product to go to one trading bloc.
China + Mexico + Canada = 70% of US corn exports; 60% of soy; 45% of poultry
5
Grain prices are favorable NOW – lock in contracts and assess forward pricing opportunities while corn and SBM are at multi-year lows.
Corn -3.9% in 2025; SBM -4.3%; both 3rd consecutive annual decline
6
AMR regulations are accelerating everywhere. Transitioning to ABF production is no longer a ‘maybe’ but a ‘when’ – plan now.
EU: AMR in poultry ‘persistently high’ per EFSA/ECDC March 2025 report
7
EFSA’s 2025 WGS guidance fundamentally changes the cost and timeline of getting microbial feed additives authorized in the EU.
WGS now mandatory for all microbial characterizations; legacy dossiers need revision
8
Amino acids and precision nutrition remain the most cost-effective tool for diet optimization: lower CP, better FCR, lower N excretion, reduced enteric pathogen substrate.
Amino acids = 33.6% of global feed additive market by value
9
Brazil’s HPAI outbreak demonstrated both the vulnerability of global trade and the effectiveness of regionalized response protocols.
Brazil exports fell 12.9% in May but year-end positive; China temporarily banned; UAE stepped up
10
Climate/heat stress is an underappreciated production risk that compounds disease susceptibility and reduces performance in high-performing genetics.
IPCC: global surface temperature +0.9°C since mid-20th century; impacts on poultry FCR, immunity, mortality increasing
8.2 Action Priority Matrix for Management Teams
Priority Area
Immediate Actions (0–6 months)
Medium-Term (6–18 months)
HPAI Biosecurity
Complete USDA-style biosecurity assessments; audit wild bird access; upgrade water and air biosecurity; train all staff
Brazil HPAI market re-entry for China – recovery of the world’s #1 poultry export relationship
US corn/soy 2026 planting intentions (March) – USDA Prospective Plantings report is the key 2026 procurement signal
2025 demonstrated that the feed and animal production industry operates in an environment of simultaneous, compounding risks – biological, geopolitical, regulatory, and climatic. The companies that performed best were those with robust biosecurity infrastructure, agile procurement teams, clear AMR transition roadmaps, and diversified market exposure. There is no single silver bullet. Systematic risk management, not reactive crisis response, is the competitive differentiator going forward.
KEY SOURCES & REFERENCES
This article draws on data and analysis from the following sources:
Organization
Document / Resource Referenced
USDA APHIS / FAS
HPAI flocks data (2025); Livestock & Poultry World Markets (Dec 2025); WASDE reports; Five-Pronged HPAI Strategy
FAO
Food Outlook June 2025; OECD-FAO Agricultural Outlook 2025–2034; FAO Meat Price Index
OECD
OECD-FAO Agricultural Outlook 2025–2034 (July 2025)
WOAH
HPAI Report #68 (Feb 2025); State of World Animal Health 2025; HPAI 10-Year Strategy 2024–2033
EFSA / ECDC
Joint AMR Report (March 2025); 2025 QPS updated list; EFSA 2025 Guidance on Microorganisms (Nov 2025)
PAHO / WHO
Epidemiological Update H5N1 in the Americas (Jan 2025)
Corn & Other Feed Grains Outlook (2025–26 WASDE updates)
Frontiers in Veterinary Science
Phytogenic feed additives – gut health modulation (Aug 2025); Antibiotic alternatives – One Health (Jul 2025)
Intrinsically Heat-Stable Xylanase: A New Standard for Improving Performance under High-Temperature Pelleting
Author: Ajay Bhoyar, Senior Global Technical Manager, EW Nutrition
The global use of feed enzymes has become a central feature of efficient monogastric animal production systems. Rising feed ingredient costs, tighter margins, and increasing regulatory pressure to reduce environmental impact have all accelerated enzyme innovation. At the same time, feed mills have shifted toward higher conditioning temperatures and time in pursuit of improved pellet durability, pathogen control, and throughput. However, this creates a hostile environment for most exogenous feed enzymes, which can lose significant activity under the harsh conditions of feed processing.
Historically, enzyme manufacturers have attempted to overcome heat degradation of by coating, encapsulating, or post-pelleting liquid application (PPLA) of enzymes. While these approaches provide partial solutions, they can also have limitations, including delayed enzyme activity, uneven distribution, reduced mixing uniformity, and reliance on specialized liquid enzyme applicators.
These limitations prompted a novel direction: enzymes designed or selected to be intrinsically heat-stable, capable of surviving pelleting without protective matrices.
This article highlights recent advancements in intrinsically heat-stable xylanase technology, explains its advantages over coated and post-pelleting enzyme solutions, and outlines its practical benefits for feed manufacturers, integrators, and nutritionists operating under modern high-temperature feed pelleting conditions.
Intrinsically Thermostable Enzymes
An enzyme is considered intrinsically heat-stable when its native protein structure resists unfolding and retains catalytic activity under high temperatures associated with feed processing—typically 80–95°C for 30–90 seconds. Unlike coated enzymes that rely on external protection, intrinsically thermostable enzymes depend on their internal protein architecture for heat tolerance. Enzymes from organisms living in compost, thermal springs, and geothermal soils naturally withstand temperatures of 80–100 °C or higher. Intrinsically thermostable enzymes are often sourced from thermophiles (organisms living in hot springs and deep-sea vents) or engineered for stability. They resist denaturation (loss of shape and function) at high-temperature processing.
Fig.1: Key benefits of intrinsically thermostable enzymes
Limitations of Current Thermostability Solutions
Coating / Encapsulation
A method of protecting enzymes from heat is to encapsulate or coat them with a protective coating. An ideal enzyme coating for animal feed needs to:
1. Protect the enzyme through steam conditioning (typically 85–90°C or higher) and through subsequent pelleting.
2. Release the enzyme from the coating quickly in the gastrointestinal tract of the target animal, to ensure optimum efficacy. (Gilbert and Cooney, 2007)
There is some evidence, however, suggesting that the coating of enzymes may reduce the efficacy of the product, compared to an uncoated version of the same product (Kwakkel et al., 2000).
Post-Pelleting Liquid Application (PPLA)
Post-pelleting liquid enzyme application requires sophisticated applicators to minimize the risk of uneven spraying or calibration errors, which is often not feasible in small or mid-size mills. Accurate application of the liquid enzyme, as with some other critical liquid micro-ingredients, requires specialized spraying equipment and, even then, consistency of accurate enzyme application can be an issue (Bedford and Cowieson, 2009). Research has shown that as much as 30% of the enzyme activity can be found in the pellet fines, and therefore, adding the enzyme before screening would result in a lower than expected dosage in the final feed and wastage of the enzyme product (Engelen, 1998). In some cases, adjusting the pelleting machines to the output of the PPLA’s spray nozzles to ensure a homogenous and even application of the enzyme on the pellets may reduce the overall pellet production rate, especially in big feed mills with very high throughput.
These limitations of the coated or PPLA technologies strengthen the value proposition of intrinsically heat-stable enzymes.
Nutritional and Commercial Benefits of Intrinsically Heat-Stable Xylanase
The use of intrinsically heat-stable xylanase delivers consistent nutritional benefits in poultry and swine feeds, including predictable non-starch polysaccharide (NSP) degradation, a significant increase in the metabolizable energy (ME) value of the feed, and enhanced gut health resilience supporting reduced antibiotic use.
From a commercial and operational perspective, this technology simplifies enzyme application, improves mixing uniformity, reduces formulation risk, and lowers feed cost per unit of meat or egg produced.
In-Vitro Thermal Stability Profile of Axxess XY
Axxess XY is a novel, intrinsically thermostable GH10 xylanase originating from Thermotoga maritima, a hyperthermophilic bacterium found in hydrothermal vents near volcanic grounds, and commercially it is produced by proprietary strain of Bacillus subtilis.
The superior heat stability of Axxess XY has been proven under various commercial pelleting conditions across different geographies. Axxess XY showed excellent post-pelleting recovery under commercial feed-milling conditions across varying temperatures and conditioning times (Fig. 2).
In one study, in addition to excellent post-pelleting recovery, Axxess XY also demonstrated high xylanase stability in pelleted feed over a 2-month feed storage period at>40°C, with humidity around 65%.
Fig.2: Demonstrated Intrinsic Thermostability of Axxess XY Across Geographies
Conclusions
As feed mills continue to operate at higher conditioning temperatures and longer retention times, enzyme heat stability has become a critical success factor in modern feed production. Intrinsically heat-stable xylanase offers a practical and reliable solution to this challenge by maintaining enzyme activity through pelleting without the need for coatings or post-pelleting liquid application systems.
By relying on its native protein structure rather than external protection, intrinsically thermostable xylanase delivers consistent post-pelleting recovery, uniform distribution in feed, and predictable nutritional performance across different feed mills and processing conditions. This reliability translates into improved nutrient utilization, better gut health support, and reduced cost per kilogram of meat or eggs produced.
From an operational standpoint, intrinsically heat-stable xylanase simplifies enzyme application, reduces dependence on specialized equipment, and minimizes the need for over-formulation or safety margins. These advantages help feed manufacturers and integrators improve efficiency, lower risk, and achieve more consistent results, especially under demanding commercial pelleting conditions.
In summary, intrinsically heat-stable xylanase aligns well with the evolving needs of today’s feed industry, offering a robust, cost-effective, and future-ready enzyme solution for high-performance animal production systems.
References:
Bedford, M. R., and A. J. Cowieson. 2009. “Phytate and Phytase Interactions.” In Proceedings of the 17th European Symposium on Poultry Nutrition, 7–13. Edinburgh, UK.
