Ionophores: An Overlooked Risk for the Spread of Medically Relevant Antibiotic Resistance
Author: Dr. Inge Heinzl, Editor EW Nutrition
Antibiotic resistance is one of the biggest threats to global health today. When bacteria become resistant to antibiotics, infections that were once easily treatable can become deadly. For decades, the discussion surrounding the causes of antimicrobial resistance (AMR) has primarily focused on the misuse of antibiotics in human medicine and agriculture. But some antibiotics have escaped critical scrutiny—until now.
Ionophores, a special group of antibiotics
Ionophores are a group of antibiotics used as feed additives in ruminants and pigs as growth promoters and in poultry as anticoccidials since the early 1970s (Chapman et al., 2010). They are among the most widely used classes of antibiotics in animal production. In the US, e.g., more than 4 million kilograms were sold in 2016 (Wong, 2019).
Unlike many other antibiotics, ionophores are not used in human medicine because of their toxicity. For this reason, regulators have often assumed that ionophores pose little to no threat to human health. In North America, for example, ionophores are officially classified as having low or no importance for human medicine, which means their use is less strictly regulated than antibiotics that are directly relevant for human health.
However, new scientific findings challenge this assumption. A research team led by Asalia Ibrahim (2025) has provided compelling evidence that the use of ionophores in agriculture may indirectly contribute to the spread of resistance to antibiotics crucial for treating human infections.
What did the researchers discover?
The researchers focused on two specific genes, narA and narB, transporters which enable Enterococcus faecium to resist ionophores like narasin, salinomycin, and maduramicin. Initially, these genes were found in bacteria isolated from Swedish broiler chickens AND on the same plasmid as vancomycin resistance genes (Nilsson et al., 2012). More recent studies have identified the NarA and NarB genes in other countries as well, raising questions about their global distribution and their connection to resistance to medically important antibiotics.
To investigate, Asalia Ibrahim (2025) analyzed publicly available genome data from the NCBI Pathogens database, a massive resource that collects bacterial genome sequences from around the world. They identified more than 2,400 bacterial isolates from 51 countries that carry both narA and narB. The bacteria were found in various host animals, including poultry, swine, and cattle, but also in humans. Alarmingly, over 500 of the samples containing these resistance genes came from human sources!
Why is this a problem?
The core concern is that these ionophore resistance genes do not exist in isolation. Instead, they are often genetically linked with other resistance genes that protect bacteria from antibiotics that are critical for human medicine.
This can happen in two ways:
Cross-resistance, where a single gene provides resistance to multiple drugs at once. In this case, it appears unlikely because ionophores belong to a class (polyether antibiotics) that is not used for humans.
Co-selection occurs when different resistance genes sit close together on the same piece of genetic material (like a plasmid) or in the same bacterial genome. If one gene is selected because the antibiotic it resists is used, then the other genes hitch a ride and spread too.
The researchers found clear evidence for co-selection. Many narAB-carrying bacteria also contained resistance genes for vancomycin, a last-resort antibiotic (Nilsson et al., 2012), erythromycin, tetracycline (Pikkemaat et al., 2022), and other antibiotics. On average, each narAB isolate carried more than 10 additional resistance determinants, including both resistance genes and mutations.
The link is not just theoretical. When the Norwegian poultry industry stopped using narasin in 2016, the levels of vancomycin-resistant Enterococcus dropped significantly (Simm et al., 2019). This real-world example suggests that the use of ionophores can indeed help maintain resistance to medically relevant antibiotics in animal populations, potentially allowing these bacteria to enter the food chain and reach humans.
What does this mean for food safety and public health?
The study’s findings highlight how actions taken in agriculture can have far-reaching effects on human health. Suppose bacteria carrying narAB genes also carry resistance to life-saving human antibiotics. In that case, the routine use of ionophores in animal feed can indirectly contribute to maintaining a reservoir of resistant genes. These bacteria can spread from animals to humans through direct contact, contaminated meat, or environmental exposure.
This raises questions about the long-held belief that ionophores are risk-free. In reality, they might be acting as a hidden driver for the maintenance and spread of resistance genes that severely limit our treatment options in human medicine.
What should be done?
The researchers argue that ionophores need to be reevaluated within the broader framework of the “One Health” approach, which recognizes that the health of people, animals, and ecosystems are deeply interconnected. Simply because ionophores are not used in hospitals does not mean they are harmless to human health.
Possible steps could include:
Stricter monitoring of ionophore use in livestock.
Better surveillance of resistance genes like narA and narB in both animal and human bacterial isolates.
Considering limits or alternatives to routine ionophore use in industrial farming.
More research to understand how these resistance genes move between bacteria, species, and environments.
The bottom line
Ionophores play a crucial role in intensive animal production worldwide, helping to maintain the health and productivity of animals. But this convenience comes at a potential cost. The research of Ibrahim et al. (2025) serves as a clear reminder that the use of antibiotics—whether for humans or animals—can have unintended consequences for global health.
Prudent, science-based management of all antibiotics is crucial to slowing the spread of antimicrobial resistance and preserving the effectiveness of life-saving drugs for future generations.
References
Chapman, H.D., T.K. Jeffers, and R.B. Williams. “Forty Years of Monensin for the Control of Coccidiosis in Poultry.” Poultry Science 89, no. 9 (September 2010): 1788–1801. https://doi.org/10.3382/ps.2010-00931.
Ibrahim, Asalia, Jason Au, and Alex Wong. “The Ionophore Resistance Genes narA and narB Are Geographically Widespread and Linked to Resistance to Medically Important Antibiotics.” mSphere, June 17, 2025. https://doi.org/10.1128/msphere.00243-25.
Nilsson, O., C. Greko, B. Bengtsson, and S. Englund. “Genetic Diversity among VRE Isolates from Swedish Broilers with the Coincidental Finding of Transferrable Decreased Susceptibility to Narasin.” Journal of Applied Microbiology 112, no. 4 (March 5, 2012): 716–22. https://doi.org/10.1111/j.1365-2672.2012.05254.x.
Pikkemaat, M.G., M. Rapallini, J.H.M. Stassen, M. Alewijn, and B.A. Wullings. “Ionophore Resistance and Potential Risk of Ionophore Driven Co-Selection of Clinically Relevant Antimicrobial Resistance in Poultry.” Food Safety Report, Wageningen, 2022. https://doi.org/10.18174/565488.
Simm, Roger, Jannice Schau Slettemeås, Madelaine Norström, Katharine R. Dean, Magne Kaldhusdal, and Anne Margrete Urdahl. “Significant Reduction of Vancomycin Resistant E. Faecium in the Norwegian Broiler Population Coincided with Measures Taken by the Broiler Industry to Reduce Antimicrobial Resistant Bacteria.” PLOS ONE 14, no. 12 (December 12, 2019). https://doi.org/10.1371/journal.pone.0226101.
Wong, Alex. “Unknown Risk on the Farm: Does Agricultural Use of Ionophores Contribute to the Burden of Antimicrobial Resistance?” mSphere 4, no. 5 (October 30, 2019). https://doi.org/10.1128/msphere.00433-19.
The lessons of 2025 for poultry and feed producers
by Ilinca Anghelescu, Global Director Marketing & Communications, EW Nutrition
2025 was a year defined by four converging forces for the global feed and animal production industry: an unprecedented HPAI crisis that cost American consumers alone $14.5 billion extra in egg expenditures; historic record corn production driving feed ingredient prices lower; a highly disruptive US tariff regime that reshuffled global trade flows for soybeans, corn, chicken, and pork; and accelerating regulatory pressure on antimicrobial use across Europe and globally.
The strategic imperatives from 2025 are clear: biosecurity investment is no longer optional, ingredient price volatility demands agile procurement strategies, trade compliance is a weekly operational concern, and antibiotic-free production transitions require credible, phased plans now.
KEY METRIC: Global chicken meat production reached approximately 105 million MT in 2025 (+2%), even as egg production suffered severely. The global feed market is valued at $542 billion in 2025, growing at 3.3% CAGR. Corn hit record production of 17 billion bushels in the US alone – the highest since 1936 in terms of harvested area.
The H5N1 clade 2.3.4.4b strain of Highly Pathogenic Avian Influenza (HPAI) continued to dominate animal health headlines in 2025. Since its reemergence in February 2022, the US outbreak alone has resulted in the confirmed loss of over 175 million birds across 1,700+ flocks – the costliest poultry disease event in recorded history.