Eeckhout, M., M. De Schrijver, and E. Vanderbeke. 1995. “The Influence of Process Parameters on the Stability of Feed Enzymes during Steam Pelleting.” In Proceedings of the 2nd European Symposium on Feed Enzymes, 163–169. Noordwijkerhout, The Netherlands.
Engelen, G. M. A. 1998. Technology of Liquid Additives in Post-Pelleting Applications. Wageningen, The Netherlands: Wageningen Institute of Animal Science.
Gilbert, T. C., and G. Cooney. 2011. “Thermostability of Feed Enzymes and Their Practical Application in the Feed Mill.” In Enzymes in Farm Animal Nutrition, 2nd ed., edited by M. R. Bedford and G. G. Partridge, 249–259. Wallingford, UK: CABI.
Kwakkel, R. P., P. L. van der Togt, and K. A. B. M. Holkenborg. 2000. “Bio-Efficacy of Two Phytase Formulations Supplemented to a Corn–Soybean Broiler Diet.” In Proceedings of the 3rd European Symposium on Feed Enzymes, 63–64. Noordwijkerhout, The Netherlands.
Learning from AGP mechanisms to advance poultry nutrition
By Ilinca Anghelescu, Dr. Andreas Michels, Predrag Persak
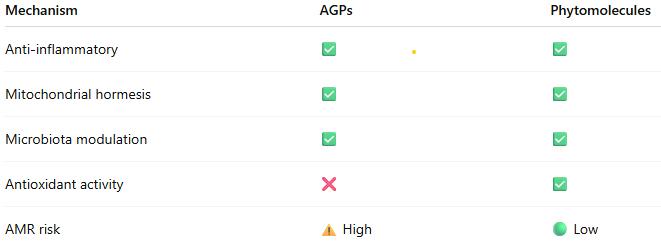
Our understanding of how nutrition influences growth and resilience in poultry has greatly expanded in recent years. It is now clear that animal performance stems to a large extent from a balance between metabolism, immune function, and the gut microbiome. These systems interact continuously, and even small nutritional or environmental changes can shift the animals’ physiological response. This growing knowledge has encouraged the development of nutritional strategies and feed components that work through adaptive, non-antibiotic mechanisms. One recent proposed explanation for these responses has rapidly gained ground: hormetic modeling.
Hormetic modeling describes how small or moderate doses of nutritional components can activate beneficial adaptive responses (improved resilience or metabolic efficiency), while excessive doses become harmful. This idea parallels, largely speaking, Paracelsus’s famous principle: “The dose makes the poison.” In poultry nutrition, such hormetic patterns are well recognized in nutrients like trace elements (selenium, zinc) and specific amino acids (for example, arginine). At optimal levels, these nutrients support antioxidant defense, growth, and immune balance, whereas excessive intake may cause oxidative or metabolic stress
This review examines the hormetic principle and its application to modern poultry/swine feeding concepts, exploring how balanced nutrient design and controlled inclusion of bioactive compounds can strengthen cellular adaptation, improve stress tolerance, and enhance production efficiency.
How do AGPs actually work?
Despite AGP’s widespread historical use, the precise mechanisms by which subtherapeutic doses of antibiotics enhance animal productivity remained poorly understood. Recent advances in systems biology and mitochondrial research propose new answers, much needed to develop future advanced nutritional systems.
The traditional explanations for AGP efficacy have focused primarily on antimicrobial effects:
reducing nutrient competition from microorganisms
decreasing harmful bacterial metabolites
improving gut wall morphology (thinner gut wall ➡ better nutrient absorption)
preventing subclinical infections
However, these mechanisms alone could not fully explain why different classes of antibiotics with diverse mechanisms of action produce similar growth-promoting effects (Gutierrez-Chavez et al., 2025).
Niewold (2007) hypothesized that the primary mechanism of AGPs is non-antibiotic anti-inflammatory activity, reducing the energetic costs of chronic low-grade inflammation. Inflammation diverts nutrients from growth toward immune responses, with cytokine production (particularly IL-1β, IL-6, and TNF-α) suppressing anabolic pathways (Kogut et al., 2018). AGPs appear to selectively inhibit pro-inflammatory cytokine production without completely suppressing immune function.
A paper published in 2024 by Fernandez Miyakawa et al. proposes that antibiotics at subtherapeutic levels act primarily through mitochondrial hormesis and adaptive stress responses, and not simply through antimicrobial activity. In this model, mitochondria act as bioenergetic hubs and signaling centers. Low-dose antibiotics trigger mild mitochondrial stress, which triggers the activation of adaptive protective pathways.This in turn induces mitokine release, leading to systemic adaptive responses improving growth, feed efficiency, and disease tolerance.
Mechanism of action in the hormetic model of AGP efficiency
Hormesis is a biphasic mechanism whereby high doses are toxic, but low doses stimulate adaptive responses and are beneficial. In the case of AGPs, Fernandez Miyakawa et al. propose that low doses stimulate growth, stress resistance, and cellular repair.
Key signaling pathways
As Bottje et al. (2006, 2009) shows, efficient animals often have mitochondrial inner membranes that are less permeable to protons and other ions, allowing for more effective coupling between electron transport and ATP synthesis, which reduces energy loss through proton leak and maximizes the production of ATP per oxygen molecule consumed. Lower membrane permeability is influenced by factors like decreased membrane surface area per protein mass, specific membrane protein content (such as adenine nucleotide translocase), and fatty acid composition in the membrane phospholipids, all contributing to a tighter barrier that prevents unregulated electron or proton flow and supports higher energetic efficiency. Such features make mitochondria in efficient species more capable of maintaining membrane integrity and ATP generation, especially when facing environmental stress, as seen in freeze-tolerant animals whose mitochondria do not undergo damaging permeability transitions under extreme conditions.
Nrf2
Many AGPs interfere with mitochondrial protein synthesis and electron transport chain. At subtherapeutic levels, they cause a mild ROS increase, which triggers the activation of redox-sensitive transcription factor Nrf2. Since Nrf2 regulates over 250 antioxidant, detoxification, and anti-inflammatory genes, the result is improved cell survival, redox balance, and tolerance to stress.
Figure 1 From Zhang et al., 2024
Mitokine production
Mitokines are “signaling molecules that enable communication of local mitochondrial stress to other mitochondria in distant cells and tissues” (Burtscher 2023). Through fibroblast growth factor 21 (FGF21), growth differentiation factor 15 (GDF15), adrenomedullin2 (ADM2) etc, these stress signals are released systemically and coordinate tissue-wide responses, leading to improved growth and resilience.
Inflammation and disease defense
While the negative side of antibiotic growth promoters is well researched and understood (Rahman et al., 2022), science can advance by isolating the positive effects and attempting to offer different pathways to the same benefits. One such lesson can be derived from understanding inflammation pathways and responses.
Chronic low-grade intestinal inflammation is common in modern poultry production, due to diet, microbiota shifts, high metabolic demands etc. This inflammation diverts energy from growth to immune responses.
AGPs reduce the energy costs of this inflammation in three main ways:
Reduces inflammation through adaptive stress response
Raising the threshold to trigger inflammation
Promoting overall resilience, rather than simply killing pathogens
Fernandez Miyakawa et al. suggest, in this emerging model, that disease defense can operate two different actions: resistance to health challenges through reduction of the pathogen load (which is driven by the immune system and is energy costly); and overall resilience by reducing host damage without reducing the pathogen load. AGPs, the authors claim, mainly promote resilience by enhancing mitochondrial stress responses and tissue repair, i.e. more precisely:
Direct mitochondrial stimulation in intestinal epithelial cells
Metabolic optimization supporting growth and feed efficiency
Figure 2 From Fernandez Miyakawa et al., 2024.
In this context, “metabolic optimization” refers to the enhancement of metabolic processes within livestock or poultry to support efficient growth, feed conversion, and physiological resilience, without relying on immune-mediated pathways that are energetically costly. Scientific evidence shows that metabolic optimization involves improving nutrient assimilation, promoting more efficient energy production in tissues (such as mitochondrial ATP synthesis), and minimizing wasteful metabolic byproducts, resulting in reduced feed intake per unit of growth and better utilization of dietary nutrients (Rauw 2025, El-Hack 2025).
Function of feed additives and feed components
Feed additives and feed components in many ways represent the complete other side of the spectrum from antibiotics, but are there some features where antibiotics and feed additives come close in their functions? There is a good case to be made for certain feed additives ultimately working in the animal to achieve similar benefits to the desirable, non-medicinal usage of AGP´s. Especially with the emergent model of AGP mechanism described above, it is worth discussing how certain feed additives can support the same end goal: promoting animal resilience.
Lillejhoj et al (2018), Gutierrez-Chavez et al. (2025) and others outline the end-results such products must achieve:
Growth performance & feed conversion efficiency
Promotion of animal productivity under real-world conditions
Support gut homeostasis
Non-adverse effect on the immune system
Reduction of oxidative stress
Support organism in mitigation of enteric inflammatory consequences
Within the hormetic model, possibly the most important systemic benefit is, in one phrase, promoting resilience. Phytomolecules have long been used, in human and animal medicine, for the same end goal. The mechanisms described below should naturally be seen with caution, as phytomolecule microbiome effects can be subtler and context-dependent. However, the substantiating literature has been increasingly accumulating on these specific topics.
1. Immunometabolic regulation
Phytomolecules demonstrate remarkably similar anti-inflammatory effects to what Niewold (2007) suggested was a primary mechanism of AGPs: non-antibiotic anti-inflammatory activity, reducing the energetic costs of chronic low-grade inflammation. Inflammation diverts nutrients from growth toward immune responses, with cytokine production (particularly IL-1β, IL-6, and TNF-α) suppressing anabolic pathways (Kogut et al., 2018). AGPs appear to selectively inhibit pro-inflammatory cytokine production without completely suppressing immune function. A similar effect can be observed with various types of phytomolecules, which significantly reduced pro-inflammatory and/or increased anti-inflammatory cytokine expression in animals challenged with several pathogens. The anti-inflammatory mechanism appears to involve inhibition of NF-κB activation and modulation of MAPK signaling pathways (Kim et al., 2010; Long et al., 2021).
2. Mitochondrial hormesis and energy metabolism
Fernández Miyakawa et al. (2024, see above) proposed that AGPs exert growth-promoting effects through mitochondrial hormesis – subtherapeutic antibiotic doses induce mild mitochondrial stress, triggering adaptive responses that enhance mitochondrial function, energy metabolism, and cellular resilience. This mechanism, while requiring further validation, explains why different antibiotics with diverse targets produce similar growth outcomes.
The mitochondrial stress response involves activation of the IL-6 receptor family signaling cascade, which regulates metabolism, growth, regeneration, and homeostasis in liver and other tissues (Perry et al., 2024). Subtherapeutic antibiotic exposure activates proteins involved in growth and proliferation through IL-6R gp130 subunit signaling, including JAK, STAT, mTOR, and MAPK pathways.
Phytomolecules demonstrate similar mitochondrial effects. Perry et al. (2024) showed that increased activity of AMPK, mTOR, PGC-1α, PTEN, HIF, and S6K can also be available via phytomolecule activity, suggesting enhanced anabolic metabolism.
Capsicum oleoresin supplementation in broilers increased jejunal lipase and trypsin activity, enhanced ileal amylase activity, improved jejunal morphology, and modulated immune organ development, indicating enhanced digestive efficiency and nutrient utilization (Li et al., 2022).
Compounds such as vanillin, thymol, eugenol have been shown to improve glucose and lipid metabolism through TRPV1 activation and mitochondrial function enhancement (Gupta et al., 2022; Zhang et al., 2017).
3. Gut microbiota modulation
AGPs selectively reduce specific microbial populations, particularly Lactobacillus species that produce bile salt hydrolase (BSH). Since BSH reduces fat digestibility and thus weight gain, AGP-mediated reduction of BSH-producing bacteria enhances energy extraction and growth (Lin, 2014; Bourgin et al., 2021).
Recent research by Zhan et al. (2025) using single-molecule real-time 16S rRNA sequencing demonstrated that therapeutic antibiotic doses (lincomycin, gentamicin, florfenicol, benzylpenicillin, ceftiofur, enrofloxacin) significantly altered chicken gut microbiota composition, with Pseudomonadota and Bacillota becoming dominant phyla after exposure. Different antibiotics produced distinct temporal effects on microbial diversity and community structure.
Phytomolecules exert targeted antimicrobial effects while promoting beneficial bacteria. Dietary supplementation with 800 mg/kg Capsicum extract in Japanese quails reduced cecal counts of pathogenic bacteria (Salmonella spp., E. coli, coliforms) while modulating Lactobacilli populations (Reda et al., 2020).
In pigs, 80 mg/kg natural capsicum extract increased cecal propionic acid and total volatile fatty acid concentrations, with increased butyric acid in the colon – indicating enhanced fermentation by beneficial bacteria (Long et al., 2021).
Capsicum and Curcuma oleoresins altered intestinal microbiota composition in commercial broilers challenged with necrotic enteritis, reducing disease severity through microbiome modulation (Kim et al., 2015).
Capsaicin demonstrates selective antimicrobial activity, inhibiting pathogenic Gram-negative bacteria while favoring development of certain Gram-positive bacteria. The antibacterial mechanism involves induction of osmotic stress and membrane structure damage (Adaszek et al., 2019; Rosca et al., 2020).
4. Intestinal barrier function and gut health
AGPs have been associated with improved intestinal morphology, including increased villus height and reduced crypt depth, which enhance absorptive capacity (Gaskins et al., 2002).
Phytomolecules produce similar or superior effects. Capsicum extract (80 mg/kg) in pigs increased ileal villus height and upregulated MUC-2 gene expression, indicating enhanced gut barrier function and integrity. The improved barrier function correlated with reduced diarrhea incidence (Liu et al., 2013; Long et al., 2021).
Allium hookeri extract increased expression of tight junction proteins (claudins, occludins, ZO-1) in LPS-challenged broiler chickens, demonstrating direct enhancement of barrier integrity (Lee et al., 2017).
5. Oxidative stress mitigation
Oxidative stress impairs growth by damaging cellular components and triggering inflammatory responses. AGPs reduce oxidative stress indirectly through anti-inflammatory effects and microbiota modulation (Bortoluzzi et al., 2021).
Phytomolecules possess direct antioxidant properties. Capsicum extract (50 mg/kg) in heat-stressed quails reduced serum and ovarian malondialdehyde (MDA) while increasing superoxide dismutase (SOD) and catalase (CAT) activities. Ovarian transcription factors showed decreased NF-κB and increased Nrf2 and HO-1 expression (Sahin et al., 2016).
A mixture of herbal extracts including pepper reduced thiobarbituric acid reactive substances and MDA in broiler liver and muscle, while increasing glutathione peroxidase (GSH-Px) activity and improving antioxidant enzyme expression (Saleh et al., 2018).
Capsicum extract (80 mg/kg) in pigs increased total antioxidant capacity, SOD, and CAT while reducing MDA levels, demonstrating robust antioxidant effects (Long et al., 2021).
Standardization and controlled release: Critical success factors
A major criticism of phytomolecules has been inconsistent efficacy across studies. However, this variability largely reflects differences in:
Active compound concentrations
Bioavailability and stability
Dosing precision
Product quality and standardization
Microencapsulation is one of the technologies that address the standardization and bioavailability challenges. It protects volatile compounds from degradation during feed processing and storage, with encapsulated essential oils showing significantly higher retention compared to unprotected forms (Stevanović et al., 2018). By creating a protective barrier around active ingredients, microencapsulation enables controlled release in specific regions of the gastrointestinal tract, improving absorption efficiency and reducing dose variability (Bringas-Lantigua et al., 2011). The technology also masks unpalatable flavors that can reduce feed intake while standardizing active ingredient concentrations through precise manufacturing processes (Gharsallaoui et al., 2007). Studies demonstrate that spray-dried microencapsulated essential oils achieve encapsulation efficiencies exceeding 93% with minimal loss during storage (Hu et al., 2020), and can be engineered for enzyme-mediated release to ensure bioactive delivery at optimal intestinal sites (Elolimy et al., 2025).
Mechanistic synthesis: An integrated model
The evidence indicates that both AGPs and phytomolecules operate through an integrated network of effects:
Primary Level: Selective antimicrobial effects modify gut microbiota composition
This integrative model explains why multiple antibiotics with different mechanisms produce similar growth outcomes: they converge on common pathways regulating immunometabolism and mitochondrial function (Fernández Miyakawa et al., 2024).
Phytomolecules operate through the same mechanistic framework but with potential advantages:
Safety and antimicrobial resistance considerations
Antibiotic exposure significantly disrupts gut microbiota diversity and stability, with effects persisting beyond withdrawal periods. The study by Zhan et al. (2025) demonstrated that different antibiotics produce varying degrees of microbiota disruption, with florfenicol and gentamicin showing the strongest and most persistent effects.
In contrast, phytomolecules generally do not generate resistance through the same mechanisms as antibiotics. Some phytochemicals may actually enhance antibiotic efficacy and resensitize resistant bacteria through structural modifications of bacterial membranes (Khameneh et al., 2021; Suganya et al., 2022).
However, one study reported increased correlation between antibiotic resistance genes (ARGs) and mobile genetic elements in pig feces after mushroom powder supplementation, suggesting that certain phytogenic compounds may increase ARG mobility (Muurinen et al., 2021). This emphasizes the need for continued surveillance of phytomolecule effects on resistance gene dynamics.
Capsaicinoids and capsinoids have well-established safety profiles. Capsiate, a non-pungent analogue of capsaicin, exhibits substantially lower toxicity while maintaining similar metabolic and growth-promoting effects (Gupta et al., 2022). No adverse effects on animal health or product quality have been reported at recommended dosages in reviewed studies.
Future directions and research needs
Despite substantial progress, several areas require further investigation:
Mechanistic refinement: Detailed characterization of signaling pathways, particularly the IL-6R/gp130 cascade and mitochondrial stress responses
Precision formulation: Development of combinations optimized for specific production stages, environmental conditions, and disease pressures
Bioavailability optimization: Advanced delivery systems ensuring consistent active compound release and absorption
Microbiome-host interaction mapping: High-resolution characterization of microbial community shifts and their functional consequences
Economic validation: Large-scale production trials assessing cost-effectiveness compared to AGPs and disease management costs
Conclusions
The scientific evidence demonstrates that standardized phytomolecules operate through well-characterized biological mechanisms that substantially replicate those of AGPs:
Anti-inflammatory effects reducing energetic costs of immune activation
Mitochondrial hormesis enhancing energy metabolism and cellular resilience
Selective microbiota modulation supporting beneficial bacteria while controlling pathogens
Intestinal barrier enhancement improving nutrient absorption and reducing translocation
Antioxidant activity mitigating oxidative stress and supporting immune function
When properly standardized and formulated for controlled release, phytomolecules deliver growth promotion, feed efficiency improvements, and disease resistance comparable to AGPs, while potentially offering advantages in AMR risk profile, stress resilience, and consumer acceptance.
The mechanistic convergence between AGPs and phytomolecules, coupled with demonstrated efficacy in controlled trials, provides producers with confidence that science-based phytomolecular interventions represent legitimate alternatives to AGPs. Success depends on product standardization, appropriate dosing, and understanding that phytomolecules work through fundamental biological pathways rather than undefined or mystical mechanisms.