Metric
Data Point
Source
Total US birds affected (2022–2025)
175+ million
USDA APHIS, May 2025
US flocks confirmed positive
1,704+
USDA APHIS, May 2025
Proportion of affected birds: layers
75%
USDA / Congressional Research Service
US egg layer flock deficit vs. 2022
–8% fewer birds
CoBank / USDA
Consumer egg overspend (May 2024–Apr 2025)
$14.5 billion extra
Innovate Animal Ag analysis
Peak US retail egg price
$6.23/dozen (March 2025)
BLS / USDA
HPAI-related US taxpayer response costs
$1.8 billion+
Innovate Animal Ag
Global HPAI mammal outbreaks (2024)
1,022 (vs. 459 in 2023)
WOAH 2025
Countries self-declaring HPAI freedom (May 2025)
25
WOAH
1.2 2025-Specific Developments
United States: Early-Year Severity, Policy Response
The first six weeks of 2025 saw 28 million layers depopulated – the worst start to any calendar year on record. Ohio, Indiana, and Missouri bore the brunt. The USDA launched a five-pronged approach in February 2025 including:
Gold-standard biosecurity assessments (948 completed Jan 20–June 26)
Indemnity increase from $7 to $17 per lost layer hen
Importation of 26+ million dozen shell eggs from Brazil, Honduras, Mexico, Turkey, and South Korea
Removal of select regulatory burdens to accelerate flock repopulation
$793 million in HPAI research proposals received in response to USDA Innovation Grand Challenge
⚠ Price Manipulation Investigation: In April 2025, the DOJ Antitrust Division launched an investigation into the largest US egg producer after it reported a 247% increase in quarterly net income. Egg producers and retailers face ongoing scrutiny over whether crisis pricing exceeded what supply constraints warranted.
Brazil: First Commercial HPAI Outbreak – May 2025
On May 15, 2025, Brazil – the world’s largest poultry exporter, responsible for nearly 30% of global exports – confirmed its first-ever commercial HPAI case at a breeder facility in Montenegro, Rio Grande do Sul (17,000 birds). This was a watershed event for global poultry trade.
Consequence
Detail
China (#1 buyer of Brazilian chicken) suspended imports
Trade suspended as of May 2025; Chinese delegation visited RS in Sept 2025 to assess resumption
Brazil’s monthly poultry exports declined
Exports fell 12.9% to $655 million; volume down 14.4% to 363,100 MT (May)
UAE replaced China as Brazil’s top buyer
First time China dropped from #1 buyer since 2019
WOAH new 10-year global HPAI strategy launched
Prevention and Control of HPAI (2024–2033), February 2025
Regionalized trade bans helped contain damage
Bans limited to affected regions, not all of Brazil
Europe: Persistent Pressure
HPAI continued to circulate widely in European poultry and wild bird populations. Key 2025 events include recurrence in Australia (February), ongoing outbreaks in Germany, Hungary, Netherlands, UK, and France, and the first confirmed domestic cat HPAI death in the Netherlands (H5N1, November 2025).
CRITICAL RISK: HPAI is now classified as enzootic (endemic) in wild birds across North America by the CDC. The virus circulates year-round in wildlife reservoirs, making seasonal recurrence in commercial flocks a structural, not episodic, risk. US egg producers are 8% below their 2022 flock baseline.
EU-wide – statistically significant increase trend 2020–2024 per EFSA/ECDC joint report, March 2025
AMR pressure in broilers and layers; genomic surveillance being mandated by EU
Newcastle Disease (NCD)
Brazil – outbreak July 2024, RS state
First commercial NCD in Brazil since 2006; adds biosecurity burden on top of HPAI protocols
H5N1 in Dairy Cattle (USA)
Ongoing – cross-species spread to 50+ US states
Cattle-to-poultry transmission confirmed; biosecurity interfaces between dairy and poultry operations must be reviewed
HPAI – Antarctica
First confirmed case March 2024 (South Polar Skua)
Indicates virus reached every continent; unprecedented in poultry disease history
CHAPTER 2: GLOBAL POULTRY PRODUCTION
2.1 Global Output – 2025 Performance
Despite HPAI disruptions, global chicken meat production grew approximately 2% in 2025 to around 105 million MT (ready-to-cook), driven by demand resilience and lower feed costs for broiler production. Total global poultry meat (including turkey, duck, and others) is forecast to exceed 152 million MT for 2025, per FAO Food Outlook June 2025.
Country / Region
2025 Production Forecast (MT)
Year-on-Year Change
Key Driver
USA – Broilers
21.7 million MT
+1.4% vs. 2024
Strong hatchery data; lower feed costs; HPAI minimal in broilers
Affordability vs. beef; consumer demand in developing markets
OECD-FAO 10-Year Outlook (2025–2034)
The OECD-FAO Agricultural Outlook 2025–2034, released in July 2025, projects global poultry meat production will grow by over 19% to 173.4 million MT by 2034 compared to the 2022–24 average. Poultry will account for the majority of additional meat consumption globally, driven by:
Affordability relative to beef and pork, especially in price-sensitive emerging markets
Population and income growth in Southeast Asia, South Asia, and Sub-Saharan Africa
Rapid urbanization and expansion of Quick Service Restaurant (QSR) chains
Superior feed conversion ratio (FCR) and lower greenhouse gas emissions per kg of protein
STRATEGIC NOTE: In high-income countries, per capita poultry consumption growth is flattening as consumers focus increasingly on welfare, environment, and health attributes. Growth opportunity is almost entirely in middle-income markets. Product premiumization (antibiotic-free, cage-free, organic) is the North American and European story.
2.2 Egg Production – Crisis Sector
Egg production was the sector hardest hit by HPAI globally. In the US, 75% of all HPAI-affected birds were table-egg layers, despite layers comprising less than 4% of the total poultry population. This structural vulnerability reflects longer flock lifespans and, increasingly, cage-free housing adoption.
Indicator
2025 Data
US retail egg price peak
$6.23/dozen (March 2025)
US retail egg price decline from peak
–27% by June 2025 (wholesale –64%)
US retail egg price (January 2025)
$4.95/dozen – 96% higher than January 2024
USDA full-year 2025 egg price forecast
+41.1% vs. 2024 average
% of US laying flock in cage-free systems
~40% (120+ million birds)
Global hen egg production (2023 baseline)
91 million tonnes (~1.7 trillion eggs)
Global egg trade volume (2024)
Nearly doubled from prior years
⚠ Cage-Free Transition & Disease Vulnerability: Some analysts link cage-free housing to higher HPAI susceptibility. Regardless of epidemiological debate, the US cage-free market is now structurally undersupplied relative to corporate commitments made in 2014–2017. Producers face a squeeze: comply with welfare commitments while managing disease risk.
CHAPTER 3: FEED INGREDIENT MARKETS
3.1 Grain & Oilseed Prices – 2025 Summary
From a feed cost perspective, 2025 was broadly favorable for livestock and poultry producers. Record US corn production and generally adequate global grain and oilseed supplies put downward pressure on the major feed commodities, offering partial relief from the margin pressure of recent years.
Commodity
2025 Price Direction
Key 2025 Data
Implication for Feed
Corn (US)
DOWN –3.9% (3rd consecutive annual decline)
Record US crop: 17.0 billion bu; yield 186.5 bu/acre – record; harvested area highest since 1936
Favorable for poultry/swine FCR cost; season avg ~$4.15/bu projected
Soybean Meal
DOWN –4.3% (3rd consecutive decline)
Prices at lowest since early 2016 at one point; large South American supply weighing on markets
Significant reduction in diet protein cost; amino acid supplementation cost-competitive
Soybeans
UP slightly +3.3%
After 22.9% collapse in 2024; still well below historical peaks; US acreage declining
Bean oil +20.8% (energy diet component); meal-to-bean ratio remains attractive for crushers
Wheat (Chicago)
DOWN –4.3% (4th consecutive year)
Abundant global supply; Russia/Argentina record crops; increased feed use
Wheat competing with corn in feed formulations globally – inclusion rising in EU/Asia diets
Soybean Oil
UP +20.8%
Driven by biofuel demand (US 45Z renewable fuel credits)
Energy ingredient cost pressure; may affect fat inclusion rates in formulations
PROCUREMENT SIGNAL: The US/China trade tensions created windows of soybean buying opportunity as prices swung on trade deal news. China agreed to purchase US soybeans in late 2025 as part of a limited trade deal, causing a price uptick. Procurement teams should monitor US-China negotiations as a lead indicator for soybean pricing in 2026.
3.2 Global Feed Market Overview
Metric
2025 Data
Global animal feed market value
$542.36 billion
CAGR (2026–2034)
3.3%
Largest feed segment by additive type
Amino acids (33.6% share)
Largest feed segment by species
Poultry (dominant share)
Asia Pacific regional status
Dominant region (largest market)
Top feed ingredient challenge
Fluctuating prices for corn, SBM – still key risk for margin management
3.3 Key Ingredient Trends to Watch
Fertilizer Cost Relief
Fertilizer prices have declined significantly from their 2022 peak. A basket of N, P, and K fertilizers averaged $437/tonne in May 2025, down from the $815/tonne peak in April 2022, per FAO Food Outlook. This benefits grain production economics and should support adequate grain supplies into 2026.
Soybean Oil Competition: Biodiesel vs. Feed
US soybean oil demand from renewable fuel programs (the 45Z credit) competed directly with feed-grade fat supplies, pushing soy oil prices up 20.8% in 2025. Feed mills formulating with added fats should evaluate alternative lipid sources. Poultry fat and palm olein remain cost-competitive in some markets.
Alternative Proteins: Insect Meal, DDGS, Algae
While adoption remains limited in volume, regulatory acceptance of insect meal in EU poultry diets continues to expand. Dried Distillers Grains with Solubles (DDGS) remain a strategically important co-product, particularly in the US and EU. Feed formulators should have up-to-date matrix values and be prepared to use them when corn prices favor inclusions.
⚠ Tariff Risk for Feed Inputs: US feed manufacturers faced effective tariff rates averaging 12%+ on key agricultural inputs from China and other countries in 2025, including herbicides, pesticides, and some micro-ingredient precursors. Amino acid supplies (predominantly Chinese-origin lysine, methionine, threonine) faced added cost and supply uncertainty.
CHAPTER 4: TRADE POLICY DISRUPTIONS
4.1 The 2025 US Tariff Regime – Agricultural Impact
The Trump administration’s tariff policies beginning January 20, 2025, represented the most significant disruption to global agricultural trade in decades. The three largest US agricultural export markets – Mexico ($30.3B in 2024), Canada ($28.3B), and China ($24.7B) – were all targeted, triggering retaliatory measures that hit feed, grain, poultry, and pork exports.
Country
US Tariff (2025)
Retaliation on US Agriculture
Key Products Impacted for Feed/Poultry Industry
China
Reached 145% (paused to 30% via May 2025 truce)
15% on chicken, corn, wheat; 10% on soybeans, sorghum, pork – applied from March 2025
Chinese poultry buyers shifted away from US; US corn/soy export disruption; amino acid supply chain uncertainty
Canada
25–35% (escalated to 35% in Aug)
25% on US dairy, poultry, meat products ($21B)
Canada imports ~45% of US poultry exports; feed grain flows affected
Mexico
25–30% (USMCA-compliant goods largely exempted)
Retaliatory tariffs threatened on agricultural goods
Mexico is #1 market for US turkey exports; ongoing uncertainty
EU
14% (paused under negotiations)
Planned retaliation announced April 2025
Potential impact on US soy meal exports; EU feed ingredient costs
CHINA TRADE DEAL (MAY 2025): A 90-day tariff truce agreed May 12, 2025 reduced US tariffs on Chinese goods from 145% to 30%, and China’s tariffs on US products from 125% to 10%. China agreed to purchase US soybeans. No permanent deal was signed. The limited agreement provided short-term stability but medium-term uncertainty remains.
4.2 Impact on US Agricultural Trade Flows
Product
Trade Flow Change (2025)
Implication
Corn exports
UP >20% YoY
Record US production driving export competitiveness despite tariff uncertainty
Soybean exports
DOWN – China shifted to South America
Brazil and Argentina taking larger share of Chinese soy imports
US chicken exports
Maintained overall (6.8B USD)
Despite China restrictions, other markets (Middle East, Mexico) absorbed volume
US turkey exports
At risk – 10% of production exported; Mexico = 65% of turkey exports
HPAI + AMPV supply squeeze threatened export volumes at peak holiday season
Brazil chicken exports
Down 12.9% month of May impact; year-end positive
HPAI disruption in May/June; recovery in H2 2025 after regionalization
US egg imports (temporary)
26M dozen shell eggs imported
Emergency imports from Brazil, Honduras, Turkey, South Korea, Mexico to fill supply gap
4.3 Strategic Trade Lessons
Supply chain diversification is no longer a luxury: concentration of US soy exports to China created a single-point-of-failure vulnerability that became fully exposed in 2025.
Regionalized disease zoning is a trade-preserving tool: Brazil’s rapid implementation of regionalized HPAI bans (rather than country-wide) preserved most of its export access; this is the model the industry should support with regulators globally.
USMCA dependency is real: 70% of US corn, 60% of soybeans, 45% of poultry exports go to Mexico, Canada, China – the same three countries targeted by 2025 tariffs.
US government announced $12B in emergency farm compensation in 2025, repeating the pattern from Trump’s first term – indicating persistent trade disruption risk.
In 2025, the European Commission proposed a package to streamline EU food and feed safety legislation while maintaining high health standards. The initiative, announced mid-2025, is intended to boost competitiveness of EU producers by reducing regulatory complexity – a direct response to competitive concerns vs. non-EU producers.
5.2 EFSA 2025 Guidance on Microorganisms
On September 24, 2025, EFSA’s Scientific Committee adopted new harmonized guidance on the characterization of microorganisms in the food chain. This is a landmark shift with major implications for feed additive manufacturers, probiotics suppliers, and novel food applicants.
Key Element
Operational Implication
Whole Genome Sequencing (WGS) now mandatory for strain-level ID of all bacteria, yeasts, fungi, viruses in applications
All existing microbial feed additive dossiers must be reviewed; WGS data cannot be more than 2 years old at time of submission
Genomics-first approach to AMR assessment
Any AMR gene hit in curated databases triggers mandatory case-by-case assessment; significantly raises the regulatory bar for probiotics and fermentation products
Replaces multiple previous guidance documents
Companies must align R&D, QC, and regulatory documentation to new unified standard immediately
GM microorganisms: clearer differentiation
Products ‘produced by GMO’ now distinguished from ‘GMO active agents’ – critical for enzyme and probiotic positioning
Non-compliance = application rejection risk
Early non-alignment causes ‘clock-stops’ or formal rejection at EFSA suitability check stage
AMR remains the defining long-term regulatory risk for the animal feed and production industry. Key 2025 actions:
EFSA/ECDC Joint Report (March 2025): Highlighted persistently high resistance to critical antimicrobials in poultry, especially Campylobacter and Salmonella, with ‘statistically significant increasing trend 2020–2024.’ This directly fuels EU legislative pressure.
EU Regulation 2019/6 (Veterinary Medicines) – Article 118: Banning import of animal products containing antimicrobials used for growth promotion. Application delayed to 2026, raising questions about enforcement timelines – and competitive fairness regarding imports from countries still allowing AGPs.
EU AMR Implementation Decision 2023: New harmonized monitoring requirements for AMR in zoonotic and indicator bacteria from food-producing animals – effective January 1, 2025. All EU Member States now required to collect and report standardized AMR surveillance data.
WOAH 10-Year HPAI Strategy (2024–2033): Promotes surveillance, vaccination programs, and timely reporting as cornerstones of international HPAI management.
BOTTOM LINE ON AMR: The regulatory trajectory is clear and irreversible – sub-therapeutic antibiotic use for growth promotion is being eliminated globally. The timeline varies by region (already banned in EU since 2006; US voluntary approach from 2017; global WHO action plan). Companies that have already invested in transition are ahead; those that have not face increasing compliance risk and market access restrictions.
$793M in proposals received (417 submissions); awards expected by fall 2025; covers prevention, vaccines, therapeutics
DOJ Antitrust Investigation – Egg Producers
Launched April 2025; examining price-fixing allegations amid 247% profit increase by largest producer
Meat & Poultry Special Investigator Act (S.1312)
Proposed creation of Office of Special Investigator for Competition Matters within USDA – pending
Food Security & Farm Protection Act (S.1326)
Would prohibit states from imposing certain standards on preharvest agricultural production sold in interstate commerce – relevant to cage-free mandates
CHAPTER 6: FEED ADDITIVE & NUTRITION STRATEGIES
PRECISION NUTRITION SIGNAL: The industry’s shift to reduced crude protein (CP) diets, precisely supplemented with industrial amino acids (L-Lys, DL-Met, L-Thr, L-Trp, L-Val) remained the dominant reformulation strategy in 2025. Lower CP diets reduce feed cost, lower N excretion (environmental benefit), and reduce substrate for pathogenic bacteria. With amino acid prices remaining favorable, there are few economic arguments for maintaining high CP diets.