As the livestock industry continues to navigate the post-AGP era, standardized phytomolecules offer a scientifically sound, mechanistically validated approach to maintaining animal performance, health, and welfare while addressing antimicrobial resistance concerns.
References
Adaszek, Ł., et al. “Properties of Capsaicin and Its Utility in Veterinary and Human Medicine.” Research in Veterinary Science, vol. 123, 2019, pp. 14 – 19.
Bottje, W., et al. “Mitochondrial proton leak kinetics and relationship with feed efficiency within a single genetic line of male broilers”. Poultry Science, Volume 88, Issue 8, 1 August 2009, p. 1683-1693.
Bortoluzzi, C., et al. “A Protected Complex of Biofactors and Antioxidants Improved Growth Performance and Modulated the Immunometabolic Phenotype of Broiler Chickens Undergoing Early Life Stress.” Poultry Science, vol. 100, 2021, p. 101176.
Bourgin, M., et al. “Bile Salt Hydrolases: At the Crossroads of Microbiota and Human Health.” Microorganisms, vol. 9, no. 1122, 2021.
Bravo, D., et al. “A Mixture of Carvacrol, Cinnamaldehyde, and Capsicum Oleoresin Improves Energy Utilization and Growth Performance of Broiler Chickens Fed Maize-Based Diet.” Journal of Animal Science, vol. 92, 2014, pp. 1531 – 1536.
Bringas-Lantigua, M., et al. “Influence of Spray-Dryer Air Temperatures on Encapsulated Mandarin Oil.” Drying Technology, vol. 29, 2011, pp. 520–526.
Burtscher, J., et al. “Mitochondrial Stress and Mitokines in Aging.” Aging Cell, vol. 22, no. 2, 2023, e13770.
El-Hack, M. et al. “Integrating metabolomics for precision nutrition in poultry: optimizing growth, feed efficiency, and health”. Frontiers in Veterinary Science, Sec. Animal Nutrition and Metabolism, Volume 12 – 2025. https://doi.org/10.3389/fvets.2025.1594749
Elolimy, Ahmed A., et al. “Effects of Microencapsulated Essential Oils and Seaweed Meal on Growth Performance, Digestive Enzymes, Intestinal Morphology, Liver Functions, and Plasma Biomarkers in Broiler Chickens.” Journal of Animal Science, vol. 103, 2025, p. skaf092, https://doi.org/10.1093/jas/skaf092.
Fernández Miyakawa, Mariano E., et al. “How Did Antibiotic Growth Promoters Increase Growth and Feed Efficiency in Poultry?” Poultry Science, vol. 103, no. 2, 2024, article 103136. https://doi.org/10.1016/j.psj.2023.103136
Gaskins, H. Rex, C. T. Collier, and D. B. Anderson. “Antibiotics as Growth Promotants: Mode of Action.” Animal Biotechnology, vol. 13, no. 1, 2002, pp. 29 – 42.
Gharsallaoui, A., et al. “Applications of Spray-Drying in Microencapsulation of Food Ingredients: An Overview.” Food Research International, vol. 40, no. 9, 2007, pp. 1107-21.
Gutiérrez-Chávez, Vanesa, et al. “Capsaicinoids and Capsinoids of Chilli Pepper as Feed Additives in Livestock Production: Current and Future Trends.” Animal Nutrition, vol. 22, 2025, pp. 483 – 501. https://doi.org/10.1016/j.aninu.2025.03.014.
Gupta, A., et al. “Capsaicin and Capsinoids: Recent Updates on Their Health Benefits and Mechanisms of Action.” Phytotherapy Research, vol. 36, no. 5, 2022, pp. 1898 – 1912.
Hu, Q., Li, X., Chen, F., Wan, R., Yu, C.-W., Li, J., McClements, D. J., & Deng, Z. (2020). “Microencapsulation of an essential oil (cinnamon oil) by spray drying: Effects of wall materials and storage conditions on microcapsule properties“. Journal of Food Processing and Preservation, 44(11). https://doi.org/10.1111/jfpp.14805
Khameneh, B., et al. “Mechanisms of Antibiotic Resistance Resensitization by Phytochemicals: Review.” Phytomedicine, vol. 85, 2021, p. 153529.
Kim, D. K., et al. “Effects of Capsicum and Curcuma on Necrotic Enteritis in Broilers.” Poultry Science, vol. 94, 2015, pp. 2314 – 2321.
Kim, J. S., et al. “Anti-inflammatory Effects of Plant-Derived Molecules via NF-κB and MAPK Pathways.” International Immunopharmacology, vol. 10, no. 3, 2010, pp. 306 – 314.
Lee, S. H., et al. “Allium Hookeri Extract Enhances Tight Junction Proteins in Broilers.” Journal of Animal Physiology and Animal Nutrition, vol. 101, no. 1, 2017, pp. e48 – e56.
Li, X., et al. “Capsicum Oleoresin Supplementation Improves Digestive Enzyme Activity and Gut Morphology in Broilers.” Poultry Science, vol. 101, no. 7, 2022, p. 101844.
Lin, J. “Effect of Antibiotics on the Intestinal Microbiota and Their Role in Animal Growth.” Animal Biotechnology, vol. 25, no. 3, 2014, pp. 149 – 157.
Lillehoj, H., et al. “Phytochemicals as Antibiotic Alternatives to Promote Growth and Enhance Host Health.” Veterinary Research, vol. 49, no. 76, 2018.
Liu, Y., et al. “Dietary Capsicum Extract Enhances Intestinal Barrier Function and Growth in Pigs.” Journal of Animal Science, vol. 91, 2013, pp. 518 – 525.
Long, L., et al. “Phytogenic Feed Additives Modulate Intestinal Immunity and Antioxidant Status in Pigs and Poultry.” Frontiers in Veterinary Science, vol. 8, 2021, p. 620998.
Muurinen, J., et al. “Mushroom Powder Supplementation Increases Antibiotic Resistance Gene Mobility in Pig Feces.” Frontiers in Microbiology, vol. 12, 2021, p. 676678.
Niewold, T. A. “The Non-antibiotic Anti-inflammatory Effect of Antimicrobial Growth Promoters, the Real Mode of Action? A Hypothesis.” Poultry Science, vol. 86, 2007, pp. 605 – 609.
Perry, F., C. N. Johnson, L. Lahaye, E. Santin, D. R. Korver, M. H. Kogut, and R. J. Arsenault. “Protected Biofactors and Antioxidants Reduce the Negative Consequences of Virus and Cold Challenge by Modulating Immunometabolism via Changes in the Interleukin-6 Receptor Signaling Cascade in the Liver.” Poultry Science, vol. 103, no. 9, 2024, article 104044. https://doi.org/10.1016/j.psj.2024.104044
Rahman, Md, et al. “Insights in the Development and Uses of Alternatives to Antibiotic Growth Promoters in Poultry and Swine Production.” Antibiotics, vol. 11, no. 6, 2022, p. 766, https://doi.org/10.3390/antibiotics11060766.
Reda, F. M., et al. “Capsicum Extract Supplementation Modulates Gut Microbiota and Performance in Japanese Quails.” Animal Feed Science and Technology, vol. 265, 2020, p. 114507.
Rosca, I., et al. “Capsaicin Induces Osmotic Stress in Gram-negative Pathogens.” Veterinary Sciences, vol. 7, no. 4, 2020, p. 172.
Sahin, K., et al. “Dietary Capsicum Extract Reduces Oxidative Stress in Heat-stressed Japanese Quails.” Poultry Science, vol. 95, no. 2, 2016, pp. 231 – 240.
Saleh, A. A., et al. “Herbal Extract Mixtures Improve Antioxidant Status and Performance in Broilers.” Poultry Science, vol. 97, no. 11, 2018, pp. 3927 – 3936.
Stevanović, Z. D., et al. „Essential oils as feed additives—Future perspectives”. Molecules, 23(7), 2018, pp1717.
Suganya, R., et al. “Phytochemicals in Combination with Antibiotics: Antimicrobial Resistance Breakers.” Antibiotics, vol. 11, 2022, p. 123.
Zhang, Benyuan et al. “Mitochondrial Stress and Mitokines: Therapeutic Perspectives for the Treatment of Metabolic Diseases.” Diabetes & Metabolism Journal vol. 48,1, 2024, pp. 1-18.
Zhan, Ru, et al. “Effects of Antibiotics on Chicken Gut Microbiota: Community Alterations and Pathogen Identification.” Frontiers in Microbiology, vol. 16, 2025, article 1562510. https://doi.org/10.3389/fmicb.2025.1562510
Zhang, Y., et al. “Effects of Vanillin, Thymol, and Eugenol on Glucose and Lipid Metabolism via TRPV1 Activation.” Journal of Agricultural and Food Chemistry, vol. 65, no. 13, 2017, pp. 2719 – 2727.
Energy Metabolism in Pigs: Disease and stress impact efficiency
By Dr. Inge Heinzl, Editor, and Predrag Persak, Regional Technical Manager North Europe
For profitable pig production, efficient energy metabolism is essential. Every kilojoule consumed must be wisely spent – on maintenance, growth, reproduction, or defense. An impacted energy metabolism due to disease or stress impacts animal performance and farm profitability.
Different faces of energy
Energy metabolism determines how efficiently pigs convert feed into body mass. The Gross energy (GE) of the diet, which the use of a calorimeter can determine, is progressively reduced by losses in feces (→digestible energy – DE), urine, gases (→metabolizable energy – ME), and heat, resulting in the →net energy (NE), which is then available for maintenance and performance (growth, milk…).
The requirements for maintenance include the minimum energy that an organism needs to maintain essential functions under standardized conditions and at complete rest. This includes respiration, thermoregulation, tissue turnover, and immune system activity. Only energy in excess of these needs is available for performance. The ratio between additional retained energy and additional energy intake defines the incremental efficiency of nutrient utilization. Under normal conditions, healthy, fast-growing pigs display high incremental efficiencies for both protein and energy deposition by channeling energy efficiently into lean tissue and approximately 25-30% of the metabolizable energy from the feed is used for maintenance, 20-25% for lean gain, and the rest for fat deposition, driving daily gain and carcass quality (Patience, 2019).
However, disease, immune stress, and suboptimal environmental conditions can disrupt this delicate balance, diverting nutrients from growth to survival processes (Obled, 2003). The activation of the immune system leads to reduced feed efficiency, slower growth, and inferior meat quality.
Disease generates costs
The health challenge of disease causes energy loss through several key mechanisms (Patience, 2019).
The activation of the immune system becomes an energetic priority. It consumes significant amounts of energy and nutrients, such as glucose and specific amino acids, to produce immune cells and acute-phase proteins, such as haptoglobin and CRP, and to combat pathogens. The nutrients are redirected away from performance toward immune defense, i.e., less energy available for growth performance or even a mobilization of body reserves (fat deposits). A study conducted by Huntley et al. (2017) showed a 23.6% higher requirement for metabolizable energy to activate and maintain the immune system, resulting in a 26% lower ADG.
Physiological responses to disease, such as fever (heat production), shivering, or increased physical activity due to discomfort or listlessness, require energy.
Additional lower feed intake due to reduced appetite, leading to less energy consumption and intensifying the problem of energy repartitioning.
Environmental challenges are energy-consuming
Besides environmental conditions that cause disease due to high pathogenic pressure, environmental challenges are often related to thermoregulation.
1. Cold stress
In the case of cold stress, the ambient temperature falls below the pig’s lower critical temperature. The animal must spend extra energy to produce heat and maintain a constant body temperature. Alternatively, it can achieve this through shivering (muscle friction generates heat) and the release of thyroid hormones, which increase the metabolic rate and boost body temperature. Another possibility is huddling with other pigs. If the pigs eat more to gain extra energy for warmth, they increase production costs.
2. Heat stress
Excessive temperature leads to heat stress, and the animals attempt to cope through several mechanisms. Increased respiratory evaporation by panting is energy-intensive. Other possibilities are lying spread out on cool surfaces (conduction), seeking shade, and reducing physical activity to minimize heat production. To reduce metabolic heat production, pigs decrease their feed intake; however, this results in an energy deficit and likely mobilizes body reserves, especially in lactating sows.
3. Poor housing and management
High ventilation rates, draughts, wet floors, high stocking densities, and, too often, mixing of pigs are other stressors that require adequate energy-consuming responses. Also, an environment that facilitates excessive heat loss, e.g., through cold concrete floors, constrains the pigs to expend more ME to compensate. Poor-quality air with high levels of harmful gases, such as ammonia or hydrogen sulfide, or dust can lead to respiratory issues and energy expenditure for immune defense.
What are the detailed consequences?
Energy required for immune defense cannot be used for the production of meat, milk, or eggs. Several energy-consuming processes are triggered during an immunological challenge.
Glucose, an important energy source
Several scientists (Spurlock, 1997; Rigobelo and Ávila, 2011) have stated that glucose is primarily used to meet the increased energy demands of an activated immune system. According to Kvidera et al. (2017), the reason might be that stimulated leucocytes change their metabolism from oxidative phosphorylation to aerobic glycolysis (Palsson-McDermott and O’Neill, 2013). A trial conducted by Kvidera et al. (2017) confirmed the high need for glucose. In their trial with E. coli LPS-challenged crossbred gilts, they measured the amount of glucose required to maintain normal blood glucose levels (euglycemia). They calculated that an acutely and intensely activated immune system requires 1.1 g of glucose/kg body weight0.75/h. As they obtained similar results in ruminants (Kvidera et al., 2016 and 2017), they regard this glucose requirement as conserved across species and physiological states. In a confirming study, McGilvray and coworkers (2018) observed a significant (P<0.01) decrease in blood glucose in pigs after injection of E. coli LPS.
A further energy-consuming process is the increase in body temperature (fever): To increase body temperature by 1°C, the metabolic rate must be raised by 10-12.5% (Evans et al., 2015).
Influence on protein metabolism
Stimulation of the immune system in growing pigs may lead to a redistribution of amino acids from protein retention to immune defense. Amino acids are needed as a ‘substrate’ to synthesize immune system metabolites, such as acute-phase proteins (e.g., haptoglobin, a-fibrinogen, antitrypsin, lipopolysaccharide-binding protein, C-reactive protein, and others (Rakhshandeh and De Lange, 2011)), immunoglobulins, and glutathione (Reeds and Jahoor, 2001). This impacts the requirements for amino acids quantitatively but also qualitatively, i.e., the amino acid profile. Various studies indicated an increased need for Methionine, cysteine, branched-chain amino acids (BCAAs), aromatic amino acids, Threonine, and Glutamine during immune system stimulation (Reeds et al., 1994; Melchior et al., 2004; Calder et al., 2006; Rakhshandeh and de Lange, 2011; Rakhshandeh et al., 2014).
If the required amino acids are not available, they must be either synthesized or obtained from body protein. This costs energy, leads to muscle mass degradation, and causes an imbalance in amino acid levels. Excess amino acids are catabolized, resulting in an increase in blood urea nitrogen (BUN). McGilvray et al. (2018), e.g., observed a 25% increase in BUN in their study, in which they stimulated pigs’ immune systems with LPS.
Another possibility is using amino acids as energy sources. L-Glutamine, for example, is a crucial energy source for immune cells and the primary energy substrate for mucosal cells (Mantwill, 2025).
Carcass and meat quality
As already mentioned, immune stimulation or disease leads to protein degradation. Plank and Hill (2000) reported a loss of up to 20% of body protein (mainly skeletal muscle) in critically ill humans over 3 weeks. This protein degradation influences carcass yield and quality by reducing the amount of muscle meat.
Another effect is a decrease in the muscle cross-sectional area of fibers and a significant shift from the myosin heavy chain (MHC)-II towards the MHC-I type (Gilvray et al, 2019)
How can feed additives support pigs in health challenges?
Health challenges can occur due to infections by bacteria, viruses, fungi, or protozoa, as well as due to myco-, exo-, or endotoxins. Phytomolecules-based and toxin-binding can help animals cope with these health challenges.
Phytomolecules have several health-supporting effects
Phytomolecules can support animals in the case of a health challenge by directly fighting bacteria – antimicrobial effect (Burt, 2004; Rowaiye et al., 2025), scavenging free radicals – antioxidant effect (Saravanan et al., 2025; Dhir, 2022), or mitigating infection – anti-inflammatory effect (Saravanan et al., 2025).
A trial with the phytomolecules-based product Ventar D demonstrated its antimicrobial and microbiome-modulating effects (Heinzl, 2022). The product clearly reduced the populations of Salmonella enterica, E. coli, and Clostridium perfringens but spared the beneficial lactobacilli.
The anti-inflammatory effects of phytomolecules inhibit the activity of pro-inflammatory cytokines and chemokines from endotoxin-stimulated immune cells and epithelial cells (Lang et al., 2004; Lee et al., 2005; Liu et al., 2020), and there is an indication that the anti-inflammatory effects might be mediated by blocking the NF-κB activation pathway (Lee et al., 2005). A trial confirmed this thesis by showing a dose-dependent reduction of NFκB activity in LPS-stimulated mouse cells (-11% & -54% with 50 & 200 ppm Ventar D, respectively) (Figure 1).
Figure 1: NFκB activity in LPS-stimulated mouse cells with different inclusion rates of Ventar D (light color: no LPS; dark color: 0.25 µg LPS/mL)
Additionally, Ventar D increases interleukin-10, a cytokine with anti-inflammatory properties, and decreases interleukin-6, a pro-inflammatory cytokine. The result is a dose-dependent decline in the ratio of IL-6 to IL-10 (Figure 2), indicating the effectiveness of the product.
Figure 2: IL-6/IL-10 ratio
The effects of Ventar D, which support the immune system and redirect energy to enhance growth performance, result in higher daily gains and improved feed conversion. This was observed in a trial conducted on a commercial farm in Germany, using, on average, 26-day-old weaned piglets with a mean body weight of approximately 8 kg. Just after weaning, young animals experience stress (new feed, new groups, and separation from the dam) and are more susceptible to disease.
Two groups of piglets were fed either the regular feed of the farm (Control) or the regular feed + 100 g Ventar per MT of feed. The results for final weight and FCR are shown in Figures 3 and 4
Figure 3: Final weight in weaned piglets with and without Ventar D
Figure 4: FCR in weaned piglets with and without Ventar D
Toxin-binding products support animals against health challenges caused by toxins
As mentioned, various toxins, including myco-, endo-, and exotoxins, can harm animals. The danger of mycotoxins lurks in many feeds, and exo- and endotoxins derive from bacteria. Toxin-binding products, possibly supplemented with phytomolecules that support health (e.g., liver protection), can help animals cope with these challenges.
Solis Max 2.0, a toxin solution containing bentonite and phytomolecules, showed excellent binding performance for myco- and endotoxins (Figures 5 and 6).
Trial with endotoxins
Two samples were prepared: one with only 25 EU (1 EU equivalent to approximately 100 pg or 10,000 cells) of LPS of E. coli O55:B5 LPS/mL solution, and one with the same concentration of LPS but also containing 700 mg Solis Max 2.0/mL.
Solis Max 2.0 bound about 80% of endotoxin.