6.1 The Post-AGP Transition: Where the Industry Stands
The antibiotic-free (ABF) production movement accelerated further in 2025. With the EU ban on AGPs in place since 2006 and the US moving toward voluntary phase-out, the entire industry is in active transition. The key challenge: AGP removal creates enteric health gaps that must be addressed with alternative tools. Without effective management, removal of AGPs leads to increased necrotic enteritis, Campylobacter colonization, and poorer FCR.
6.2 Heat Stress – A Growing Production Challenge
Climate-related heat stress was a highlighted research and production topic in 2025. Modern high-performance broiler genetics have been selectively bred for rapid growth under thermoneutral conditions. Heat stress impairs feed intake, FCR, immunity, meat quality, and reproduction. Management strategies:
Vitamin C and E supplementation at heat stress periods
Betaine inclusion as an osmolyte; reduces supplemental methionine requirement under heat stress
Feed schedule adjustment (limit feeding during hottest hours; early morning/evening feeding)
Housing design investment: tunnel ventilation, evaporative cooling, adequate air velocity
6.3 In Ovo Technology
In ovo vaccination and nutrition delivery continued to advance in 2025. Key developments include high-throughput systems (3,000 eggs/hour at 99% accuracy) for in ovo vaccination and nutritional interventions. Early-life gut programming through in ovo delivery of probiotics, nutrients, and vaccine antigens is becoming an increasingly important hatchery-level biosecurity and performance tool.
CHAPTER 7: MARKET TRENDS & CONSUMER SHIFTS
7.1 Poultry Gaining Share vs. Other Proteins
Elevated beef prices throughout 2025 – driven by tight US cattle supply (herd at decades-long lows) and high demand – continued to push consumers toward poultry as a cost-effective protein. This dynamic is a structural tailwind for the broiler industry globally.
Market Dynamic
Detail
US broiler net cash farm income 2025
+27% YoY – livestock sector outperforms crop side
Global poultry market value (2025)
$316.77 billion; projected $433.98B by 2034 (CAGR 3.56%)
Global poultry export growth 2025
+1.8% to 16.9 million MT
Supermarkets poultry market share
42.1% of poultry distribution (2024)
Online poultry retail growth rate
CAGR 11.4% (fastest growing channel)
Italy – poultry share of total meat consumed
>44% in 2025
FAO Meat Price Index – poultry
Decreased in 2025 from mid-2024 high (broiler ample supply)
7.2 Cage-Free & Animal Welfare Commitments
The cage-free transition is structurally undersupplied in the US. Corporate commitments made in 2014–2017 implied a need for 220 million cage-free layers by 2025–26. Current production is well below that target. This creates both a market opportunity (premium pricing) and a risk (HPAI vulnerability concerns in cage-free systems). Producers must balance welfare compliance with biosecurity protocols.
7.3 Antibiotic-Free, Organic, and Specialty Products
Consumer and corporate buyer demand for ABF, No Antibiotics Ever (NAE), organic, and pasture-raised products continued to grow in premium markets in 2025. The pasture-raised egg segment reported 30% annual growth rates despite high price points. For integrated producers, this requires dedicated production lines with separate management protocols, supply chain segregation, and robust documentation systems.
7.4 Sustainability Pressure
Feed manufacturers and integrators are under growing pressure from retail and foodservice customers, NGOs, and regulators to demonstrate reduced environmental footprint. Key metrics under scrutiny:
GHG emissions per kg of chicken meat produced (Scope 1, 2, and 3)
Deforestation-free supply chains for soy (EU Deforestation Regulation – EUDR)
Feed conversion ratio improvement as a sustainability lever
EUDR NOTE: The EU Deforestation Regulation requires companies to ensure that soy used in feed does not originate from recently deforested land. Implementation deadlines have been debated, but traceability requirements for soy origin – particularly from Brazil – are operationally significant for EU feed manufacturers and importers.
CHAPTER 8: STRATEGIC LESSONS & ACTION PRIORITIES
8.1 Summary: Top 10 Lessons of 2025
#
Lesson
Key Data Point
1
HPAI is now a permanent structural risk, not a cyclical one. Biosecurity investment must be treated as core capital expenditure.
CDC: H5N1 now enzootic in North American wild birds; US flock 8% below 2022 baseline
2
Egg production is structurally more vulnerable than broiler production – different biosecurity and business continuity protocols are required.
75% of HPAI losses = layers; broilers grew 1.4% in 2025
3
Vaccination for HPAI is the central unresolved debate of the decade – expect DIVA strategies to become standard within 3–5 years as industry and regulators align.
417 vaccine/research proposals submitted to USDA Grand Challenge
4
Trade concentration is a strategic vulnerability. Diversify export markets actively; do not allow 70%+ of any product to go to one trading bloc.
China + Mexico + Canada = 70% of US corn exports; 60% of soy; 45% of poultry
5
Grain prices are favorable NOW – lock in contracts and assess forward pricing opportunities while corn and SBM are at multi-year lows.
Corn -3.9% in 2025; SBM -4.3%; both 3rd consecutive annual decline
6
AMR regulations are accelerating everywhere. Transitioning to ABF production is no longer a ‘maybe’ but a ‘when’ – plan now.
EU: AMR in poultry ‘persistently high’ per EFSA/ECDC March 2025 report
7
EFSA’s 2025 WGS guidance fundamentally changes the cost and timeline of getting microbial feed additives authorized in the EU.
WGS now mandatory for all microbial characterizations; legacy dossiers need revision
8
Amino acids and precision nutrition remain the most cost-effective tool for diet optimization: lower CP, better FCR, lower N excretion, reduced enteric pathogen substrate.
Amino acids = 33.6% of global feed additive market by value
9
Brazil’s HPAI outbreak demonstrated both the vulnerability of global trade and the effectiveness of regionalized response protocols.
Brazil exports fell 12.9% in May but year-end positive; China temporarily banned; UAE stepped up
10
Climate/heat stress is an underappreciated production risk that compounds disease susceptibility and reduces performance in high-performing genetics.
IPCC: global surface temperature +0.9°C since mid-20th century; impacts on poultry FCR, immunity, mortality increasing
8.2 Action Priority Matrix for Management Teams
Priority Area
Immediate Actions (0–6 months)
Medium-Term (6–18 months)
HPAI Biosecurity
Complete USDA-style biosecurity assessments; audit wild bird access; upgrade water and air biosecurity; train all staff
Brazil HPAI market re-entry for China – recovery of the world’s #1 poultry export relationship
US corn/soy 2026 planting intentions (March) – USDA Prospective Plantings report is the key 2026 procurement signal
2025 demonstrated that the feed and animal production industry operates in an environment of simultaneous, compounding risks – biological, geopolitical, regulatory, and climatic. The companies that performed best were those with robust biosecurity infrastructure, agile procurement teams, clear AMR transition roadmaps, and diversified market exposure. There is no single silver bullet. Systematic risk management, not reactive crisis response, is the competitive differentiator going forward.
KEY SOURCES & REFERENCES
This article draws on data and analysis from the following sources:
Organization
Document / Resource Referenced
USDA APHIS / FAS
HPAI flocks data (2025); Livestock & Poultry World Markets (Dec 2025); WASDE reports; Five-Pronged HPAI Strategy
FAO
Food Outlook June 2025; OECD-FAO Agricultural Outlook 2025–2034; FAO Meat Price Index
OECD
OECD-FAO Agricultural Outlook 2025–2034 (July 2025)
WOAH
HPAI Report #68 (Feb 2025); State of World Animal Health 2025; HPAI 10-Year Strategy 2024–2033
EFSA / ECDC
Joint AMR Report (March 2025); 2025 QPS updated list; EFSA 2025 Guidance on Microorganisms (Nov 2025)
PAHO / WHO
Epidemiological Update H5N1 in the Americas (Jan 2025)
Corn & Other Feed Grains Outlook (2025–26 WASDE updates)
Frontiers in Veterinary Science
Phytogenic feed additives – gut health modulation (Aug 2025); Antibiotic alternatives – One Health (Jul 2025)
Learning from AGP mechanisms to advance poultry nutrition
By Ilinca Anghelescu, Dr. Andreas Michels, Predrag Persak
Our understanding of how nutrition influences growth and resilience in poultry has greatly expanded in recent years. It is now clear that animal performance stems to a large extent from a balance between metabolism, immune function, and the gut microbiome. These systems interact continuously, and even small nutritional or environmental changes can shift the animals’ physiological response. This growing knowledge has encouraged the development of nutritional strategies and feed components that work through adaptive, non-antibiotic mechanisms. One recent proposed explanation for these responses has rapidly gained ground: hormetic modeling.