Figure 5: Endotoxin-binding capacity of Solis Max
Trial with mycotoxins
In another in vitro trial, the binding capacity of Solis Max 2.0 for six different kinds of mycotoxins was evaluated. For that purpose, samples with 800 ppb AFB1, 400 ppb OTA, 800 ppb DON, 300 ppb T2, 2,000 ppb FB1, or 1,200 ppb ZEN were prepared, and Solis max was added at two inclusion rates, one corresponding to 1 kg/t, the other to 2 kg/t. The binding capacities ranged from 40.7% for OTA to 96% for AFB1, with the lower inclusion rate, and from 61.5% for OTA to 99% for AFB1, with the higher inclusion rate.
Figure 6: Mycotoxin-binding capacity of Solis Max
Health support by toxin-binding solutions improves performance
The mitigating effects of Solis Max concerning the negative impact of toxins are also reflected in performance. A trial involving 24 female weaned piglets was conducted to evaluate the mitigating effects of Solis Max in the event of a challenge with a naturally contaminated diet (3,400 ppb of DON and 700 ppb of ZEA). Solis Max was added to one half of the challenged piglets. The addition of Solis Max to the contaminated diet not only compensates for growth performance parameters, such as weight gain and feed conversion, but also for Vulva and tail necrosis scores. The results are shown in Figures 7-11.
Figure 7: Feed intake (g)
Figure 8: Body weight gain (g)
Figure 9: FCR
Figure 10: Vulva score
Figure 11: Tail necrosis score
Tools are available to prevent the unnecessary expenditure of energy for immune protection
As the various references in the article demonstrate, health challenges such as pathogens or toxins not only spoil the appetite of animals but also require energy due to the activation of the immune system. Products based on phytomolecules, as well as toxin solutions, can help animals cope with these challenges and conserve energy for improved performance.
References:
Balli, Swetha, Karlie R. Shumway, and Shweta Sharan. “Physiology, Fever.” StatPearls [Internet]., September 4, 2023. https://www.ncbi.nlm.nih.gov/books/NBK562334/.
Burt, Sara. “Essential Oils: Their Antibacterial Properties and Potential Applications in Foods—a Review.” International Journal of Food Microbiology 94, no. 3 (August 2004): 223–53. https://doi.org/10.1016/j.ijfoodmicro.2004.03.022.
Calder, Phillip C. “Branched-Chain Amino Acids and Immunity ,.” The Journal of Nutrition 136, no. 1 (January 2006). https://doi.org/10.1093/jn/136.1.288s.
Dhir, Vivek. “Emerging Prospective of Phytomolecules as Antioxidants against Chronic Diseases.” ECS Transactions 107, no. 1 (April 24, 2022): 9571–80. https://doi.org/10.1149/10701.9571ecst.
Evans, Sharon S., Elizabeth A. Repasky, and Daniel T. Fisher. “Fever and the Thermal Regulation of Immunity: The Immune System Feels the Heat.” Nature Reviews Immunology 15, no. 6 (May 15, 2015): 335–49. https://doi.org/10.1038/nri3843.
Heinzl, Inge. “Efficient Microbiome Modulation with Phytomolecules.” EW Nutrition, June 9, 2023. https://ew-nutrition.com/pushing-microbiome-in-right-direction-phytomolecules/.
Huntley, Nichole F., John F. Patience, and C. Martin Nyachoti. “Immune Stimulation UPS Maintenance Energy Requirements.” National Hog Farmer.com, September 28, 2017. https://www.nationalhogfarmer.com/hog-health/immune-stimulation-ups-maintenance-energy-requirements.
Kvidera, S. K., E. A. Horst, M. Abuajamieh, E. J. Mayorga, M. V. Sanz Fernandez, and L. H. Baumgard. “Technical Note: A Procedure to Estimate Glucose Requirements of an Activated Immune System in Steers.” Journal of Animal Science 94, no. 11 (November 1, 2016): 4591–99. https://doi.org/10.2527/jas.2016-0765.
Kvidera, S.K., E.A. Horst, M. Abuajamieh, E.J. Mayorga, M.V. Sanz Fernandez, and L.H. Baumgard. “Glucose Requirements of an Activated Immune System in Lactating Holstein Cows.” Journal of Dairy Science 100, no. 3 (March 2017): 2360–74. https://doi.org/10.3168/jds.2016-12001.
LANG, A. “Allicin Inhibits Spontaneous and Tnf-$alpha; Induced Secretion of Proinflammatory Cytokines and Chemokines from Intestinal Epithelial Cells.” Clinical Nutrition, May 2004. https://doi.org/10.1016/s0261-5614(04)00058-5.
Lee, Seung Ho, Sun Young Lee, Dong Ju Son, Heesoon Lee, Hwan Soo Yoo, Sukgil Song, Ki Wan Oh, Dong Cho Han, Byoung Mog Kwon, and Jin Tae Hong. “Inhibitory Effect of 2′-Hydroxycinnamaldehyde on Nitric Oxide Production through Inhibition of NF-ΚB Activation in RAW 264.7 Cells.” Biochemical Pharmacology 69, no. 5 (March 2005): 791–99. https://doi.org/10.1016/j.bcp.2004.11.013.
Liu, S. D., M. H. Song, W. Yun, J. H. Lee, H. B. Kim, and J. H. Cho. “Effect of Carvacrol Essential Oils on Growth Performance and Intestinal Barrier Function in Broilers with Lipopolysaccharide Challenge.” Animal Production Science 60, no. 4 (January 22, 2020): 545–52. https://doi.org/10.1071/an18326.
Liu, S. D., M. H. Song, W. Yun, J. H. Lee, H. B. Kim, and J. H. Cho. “Effect of Carvacrol Essential Oils on Growth Performance and Intestinal Barrier Function in Broilers with Lipopolysaccharide Challenge.” Animal Production Science 60, no. 4 (January 22, 2020): 545–52. https://doi.org/10.1071/an18326.
Mantwill, Elke. “Eiweiß & Immunsystem.” sportärztezeitung, April 10, 2025. https://sportaerztezeitung.com/rubriken/ernaehrung/9197/eiweiss-immunsystem/.
McGilvray, Whitney D, David Klein, Hailey Wooten, John A Dawson, Deltora Hewitt, Amanda R Rakhshandeh, Cornelius F de Lange, and Anoosh Rakhshandeh. “Immune System Stimulation Induced byEscherichia ColiLipopolysaccharide Alters Plasma Free Amino Acid Flux and Dietary Nitrogen Utilization in Growing Pigs1.” Journal of Animal Science 97, no. 1 (October 11, 2018): 315–26. https://doi.org/10.1093/jas/sky401.
Melchior, D., B. Sève, and N. Le Floc’h. “Chronic Lung Inflammation Affects Plasma Amino Acid Concentrations in Pigs.” Journal of Animal Science 82, no. 4 (April 1, 2004): 1091–99. https://doi.org/10.2527/2004.8241091x.
Obled, C. “Amino Acid Requirements in Inflammatory States.” Canadian Journal of Animal Science 83, no. 3 (September 1, 2003): 365–73. https://doi.org/10.4141/a03-021.
Palsson‐McDermott, Eva M., and Luke A. O’Neill. “The Warburg Effect Then and Now: From Cancer to Inflammatory Diseases.” BioEssays 35, no. 11 (September 20, 2013): 965–73. https://doi.org/10.1002/bies.201300084.
Pastorelli, H., J. van Milgen, P. Lovatto, and L. Montagne. “Meta-Analysis of Feed Intake and Growth Responses of Growing Pigs after a Sanitary Challenge.” Animal 6, no. 6 (2012): 952–61. https://doi.org/10.1017/s175173111100228x.
Patience, John. “One of the Most Important Decisions in Swine Production: Dietary Energy Level – Dr. John Patience by The Swine It Podcast Show.” Spotify for Creators, December 2, 2019. https://anchor.fm/swineitpodcast/episodes/One-of-the-most-important-decisions-in-swine-production-dietary-energy-level—Dr–John-Patience-e99j9u.
Plank, Lindsay D., and Graham L. Hill. “Sequential Metabolic Changes Following Induction of Systemic Inflammatory Response in Patients with Severe Sepsis or Major Blunt Trauma.” World Journal of Surgery 24, no. 6 (June 2000): 630–38. https://doi.org/10.1007/s002689910104.
Rakhshandeh, A., and C.F.M. de Lange. “Evaluation of Chronic Immune System Stimulation Models in Growing Pigs.” Animal 6, no. 2 (2012): 305–10. https://doi.org/10.1017/s1751731111001522.
Rakhshandeh, A., and C.F.M. De Lange. “Immune System Stimulation in the Pig: Effect on Performance and Implications for Amino Acid Nutrition.” Essay. In Manipulating Pig Production XIII, 31–46. Werribee, Victoria, Australia: Australasian Pig Science Association Incorporation, 2011.
Rakhshandeh, Anoosh, John K. Htoo, Neil Karrow, Stephen P. Miller, and Cornelis F. de Lange. “Impact of Immune System Stimulation on the Ileal Nutrient Digestibility and Utilisation of Methionine plus Cysteine Intake for Whole-Body Protein Deposition in Growing Pigs.” British Journal of Nutrition 111, no. 1 (January 14, 2014): 101–10. https://doi.org/10.1017/s0007114513001955.
Reeds, P., and F. Jahoor. “The Amino Acid Requirements of Disease.” Clinical Nutrition 20 (June 2001): 15–22. https://doi.org/10.1054/clnu.2001.0402.
Reeds, Peter J, Carla R Fjeld, and Farook Jahoor. “Do the Differences between the Amino Acid Compositions of Acute-Phase and Muscle Proteins Have a Bearing on Nitrogen Loss in Traumatic States?” The Journal of Nutrition 124, no. 6 (June 1994): 906–10. https://doi.org/10.1093/jn/124.6.906.
Rigobelo, E. Cid, and F. A. De Ávila. “Hypoglycemia Caused by Septicemia in Pigs.” Essay. In Hypoglycemia – Causes and Occurrences., 221–38. London, UK: InTechOpen, 2011.
Rowaiye, Adekunle, Gordon C. Ibeanu, Doofan Bur, Sandra Nnadi, Ugonna Morikwe, Akwoba Joseph Ogugua, and Chinwe Uzoma Chukwudi. “Phyto-Molecules Show Potentials to Combat Drug-Resistance in Bacterial Cell Membranes.” Microbial Pathogenesis 205 (August 2025): 107723. https://doi.org/10.1016/j.micpath.2025.107723.
Saravanan, Haribabu, Maida Engels SE, and Muthiah Ramanathan. “Phytomolecules Are Multi Targeted: Understanding the Interlinking Pathway of Antioxidant, Anti Inflammatory and Anti Cancer Response.” In Silico Research in Biomedicine 1 (2025): 100002. https://doi.org/10.1016/j.insi.2025.100002.
Spurlock, M E. “Regulation of Metabolism and Growth during Immune Challenge: An Overview of Cytokine Function.” Journal of Animal Science 75, no. 7 (1997): 1773–83. https://doi.org/10.2527/1997.7571773x.
Suchner, U., K. S. Kuhn, and P. Fürst. “The Scientific Basis of Immunonutrition.” Proceedings of the Nutrition Society 59, no. 4 (November 2000): 553–63. https://doi.org/10.1017/s0029665100000793.
A holistic approach to animal health and nutrition: From feed issues to intestinal permeability – A conversation in Berlin
Recently, The Poultry Site’s Sarah Mikesell interviewed Predrag Persak, EW Nutrition’s Regional Technical Manager for Northern Europe. The conversation covered topics as wide as sustainability and challenges in poultry production, and as narrow as intestinal permeability. Thanks to The Poultry Site for the great talk!
Sarah Mikesell, The Poultry Site: Hi, this is Sarah Mikesell with The Poultry Site, and today we are here with Predrag Peršak. He is the Regional Technical Manager for Northern Europe with EW Nutrition. Thanks for being with us today, Predrag.
Predrag Peršak, EW Nutrition: Nice to be here, Sarah. Thank you for inviting me.
SM: Very good. It’s nice to visit with you. And today, Predrag and I are in Berlin, Germany, at an exclusive event for the poultry industry called Producing for the Future, which is sponsored by EW Nutrition. You are one of our speakers today, Predrag, so I’m going to ask you just a few questions to let everybody know a little bit about your presentation.
You’ve described animal nutrition as “never boring and never finished.” What makes this field so dynamic and constantly evolving for you?
PP: I’ve been in animal nutrition for about 25 years. And in those 25 years, I would say that not even half a year passed without something extraordinary happening. From genetics to animal husbandry, especially here in Europe, we also have a lot of pressure from consumers and slaughterhouses to adapt production to the needs of the customers.
Sustainability, sourcing raw materials, and the variety of raw materials available in Europe – and the constant development of new ones – make life for an animal nutritionist very, very interesting. It’s also very challenging, and through these challenges you learn a lot.
So, applying what we learned 20 years ago is simply not enough anymore. For someone who wants to be challenged every day with new things, this is definitely the right industry to be in – especially now.
SM: Excellent. Can you explain your holistic approach to animal nutrition and how considering multiple factors benefits practical applications on farms?
PP: The concept of a holistic approach in animal nutrition is not new. But for me – being both a veterinarian and a nutritionist – it means having deeper insight into the animal itself, into all the metabolic processes, and also into the external influences: husbandry, genetics, diseases, and management. Looking at how all of these interact, we can only really solve problems by looking at the animal as a whole system.
The same applies to feed production. You cannot look at a feed mill as just one compartment. You have to look at sourcing raw materials, their quality, how they are processed – milling, pelleting, and other technologies – and then see how that feed performs on the farm.
So, a holistic approach can be applied both from the animal perspective and from the feed production perspective, across all steps and processes. This is something we use and promote daily in our work with customers.
SM: Very good. You’ve worked with unconventional protein and fiber sources. We’re hearing a lot more about that recently. What are those, and what potential do they bring to animal nutrition?
PP: When I talk about unconventional protein and fiber sources, we need to remember that the global feed production scene is very diverse. What applies in the U.S. or Brazil does not necessarily apply in Europe or the Far East.
Here in Europe, we try to use not by-products but co-products of food production. For example, different fractions of rapeseed or sunflower meal, which are widely produced in Europe but not often used by mainstream nutritionists due to certain limitations. By finding the right processing methods and combining them with technologies, we can make these unconventional materials usable in mainstream nutrition.
The same goes for fiber sources. Both fermentable and structural fibers are increasingly important for intestinal and digestive development, as well as for overall animal health. So, processing fibers in ways that maximize usability while minimizing negative effects is a big part of my work.
SM: From a cost standpoint for producers, are those lower-cost inputs, or just alternatives they need to look at?
PP: In Germany we have a perfect expression for this: “yes and no.” There is always pressure on price, especially in poultry, because food must be accessible to everyone. But at the same time, food must not harm the environment or human health, and we should use all resources not fit for humans but still usable for animals.
So, it’s not only about cost – about availability and sustainability. Working with just two, three, or five raw materials for a long time is not the way forward. The way forward is to think of everything that can be used properly, for the benefit of the animals, and ultimately to produce enough food for the world.
Also, using locally available products is important. Feed production is very diverse around the world—raw materials in Southeast Asia differ completely from those in Europe, Brazil, or the U.S. Using technologies to enable the use of locally produced by-products makes production not only sustainable, but also economically viable for local communities. That’s really the core of the feed industry: using what is produced locally.
SM: Interesting. Very cool. How does your interdisciplinary work across poultry, pigs, and ruminants give you unique insights that might be missed with a narrower focus?
PP: I come from a small feed mill in a small country, Croatia. There, you don’t have deep specialization by species or even by category, as you find in larger markets. Specialization has its advantages, but it can also limit creativity and “outside-the-box” thinking.
By working with ruminants, I learned about fermentation processes – knowledge that can be applied to pigs and even to poultry. For example, fermentation can reduce anti-nutritional factors, allowing higher inclusion levels of certain raw materials in poultry diets.
With pigs, fermentation of fibers – especially in piglets – is crucial, and some of that knowledge could be applied to turkeys, where we still face health issues.
So, working across species demands a lot – it leaves little time for other things – but it opens up unique perspectives and cross-species applications that benefit the entire livestock industry.
SM: I was talking with someone yesterday about mycotoxins – there’s a lot of research in pigs but less in poultry. That’s kind of what you’re talking about, right? Applying knowledge across species?
PP: Absolutely. We’re focused now on poultry, but we can learn from poultry too – not only about feeding but also about farm management, biosecurity, and more. These lessons can also apply to pigs or ruminants.
It’s all holistic – you cannot solve everything with nutrition alone. It’s always a package.
SM: You presented today about the importance of intestinal permeability. Why is it important, and how can understanding it impact animal health and performance outcomes?
PP: Intestinal permeability is one of the key features we use to describe gut health. Personally, I’m very practical. For 20 years we’ve talked about “gut health,” but the real question for veterinarians and nutritionists is: what do we actually do with that knowledge?
In my presentation, I explained intestinal permeability as a “point of no return” in gut health. When leaky gut develops, everything else can deteriorate – faster or slower – but it won’t return to normal without intervention.
By comparing how different stressors or pathogens impact intestinal permeability, we can better understand severity and decide where to focus. Nutritionists already pay attention to thousands of factors, but we need to identify the most impactful ones. That was my key message: focus on the most important drivers.
SM: And leaky gut has really become something the whole industry is talking about, right? I’ve even seen it in human health – my doctor has posters about it.
PP: Exactly. Across cows, pigs, and poultry, leaky gut is getting a lot of attention. It’s a physiological or pathophysiological feature that marks the point of no return.
We can talk about dysbiosis and all the causes, but once you reach leaky gut, you understand where intervention is needed. And it’s not just hype. For example, recently Nature published research showing certain types of human bone marrow conditions are linked to leaky gut and microbial influence on blood processes.
So, this is not a passing trend. It’s fundamental. And once we solve one issue, another door opens. That’s why this industry is never boring.
SM: Very good. Well, thank you for all the information today, Predrag.
PP: Thank you, Sarah. It was a pleasure to talk with you.
Optimizing the Use of DDGS in Poultry Feeds with Xylanase
Author: Ajay Bhoyar, Sr. Global Technical Manager, EW Nutrition
As the poultry industry seeks economical and nutritious feed ingredients, distillers’ dried grains with solubles (DDGS), a co-product of grain-based ethanol production, presents a valuable option providing beneficial protein, energy, water-soluble vitamins, xanthophylls, and linoleic acid. However, the inherent variability in DDGS nutrient composition and high fiber content can pose challenges for consistent inclusion in poultry feeds. The strategic use of feed enzymes has become a significant area of focus to overcome these limitations and further enhance the nutritional value of DDGS in poultry diets. This article will explore the optimization of DDGS utilization in poultry feeds by emphasizing the inclusion of xylanase enzyme that can efficiently degrade the insoluble arabinoxylans. By understanding the factors affecting DDGS quality and strategically employing xylanase, poultry producers can potentially achieve higher inclusion rates of this readily available byproduct, aiming to reduce feed costs while maintaining or even improving production performance and overall health.
Price competitiveness of DDGS
The price of DDGS relative to other feed ingredients, primarily corn and soybean meal, is a significant factor in its global utilization. DDGS often partially replaces these traditional energy (corn) and protein (soybean meal) sources in animal feeds, leading to significant diet cost savings for poultry producers. DDGS contains a high amount of a combination of energy, amino acids, and phosphorus. However, it is usually undervalued as its price is mainly determined based on the prevailing prices of corn and soybean meal.