Hormetic modeling describes how small or moderate doses of nutritional components can activate beneficial adaptive responses (improved resilience or metabolic efficiency), while excessive doses become harmful. This idea parallels, largely speaking, Paracelsus’s famous principle: “The dose makes the poison.” In poultry nutrition, such hormetic patterns are well recognized in nutrients like trace elements (selenium, zinc) and specific amino acids (for example, arginine). At optimal levels, these nutrients support antioxidant defense, growth, and immune balance, whereas excessive intake may cause oxidative or metabolic stress
This review examines the hormetic principle and its application to modern poultry/swine feeding concepts, exploring how balanced nutrient design and controlled inclusion of bioactive compounds can strengthen cellular adaptation, improve stress tolerance, and enhance production efficiency.
How do AGPs actually work?
Despite AGP’s widespread historical use, the precise mechanisms by which subtherapeutic doses of antibiotics enhance animal productivity remained poorly understood. Recent advances in systems biology and mitochondrial research propose new answers, much needed to develop future advanced nutritional systems.
The traditional explanations for AGP efficacy have focused primarily on antimicrobial effects:
reducing nutrient competition from microorganisms
decreasing harmful bacterial metabolites
improving gut wall morphology (thinner gut wall ➡ better nutrient absorption)
preventing subclinical infections
However, these mechanisms alone could not fully explain why different classes of antibiotics with diverse mechanisms of action produce similar growth-promoting effects (Gutierrez-Chavez et al., 2025).
Niewold (2007) hypothesized that the primary mechanism of AGPs is non-antibiotic anti-inflammatory activity, reducing the energetic costs of chronic low-grade inflammation. Inflammation diverts nutrients from growth toward immune responses, with cytokine production (particularly IL-1β, IL-6, and TNF-α) suppressing anabolic pathways (Kogut et al., 2018). AGPs appear to selectively inhibit pro-inflammatory cytokine production without completely suppressing immune function.
A paper published in 2024 by Fernandez Miyakawa et al. proposes that antibiotics at subtherapeutic levels act primarily through mitochondrial hormesis and adaptive stress responses, and not simply through antimicrobial activity. In this model, mitochondria act as bioenergetic hubs and signaling centers. Low-dose antibiotics trigger mild mitochondrial stress, which triggers the activation of adaptive protective pathways.This in turn induces mitokine release, leading to systemic adaptive responses improving growth, feed efficiency, and disease tolerance.
Mechanism of action in the hormetic model of AGP efficiency
Hormesis is a biphasic mechanism whereby high doses are toxic, but low doses stimulate adaptive responses and are beneficial. In the case of AGPs, Fernandez Miyakawa et al. propose that low doses stimulate growth, stress resistance, and cellular repair.
Key signaling pathways
As Bottje et al. (2006, 2009) shows, efficient animals often have mitochondrial inner membranes that are less permeable to protons and other ions, allowing for more effective coupling between electron transport and ATP synthesis, which reduces energy loss through proton leak and maximizes the production of ATP per oxygen molecule consumed. Lower membrane permeability is influenced by factors like decreased membrane surface area per protein mass, specific membrane protein content (such as adenine nucleotide translocase), and fatty acid composition in the membrane phospholipids, all contributing to a tighter barrier that prevents unregulated electron or proton flow and supports higher energetic efficiency. Such features make mitochondria in efficient species more capable of maintaining membrane integrity and ATP generation, especially when facing environmental stress, as seen in freeze-tolerant animals whose mitochondria do not undergo damaging permeability transitions under extreme conditions.
Nrf2
Many AGPs interfere with mitochondrial protein synthesis and electron transport chain. At subtherapeutic levels, they cause a mild ROS increase, which triggers the activation of redox-sensitive transcription factor Nrf2. Since Nrf2 regulates over 250 antioxidant, detoxification, and anti-inflammatory genes, the result is improved cell survival, redox balance, and tolerance to stress.
Figure 1 From Zhang et al., 2024
Mitokine production
Mitokines are “signaling molecules that enable communication of local mitochondrial stress to other mitochondria in distant cells and tissues” (Burtscher 2023). Through fibroblast growth factor 21 (FGF21), growth differentiation factor 15 (GDF15), adrenomedullin2 (ADM2) etc, these stress signals are released systemically and coordinate tissue-wide responses, leading to improved growth and resilience.
Inflammation and disease defense
While the negative side of antibiotic growth promoters is well researched and understood (Rahman et al., 2022), science can advance by isolating the positive effects and attempting to offer different pathways to the same benefits. One such lesson can be derived from understanding inflammation pathways and responses.
Chronic low-grade intestinal inflammation is common in modern poultry production, due to diet, microbiota shifts, high metabolic demands etc. This inflammation diverts energy from growth to immune responses.
AGPs reduce the energy costs of this inflammation in three main ways:
Reduces inflammation through adaptive stress response
Raising the threshold to trigger inflammation
Promoting overall resilience, rather than simply killing pathogens
Fernandez Miyakawa et al. suggest, in this emerging model, that disease defense can operate two different actions: resistance to health challenges through reduction of the pathogen load (which is driven by the immune system and is energy costly); and overall resilience by reducing host damage without reducing the pathogen load. AGPs, the authors claim, mainly promote resilience by enhancing mitochondrial stress responses and tissue repair, i.e. more precisely:
Direct mitochondrial stimulation in intestinal epithelial cells
Metabolic optimization supporting growth and feed efficiency
Figure 2 From Fernandez Miyakawa et al., 2024.
In this context, “metabolic optimization” refers to the enhancement of metabolic processes within livestock or poultry to support efficient growth, feed conversion, and physiological resilience, without relying on immune-mediated pathways that are energetically costly. Scientific evidence shows that metabolic optimization involves improving nutrient assimilation, promoting more efficient energy production in tissues (such as mitochondrial ATP synthesis), and minimizing wasteful metabolic byproducts, resulting in reduced feed intake per unit of growth and better utilization of dietary nutrients (Rauw 2025, El-Hack 2025).
Function of feed additives and feed components
Feed additives and feed components in many ways represent the complete other side of the spectrum from antibiotics, but are there some features where antibiotics and feed additives come close in their functions? There is a good case to be made for certain feed additives ultimately working in the animal to achieve similar benefits to the desirable, non-medicinal usage of AGP´s. Especially with the emergent model of AGP mechanism described above, it is worth discussing how certain feed additives can support the same end goal: promoting animal resilience.
Lillejhoj et al (2018), Gutierrez-Chavez et al. (2025) and others outline the end-results such products must achieve:
Growth performance & feed conversion efficiency
Promotion of animal productivity under real-world conditions
Support gut homeostasis
Non-adverse effect on the immune system
Reduction of oxidative stress
Support organism in mitigation of enteric inflammatory consequences
Within the hormetic model, possibly the most important systemic benefit is, in one phrase, promoting resilience. Phytomolecules have long been used, in human and animal medicine, for the same end goal. The mechanisms described below should naturally be seen with caution, as phytomolecule microbiome effects can be subtler and context-dependent. However, the substantiating literature has been increasingly accumulating on these specific topics.
1. Immunometabolic regulation
Phytomolecules demonstrate remarkably similar anti-inflammatory effects to what Niewold (2007) suggested was a primary mechanism of AGPs: non-antibiotic anti-inflammatory activity, reducing the energetic costs of chronic low-grade inflammation. Inflammation diverts nutrients from growth toward immune responses, with cytokine production (particularly IL-1β, IL-6, and TNF-α) suppressing anabolic pathways (Kogut et al., 2018). AGPs appear to selectively inhibit pro-inflammatory cytokine production without completely suppressing immune function. A similar effect can be observed with various types of phytomolecules, which significantly reduced pro-inflammatory and/or increased anti-inflammatory cytokine expression in animals challenged with several pathogens. The anti-inflammatory mechanism appears to involve inhibition of NF-κB activation and modulation of MAPK signaling pathways (Kim et al., 2010; Long et al., 2021).
2. Mitochondrial hormesis and energy metabolism
Fernández Miyakawa et al. (2024, see above) proposed that AGPs exert growth-promoting effects through mitochondrial hormesis – subtherapeutic antibiotic doses induce mild mitochondrial stress, triggering adaptive responses that enhance mitochondrial function, energy metabolism, and cellular resilience. This mechanism, while requiring further validation, explains why different antibiotics with diverse targets produce similar growth outcomes.
The mitochondrial stress response involves activation of the IL-6 receptor family signaling cascade, which regulates metabolism, growth, regeneration, and homeostasis in liver and other tissues (Perry et al., 2024). Subtherapeutic antibiotic exposure activates proteins involved in growth and proliferation through IL-6R gp130 subunit signaling, including JAK, STAT, mTOR, and MAPK pathways.
Phytomolecules demonstrate similar mitochondrial effects. Perry et al. (2024) showed that increased activity of AMPK, mTOR, PGC-1α, PTEN, HIF, and S6K can also be available via phytomolecule activity, suggesting enhanced anabolic metabolism.
Capsicum oleoresin supplementation in broilers increased jejunal lipase and trypsin activity, enhanced ileal amylase activity, improved jejunal morphology, and modulated immune organ development, indicating enhanced digestive efficiency and nutrient utilization (Li et al., 2022).
Compounds such as vanillin, thymol, eugenol have been shown to improve glucose and lipid metabolism through TRPV1 activation and mitochondrial function enhancement (Gupta et al., 2022; Zhang et al., 2017).
3. Gut microbiota modulation
AGPs selectively reduce specific microbial populations, particularly Lactobacillus species that produce bile salt hydrolase (BSH). Since BSH reduces fat digestibility and thus weight gain, AGP-mediated reduction of BSH-producing bacteria enhances energy extraction and growth (Lin, 2014; Bourgin et al., 2021).
Recent research by Zhan et al. (2025) using single-molecule real-time 16S rRNA sequencing demonstrated that therapeutic antibiotic doses (lincomycin, gentamicin, florfenicol, benzylpenicillin, ceftiofur, enrofloxacin) significantly altered chicken gut microbiota composition, with Pseudomonadota and Bacillota becoming dominant phyla after exposure. Different antibiotics produced distinct temporal effects on microbial diversity and community structure.
Phytomolecules exert targeted antimicrobial effects while promoting beneficial bacteria. Dietary supplementation with 800 mg/kg Capsicum extract in Japanese quails reduced cecal counts of pathogenic bacteria (Salmonella spp., E. coli, coliforms) while modulating Lactobacilli populations (Reda et al., 2020).
In pigs, 80 mg/kg natural capsicum extract increased cecal propionic acid and total volatile fatty acid concentrations, with increased butyric acid in the colon – indicating enhanced fermentation by beneficial bacteria (Long et al., 2021).
Capsicum and Curcuma oleoresins altered intestinal microbiota composition in commercial broilers challenged with necrotic enteritis, reducing disease severity through microbiome modulation (Kim et al., 2015).
Capsaicin demonstrates selective antimicrobial activity, inhibiting pathogenic Gram-negative bacteria while favoring development of certain Gram-positive bacteria. The antibacterial mechanism involves induction of osmotic stress and membrane structure damage (Adaszek et al., 2019; Rosca et al., 2020).
4. Intestinal barrier function and gut health
AGPs have been associated with improved intestinal morphology, including increased villus height and reduced crypt depth, which enhance absorptive capacity (Gaskins et al., 2002).
Phytomolecules produce similar or superior effects. Capsicum extract (80 mg/kg) in pigs increased ileal villus height and upregulated MUC-2 gene expression, indicating enhanced gut barrier function and integrity. The improved barrier function correlated with reduced diarrhea incidence (Liu et al., 2013; Long et al., 2021).
Allium hookeri extract increased expression of tight junction proteins (claudins, occludins, ZO-1) in LPS-challenged broiler chickens, demonstrating direct enhancement of barrier integrity (Lee et al., 2017).
5. Oxidative stress mitigation
Oxidative stress impairs growth by damaging cellular components and triggering inflammatory responses. AGPs reduce oxidative stress indirectly through anti-inflammatory effects and microbiota modulation (Bortoluzzi et al., 2021).
Phytomolecules possess direct antioxidant properties. Capsicum extract (50 mg/kg) in heat-stressed quails reduced serum and ovarian malondialdehyde (MDA) while increasing superoxide dismutase (SOD) and catalase (CAT) activities. Ovarian transcription factors showed decreased NF-κB and increased Nrf2 and HO-1 expression (Sahin et al., 2016).
A mixture of herbal extracts including pepper reduced thiobarbituric acid reactive substances and MDA in broiler liver and muscle, while increasing glutathione peroxidase (GSH-Px) activity and improving antioxidant enzyme expression (Saleh et al., 2018).
Capsicum extract (80 mg/kg) in pigs increased total antioxidant capacity, SOD, and CAT while reducing MDA levels, demonstrating robust antioxidant effects (Long et al., 2021).
Standardization and controlled release: Critical success factors
A major criticism of phytomolecules has been inconsistent efficacy across studies. However, this variability largely reflects differences in:
Active compound concentrations
Bioavailability and stability
Dosing precision
Product quality and standardization
Microencapsulation is one of the technologies that address the standardization and bioavailability challenges. It protects volatile compounds from degradation during feed processing and storage, with encapsulated essential oils showing significantly higher retention compared to unprotected forms (Stevanović et al., 2018). By creating a protective barrier around active ingredients, microencapsulation enables controlled release in specific regions of the gastrointestinal tract, improving absorption efficiency and reducing dose variability (Bringas-Lantigua et al., 2011). The technology also masks unpalatable flavors that can reduce feed intake while standardizing active ingredient concentrations through precise manufacturing processes (Gharsallaoui et al., 2007). Studies demonstrate that spray-dried microencapsulated essential oils achieve encapsulation efficiencies exceeding 93% with minimal loss during storage (Hu et al., 2020), and can be engineered for enzyme-mediated release to ensure bioactive delivery at optimal intestinal sites (Elolimy et al., 2025).
Mechanistic synthesis: An integrated model
The evidence indicates that both AGPs and phytomolecules operate through an integrated network of effects:
Primary Level: Selective antimicrobial effects modify gut microbiota composition
This integrative model explains why multiple antibiotics with different mechanisms produce similar growth outcomes: they converge on common pathways regulating immunometabolism and mitochondrial function (Fernández Miyakawa et al., 2024).
Phytomolecules operate through the same mechanistic framework but with potential advantages:
Safety and antimicrobial resistance considerations
Antibiotic exposure significantly disrupts gut microbiota diversity and stability, with effects persisting beyond withdrawal periods. The study by Zhan et al. (2025) demonstrated that different antibiotics produce varying degrees of microbiota disruption, with florfenicol and gentamicin showing the strongest and most persistent effects.
In contrast, phytomolecules generally do not generate resistance through the same mechanisms as antibiotics. Some phytochemicals may actually enhance antibiotic efficacy and resensitize resistant bacteria through structural modifications of bacterial membranes (Khameneh et al., 2021; Suganya et al., 2022).
However, one study reported increased correlation between antibiotic resistance genes (ARGs) and mobile genetic elements in pig feces after mushroom powder supplementation, suggesting that certain phytogenic compounds may increase ARG mobility (Muurinen et al., 2021). This emphasizes the need for continued surveillance of phytomolecule effects on resistance gene dynamics.
Capsaicinoids and capsinoids have well-established safety profiles. Capsiate, a non-pungent analogue of capsaicin, exhibits substantially lower toxicity while maintaining similar metabolic and growth-promoting effects (Gupta et al., 2022). No adverse effects on animal health or product quality have been reported at recommended dosages in reviewed studies.
Future directions and research needs
Despite substantial progress, several areas require further investigation:
Mechanistic refinement: Detailed characterization of signaling pathways, particularly the IL-6R/gp130 cascade and mitochondrial stress responses
Precision formulation: Development of combinations optimized for specific production stages, environmental conditions, and disease pressures
Bioavailability optimization: Advanced delivery systems ensuring consistent active compound release and absorption
Microbiome-host interaction mapping: High-resolution characterization of microbial community shifts and their functional consequences
Economic validation: Large-scale production trials assessing cost-effectiveness compared to AGPs and disease management costs
Conclusions
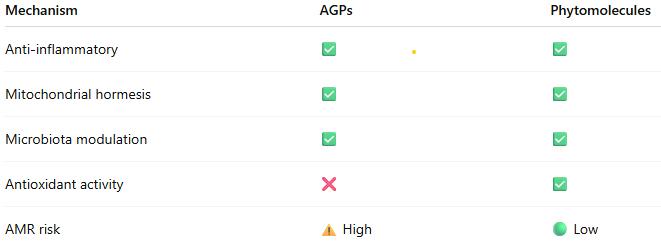
The scientific evidence demonstrates that standardized phytomolecules operate through well-characterized biological mechanisms that substantially replicate those of AGPs:
Anti-inflammatory effects reducing energetic costs of immune activation
Mitochondrial hormesis enhancing energy metabolism and cellular resilience
Selective microbiota modulation supporting beneficial bacteria while controlling pathogens
Intestinal barrier enhancement improving nutrient absorption and reducing translocation
Antioxidant activity mitigating oxidative stress and supporting immune function
When properly standardized and formulated for controlled release, phytomolecules deliver growth promotion, feed efficiency improvements, and disease resistance comparable to AGPs, while potentially offering advantages in AMR risk profile, stress resilience, and consumer acceptance.