Variability in the nutritional quality of DDGS
The nutrient composition of DDGS varies based on the starting grain, ethanol production methods, and drying processes. Generally, DDGS contains high levels of protein, fiber, and minerals, with varying amounts of fat and starch depending on the type of grain used and how it is processed. DDGS has a reputation for having variable nutrient composition, protein quality, and a high content of mycotoxins (Stein et al., 2006; Pedersen et al., 2007; Anderson et al., 2012). High quantities of DDGS in feed increase dietary fiber, adversely affecting nutrient digestibility.
The variations in production methods lead to significant differences in the following nutritional components of DDGS:
Crude Fat: This is one of the most variable components, ranging from 5 to 9 percent in reduced-oil DDGS and greater than 10 percent in traditional high-oil DDGS.
Energy: The apparent metabolizable energy (AMEn) for poultry varies among DDGS sources. Fiber digestibility and the digestibility of the extracted oil also contribute to this variability. The high temperatures during the drying stage of DDGS production accelerate lipid peroxidation, forming breakdown products from the fats. This peroxidation contributes to the changes and variability observed in the fat component of DDGS and is a factor that can affect nutrient digestibility and overall energy value.
Crude Protein and Amino Acids (especially Lysine): While crude protein content might not always increase inversely with fat reduction, the digestibility of amino acids, especially lysine, can be affected by drying temperatures. Lysine digestibility of DDGS is a primary concern of poultry nutritionists due to the susceptibility of this amino acid to Maillard reactions during the drying process of DDGS, which can reduce both the concentration and digestibility of lysine (Almeida et al. 2013). Prediction equations have been developed to accurately estimate actual AMEn and standardized ileal digestible amino acid content of DDGS sources based on chemical composition.
Phosphorus: The phosphorus content can vary depending on the amount of Condensed Distiller’s Solubles (CDS) added. The bioavailability of phosphorus can also be influenced by processing. The phosphorus content in the corn DDGS may vary from 0.69 to 0.98 % (Olukosi and Adebiyi, 2013).
Fiber: The neutral detergent fiber (NDF) content is another variable component. Differences in processing conditions among ethanol plants can lead to variations in fiber digestibility.
Table 1. Variation in composition of corn DDGS sources (dry matter basis; adapted from (Pederson et al., 2014)
Analyte
Average
Range
Moisture %
8.7
6.5 – 12.5
Crude protein %
31.4
27.1 – 36.4
Crude fiber %
7.7
6.4 – 9.5
Ether Extract %
9.1
6.5 – 11.8
NDF %
35.1
30.2 – 39.7
ADF %
10.1
8.9 – 11.9
Nonstarch Polysaccharides (NSP) in DDGS
Non-starch polysaccharides (NSP) are a significant component of DDGS. The NSP profile of DDGS is crucial for understanding its digestibility and energy content. The corn DDGS has a complex fiber structure that may limit its digestibility in swine and poultry. NSPs in corn DDGS represent 25-34% of its composition, primarily insoluble (Pedersen et al. 2014). The complexity of the fiber structure in corn DDGS makes it more challenging to degrade with enzymes than wheat DDGS. Therefore, while including DDGS in the poultry feeds, choosing an exogenous xylanase enzyme that is highly efficient in breaking down both soluble and insoluble arabinoxylans is essential for maximum energy utilization.
Use of xylanase in DDGS diets for poultry
Supplementing exogenous enzymes in swine and poultry diets have numerous potential benefits including: reduction of digesta viscosity to enhance lipid and protein digestion; increase the metabolizable energy content of the diet; increase feed intake, growth rate and feed conversion; decreased size and alter the microbial population of the gastrointestinal tract; reduce water consumption and water content of excreta in poultry; reduce the amount of excreta as well as ammonia, nitrogen and phosphorus content (Khattak et al., 2006). The selection of a specific enzyme must be based on the type and availability of the target substrate in the diet.
The improved energy utilization of DDGS in poultry can be achieved through the enzymatic degradation of fiber (NSP). Nonstarch polysaccharides within DDGS exist in matrices with starch and protein, so NSP degradation via exogenous enzymes can also release other nutrients for subsequent digestion and absorption (Jha et al. 2015).
The cell wall matrix in corn DDGS is more complex. Moreover, the most readily degradable arabinoxylan for the fiber-degrading enzymes is modified during DDGS production (Pedersen et al. 2014). Many studies reported a greater branch density and complexity of corn arabinoxylan than wheat (Bedford, 1995; Saulnier et al.,1995a; Jilek and Bunzel, 2013; Yang et al., 2013). These observations indicate that the fiber-degrading enzymes applied for the degradation of corn DDGS need to be targeted towards highly complex substrates. This calls for selecting xylanase, which effectively breaks down the insoluble arabinoxylans in diets.
Axxess XY: Highly effective xylanase in breaking down soluble and insoluble arabinoxylans
A bacterial GH10 family xylanase, like Axxess XY, is more beneficial in animal production due to their efficient mechanism of action, broader substrate specificity, and better thermostability. Generally, the GH10 xylanases exhibit broader substrate specificity and can efficiently hydrolyze various forms of xylan, including soluble and insoluble substrates. GH10 xylanases exhibit higher catalytic versatility and can catalyze the cleavage of the xylan backbone at the non-reducing side of substituted xylose residues, whereas GH11 enzymes require unsubstituted regions of the xylan backbone (Collins et al., 2005; Chakdar et al., 2016).
Fig.1. Activity of a bacterial GH10 xylanase against soluble and insoluble arabinoxylans
Axxess XY facilitates DDGS use and reduces the cost of broiler production.
Including xylanase enzyme, which is highly effective in breaking down soluble and insoluble arabinoxylans in poultry feeds, can reduce feed costs, allowing higher inclusion of DDGS while maintaining the bird’s commercial performance.
In a recently conducted 42-day trial at a commercial farm, Axxess XY maintained broiler performance with a 100 kcal/kg reduction in metabolizable energy and 8% use of Corn DDGS in a corn-SBM based diet (Figure 2). This significantly reduced feed cost/kg body weight.
Incorporating DDGS into poultry diets presents a sustainable and cost-effective solution, but its full potential is often limited by variability in nutrient composition and high fiber content. Xylanase enzymes, particularly those in the GH10 family like Axxess XY, can overcome these barriers by breaking down complex arabinoxylans and unlocking inaccessible nutrients. With proven benefits in energy utilization, nutrient digestibility, and overall production efficiency, xylanase inclusion emerges as a strategic approach to optimize DDGS usage, ultimately supporting economic and environmental sustainability goals in poultry production.
References
Almeida, F.N.; Htoo, J.K.; Thomson, J.; Stein, H.H. Amino acid digestibility of heat-damaged distillers’ dried grains with soluble fed to pigs. J. Anim. Sci. Biotechnol. 2013, 4, 2–11.
Bedford, M.R., 1995. Mechanism of action and potential environmental benefits from the use of feed enzymes. Anim. Feed Sci. Technol. 53, 145–155.
Collins, Tony, Charles Gerday, and Georges Feller. “Xylanases, Xylanase Families and Extremophilic Xylanases.” FEMS Microbiology Reviews 29, no. 1 (January 2005): 3–23. https://doi.org/10.1016/j.femsre.2004.06.005.
Jha, R.; Woyengo, T.A.; Li, J.; Bedford, M.R.; Vasanthan, T.; Zijlstra, R.T. Enzymes enhance degradation of the fiber–starch–protein matrix of distillers dried grains with solubles as revealed by a porcine in vitro fermentation model and microscopy. J. Anim. Sci. 2015, 93, 1039–1051.
Jilek, M.L., Bunzel, M., 2013. Dehydrotriferulic and dehydrodiferulic acid profiles of cereal and pseudocereal flours. Cereal Chem. J. 90, 507–514
Jones, C.K., Bergstrom, J.R., Tokach, M.D., DeRouchey, J.M., Goodband, R.D., Nelssen, J.L., Dritz, S.S., 2010. Efficacy of commercial enzymes in diets containing various concentrations and sources of dried distillers’ grains with solubles for nursery pigs. J. Anim. Sci. 88, 2084–2091.
Khattak, F.M., T.N. Pasha, Z. Hayat, and A. Mahmud. 2006. Enzymes in poultry nutrition. J. Anim. Pl. Sci. 16:1-7.
Olukosi, O.A., and A.O. Adebiyi. 2013. Chemical composition and prediction of amino acid content of maize- and wheat-distillers’ Dried Grains with Soluble. Anim. Feed Sci. Technol. 185:182-189.
Pedersen M. B., Dalsgaard S., Bach Knudsen K.E., Yu S., Lærke H.N., Compositional profile and variation of Distillers Dried Grains with Solubles from various origins with focus on non-starch polysaccharides, Animal Feed Science and Technology, Volume 197, 2014, Pages 130–14.
Saulnier, L., Vigouroux, J., Thibault, J.-F., 1995a. Isolation and partial characterization of feruloylated oligosaccharides from maize bran. Carbohydr. Res. 272,241–253.
Yang, J., Maldonado-Gómez, M.X., Hutkins, R.W., Rose, D.J., 2013. Production and in vitro fermentation of soluble, non-digestible, feruloylated oligo- andpolysaccharides from maize and wheat brans. J. Agric. Food Chem.
Yoon, S.Y., Yang, Y.X., Shinde, P.L., Choi, J.Y., Kim, J.S., Kim, Y.W., Yun, K., Jo, J.K., Lee, J.H., Ohh, S.J., Kwon, I.K., Chae, B.J., 2010. Effects of mannanase and distillers’ dried grain with solubles on growth performance, nutrient digestibility, and carcass characteristics of grower-finisher pigs. J. Anim. Sci. 88,181–191.






")