The mechanistic convergence between AGPs and phytomolecules, coupled with demonstrated efficacy in controlled trials, provides producers with confidence that science-based phytomolecular interventions represent legitimate alternatives to AGPs. Success depends on product standardization, appropriate dosing, and understanding that phytomolecules work through fundamental biological pathways rather than undefined or mystical mechanisms.
As the livestock industry continues to navigate the post-AGP era, standardized phytomolecules offer a scientifically sound, mechanistically validated approach to maintaining animal performance, health, and welfare while addressing antimicrobial resistance concerns.
References
Adaszek, Ł., et al. “Properties of Capsaicin and Its Utility in Veterinary and Human Medicine.” Research in Veterinary Science, vol. 123, 2019, pp. 14 – 19.
Bottje, W., et al. “Mitochondrial proton leak kinetics and relationship with feed efficiency within a single genetic line of male broilers”. Poultry Science, Volume 88, Issue 8, 1 August 2009, p. 1683-1693.
Bortoluzzi, C., et al. “A Protected Complex of Biofactors and Antioxidants Improved Growth Performance and Modulated the Immunometabolic Phenotype of Broiler Chickens Undergoing Early Life Stress.” Poultry Science, vol. 100, 2021, p. 101176.
Bourgin, M., et al. “Bile Salt Hydrolases: At the Crossroads of Microbiota and Human Health.” Microorganisms, vol. 9, no. 1122, 2021.
Bravo, D., et al. “A Mixture of Carvacrol, Cinnamaldehyde, and Capsicum Oleoresin Improves Energy Utilization and Growth Performance of Broiler Chickens Fed Maize-Based Diet.” Journal of Animal Science, vol. 92, 2014, pp. 1531 – 1536.
Bringas-Lantigua, M., et al. “Influence of Spray-Dryer Air Temperatures on Encapsulated Mandarin Oil.” Drying Technology, vol. 29, 2011, pp. 520–526.
Burtscher, J., et al. “Mitochondrial Stress and Mitokines in Aging.” Aging Cell, vol. 22, no. 2, 2023, e13770.
El-Hack, M. et al. “Integrating metabolomics for precision nutrition in poultry: optimizing growth, feed efficiency, and health”. Frontiers in Veterinary Science, Sec. Animal Nutrition and Metabolism, Volume 12 – 2025. https://doi.org/10.3389/fvets.2025.1594749
Elolimy, Ahmed A., et al. “Effects of Microencapsulated Essential Oils and Seaweed Meal on Growth Performance, Digestive Enzymes, Intestinal Morphology, Liver Functions, and Plasma Biomarkers in Broiler Chickens.” Journal of Animal Science, vol. 103, 2025, p. skaf092, https://doi.org/10.1093/jas/skaf092.
Fernández Miyakawa, Mariano E., et al. “How Did Antibiotic Growth Promoters Increase Growth and Feed Efficiency in Poultry?” Poultry Science, vol. 103, no. 2, 2024, article 103136. https://doi.org/10.1016/j.psj.2023.103136
Gaskins, H. Rex, C. T. Collier, and D. B. Anderson. “Antibiotics as Growth Promotants: Mode of Action.” Animal Biotechnology, vol. 13, no. 1, 2002, pp. 29 – 42.
Gharsallaoui, A., et al. “Applications of Spray-Drying in Microencapsulation of Food Ingredients: An Overview.” Food Research International, vol. 40, no. 9, 2007, pp. 1107-21.
Gutiérrez-Chávez, Vanesa, et al. “Capsaicinoids and Capsinoids of Chilli Pepper as Feed Additives in Livestock Production: Current and Future Trends.” Animal Nutrition, vol. 22, 2025, pp. 483 – 501. https://doi.org/10.1016/j.aninu.2025.03.014.
Gupta, A., et al. “Capsaicin and Capsinoids: Recent Updates on Their Health Benefits and Mechanisms of Action.” Phytotherapy Research, vol. 36, no. 5, 2022, pp. 1898 – 1912.
Hu, Q., Li, X., Chen, F., Wan, R., Yu, C.-W., Li, J., McClements, D. J., & Deng, Z. (2020). “Microencapsulation of an essential oil (cinnamon oil) by spray drying: Effects of wall materials and storage conditions on microcapsule properties“. Journal of Food Processing and Preservation, 44(11). https://doi.org/10.1111/jfpp.14805
Khameneh, B., et al. “Mechanisms of Antibiotic Resistance Resensitization by Phytochemicals: Review.” Phytomedicine, vol. 85, 2021, p. 153529.
Kim, D. K., et al. “Effects of Capsicum and Curcuma on Necrotic Enteritis in Broilers.” Poultry Science, vol. 94, 2015, pp. 2314 – 2321.
Kim, J. S., et al. “Anti-inflammatory Effects of Plant-Derived Molecules via NF-κB and MAPK Pathways.” International Immunopharmacology, vol. 10, no. 3, 2010, pp. 306 – 314.
Lee, S. H., et al. “Allium Hookeri Extract Enhances Tight Junction Proteins in Broilers.” Journal of Animal Physiology and Animal Nutrition, vol. 101, no. 1, 2017, pp. e48 – e56.
Li, X., et al. “Capsicum Oleoresin Supplementation Improves Digestive Enzyme Activity and Gut Morphology in Broilers.” Poultry Science, vol. 101, no. 7, 2022, p. 101844.
Lin, J. “Effect of Antibiotics on the Intestinal Microbiota and Their Role in Animal Growth.” Animal Biotechnology, vol. 25, no. 3, 2014, pp. 149 – 157.
Lillehoj, H., et al. “Phytochemicals as Antibiotic Alternatives to Promote Growth and Enhance Host Health.” Veterinary Research, vol. 49, no. 76, 2018.
Liu, Y., et al. “Dietary Capsicum Extract Enhances Intestinal Barrier Function and Growth in Pigs.” Journal of Animal Science, vol. 91, 2013, pp. 518 – 525.
Long, L., et al. “Phytogenic Feed Additives Modulate Intestinal Immunity and Antioxidant Status in Pigs and Poultry.” Frontiers in Veterinary Science, vol. 8, 2021, p. 620998.
Muurinen, J., et al. “Mushroom Powder Supplementation Increases Antibiotic Resistance Gene Mobility in Pig Feces.” Frontiers in Microbiology, vol. 12, 2021, p. 676678.
Niewold, T. A. “The Non-antibiotic Anti-inflammatory Effect of Antimicrobial Growth Promoters, the Real Mode of Action? A Hypothesis.” Poultry Science, vol. 86, 2007, pp. 605 – 609.
Perry, F., C. N. Johnson, L. Lahaye, E. Santin, D. R. Korver, M. H. Kogut, and R. J. Arsenault. “Protected Biofactors and Antioxidants Reduce the Negative Consequences of Virus and Cold Challenge by Modulating Immunometabolism via Changes in the Interleukin-6 Receptor Signaling Cascade in the Liver.” Poultry Science, vol. 103, no. 9, 2024, article 104044. https://doi.org/10.1016/j.psj.2024.104044
Rahman, Md, et al. “Insights in the Development and Uses of Alternatives to Antibiotic Growth Promoters in Poultry and Swine Production.” Antibiotics, vol. 11, no. 6, 2022, p. 766, https://doi.org/10.3390/antibiotics11060766.
Reda, F. M., et al. “Capsicum Extract Supplementation Modulates Gut Microbiota and Performance in Japanese Quails.” Animal Feed Science and Technology, vol. 265, 2020, p. 114507.
Rosca, I., et al. “Capsaicin Induces Osmotic Stress in Gram-negative Pathogens.” Veterinary Sciences, vol. 7, no. 4, 2020, p. 172.
Sahin, K., et al. “Dietary Capsicum Extract Reduces Oxidative Stress in Heat-stressed Japanese Quails.” Poultry Science, vol. 95, no. 2, 2016, pp. 231 – 240.
Saleh, A. A., et al. “Herbal Extract Mixtures Improve Antioxidant Status and Performance in Broilers.” Poultry Science, vol. 97, no. 11, 2018, pp. 3927 – 3936.
Stevanović, Z. D., et al. „Essential oils as feed additives—Future perspectives”. Molecules, 23(7), 2018, pp1717.
Suganya, R., et al. “Phytochemicals in Combination with Antibiotics: Antimicrobial Resistance Breakers.” Antibiotics, vol. 11, 2022, p. 123.
Zhang, Benyuan et al. “Mitochondrial Stress and Mitokines: Therapeutic Perspectives for the Treatment of Metabolic Diseases.” Diabetes & Metabolism Journal vol. 48,1, 2024, pp. 1-18.
Zhan, Ru, et al. “Effects of Antibiotics on Chicken Gut Microbiota: Community Alterations and Pathogen Identification.” Frontiers in Microbiology, vol. 16, 2025, article 1562510. https://doi.org/10.3389/fmicb.2025.1562510
Zhang, Y., et al. “Effects of Vanillin, Thymol, and Eugenol on Glucose and Lipid Metabolism via TRPV1 Activation.” Journal of Agricultural and Food Chemistry, vol. 65, no. 13, 2017, pp. 2719 – 2727.
Dietary interventions for resilient poultry gut health in the AMR era
by Ajay Bhoyar, Global Technical Manager, EW Nutrition
Gut health is critical for profitable poultry production, as the gastrointestinal tract (GIT) plays a dual role in nutrient digestion and absorption while serving as a crucial defense against pathogens. A healthy gut enables efficient feed conversion, robust immune function, and resilience against diseases, reducing reliance on preventive and therapeutic antibiotics. Optimum gut health has become increasingly important in poultry production to combat antimicrobial resistance (AMR), a pressing global challenge threatening animal agriculture and public health.
AMR arises when bacteria develop antibiotic resistance, often due to overuse or misuse in human and animal settings. Predictive models suggest that by 2050, AMR could result in 10 million annual deaths and a 2.0%–3.5% reduction in global gross domestic production, amounting to economic losses between 60 and 100 trillion USD. In poultry, AMR compromises flock health, leading to higher mortality, reduced growth performance, and elevated treatment costs, directly impacting profitability. Additionally, resistant pathogens increase the risk of zoonotic disease transfer, posing serious food safety concerns.
Stricter regulations and rising consumer demand for antibiotic-free poultry products drive a shift toward sustainable, antibiotic-free production systems. However, A lack of understanding about strategies to replace AMU and their effectiveness under field conditions hampers change in farming practices (Afonso et al., 2024). Addressing AMR requires a holistic approach, encompassing enhanced biosecurity, innovative health-promoting strategies, and sustainable management practices. This paper explores practical dietary interventions to support poultry gut health while reducing dependency on antimicrobials, offering solutions for the long-term sustainability of poultry production.
Gut Mediated Immunity in Chickens
The gut is a critical component of the immune system, as it is the first line of defense against pathogens that enter the body through the digestive system. Chickens have a specialized immune system in the gut, known as gut-associated lymphoid tissue (GALT), which helps to identify and respond to potential pathogens. The GALT includes Peyer’s patches, clusters of immune cells in the gut wall, and the gut-associated lymphocytes (GALs) found throughout the gut. These immune cells recognize and respond to pathogens that enter the gut.
The gut-mediated immune response in chickens involves several mechanisms, including activating immune cells, producing antibodies, and releasing inflammatory mediators. GALT and GALs play a crucial role in this response by identifying and responding to pathogens and activating other immune cells to help fight off the infection.
The gut microbiome is a diverse community of microorganisms that live in the gut. These microorganisms can significantly impact the immune response. Certain beneficial bacteria, for example, can help stimulate the immune response and protect the gut from pathogens.
Overall, the gut microbiome, GALT, and GALs work together to create an environment hostile to pathogens while supporting the growth and health of beneficial microorganisms.
Key Factors Affecting Poultry Gut Health
The key factors affecting broiler gut health can be summarized as follows:
Early gut development: Gut-associated immunity responds to early feeding and dietary nutrients and is critical for protecting against exogenous organisms during the first week of life post-hatch.
Feed and Water Quality: The form, type, and quality of feed provided to broilers can significantly impact their gut health. Consistently available cool and hygienic drinking water is crucial for optimum production performance.
Stressors: Stressful conditions, such as high environmental temperatures or poor ventilation, can lead to an imbalance in the gut microbiome and an increased risk of disease.
Infections and medications: Exposure to pathogens or other harmful bacteria can disrupt the gut microbiome and lead to gut health issues. A robust immune system is important for maintaining gut health, as it helps to prevent the overgrowth of harmful bacteria and promote the growth of beneficial bacteria.
Biosecurity: Keeping the poultry environment clean and free of pathogens is crucial for maintaining gut health, as bacteria and other pathogens can quickly spread and disrupt the gut microbiome.
Management practices: Best practices, including proper litter management, can help maintain gut health and prevent gut-related issues.
Dietary Interventions for Optimum Gut Health
Gut health means the absence of gastrointestinal disease, the effective digestion and absorption of feed, and a normal and well-established microbiota (Bischoff, 2011). Various dietary measures can be taken to support the healthy functioning of the GIT and host defense. Water and feed safety and quality, feeding management, the form the feed is provided in (e.g., pellets), the composition of the diet, and the use of various feed additives are all tools that can be used to support health (Smits et al., 2021).
Various gut health-supporting feed additives, including organic acids, probiotics, prebiotics, phytochemicals/essential oils, etc., in combination or alone, have been explored as an alternative to antimicrobials in animal production. There were differences in the impacts of the strategies between and within species; this highlights the absence of a ‘one-size-fits-all’ solution. Nevertheless, some options seem more promising than others, as their impacts were consistently equivalent or positive when compared with animal performance using antimicrobials (Afonso et al., 2024). Including insoluble fibers, toxin binders, exogenous enzymes, and antioxidants in the feed formulations also play a crucial role in gut health optimization, which goes beyond their primary functions to combat AMR challenges.
Fig. 1: Multifactorial approach to gut health management in reduced antimicrobial use
Organic Acids
The digestive process extensively includes microbial fermentation, and as a result, organic acids are commonly produced by beneficial bacteria in the crop, intestines, and ceca (Huyghebaert et al., 2010). Organic acids’ inclusion in the poultry diet can improve growth performance due to improved gut health, increased endogenous digestive enzyme secretion and activity, and nutrient digestibility. Butyrate is highly bioactive in GIT. It increases the proliferation of enterocytes, promotes mucus secretion, and may have anti-inflammatory properties (Bedford and Gong, 2018; Canani et al., 2011; Hamer et al., 2008). These effects suggest that it supports mucosal barrier function. Butyrate is becoming a commonly used ingredient in diets to promote GIT health.
Including organic acids in the feed can decontaminate feed and potentially reduce enteric pathogens in poultry. Alternately, the formaldehyde treatment of feed is highly effective at a relatively low cost (Jones, 2011; Wales, Allen, and Davies, 2010).
Organic acids like formic and citric acid are also used in poultry drinking water to lower the microbial count by lowering the water’s pH and preventing/removing biofilms in the water lines. By ensuring feed and water hygiene, producers can minimize pathogen exposure, enhance bird health, and significantly reduce their reliance on antibiotics.
Probiotics, Postbiotics, Prebiotics and Synbiotics
Probiotics and prebiotics have drawn considerable attention to optimizing gut health in animal feeds. Probiotic supplementation could have the following effects: (1) modification of the intestinal microbiota, (2) stimulation of the immune system, (3) reduction in inflammatory reactions, (4) prevention of pathogen colonization, (5) enhancement of growth performance, (6) alteration of the ileal digestibility and total tract apparent digestibility coefficient, and (7) decrease in ammonia and urea excretion (Jha et al., 2020). Certain Lactobacilli or Enterococci species may be used with newly hatched or newborn animals; single or multi-strain starter cultures can be used to steer the initial microbiota in a desired direction (Liao and Nyachoti, 2017). Apart from using probiotics in feed and drinking water, probiotic preparations can be sprayed on day-old chicks in the hatchery or immediately after placement in the brooding house. This way, the probiotic strains/beneficial bacteria gain access to the gut at the earliest possible time (early seeding). Postbiotics are bioactive compounds produced by probiotics during fermentation, such as short-chain fatty acids, peptides, and bacterial cell wall components. Unlike live probiotics, postbiotics are stable, safer, and provide consistent health benefits.
Prebiotics like mannan-oligosaccharides (MOS), inulin, and its hydrolysate (fructo-oligosaccharides: FOS) play an important role in modulating intestinal microflora and potential immune response. Prebiotics reduce pathogen colonization in poultry and promote selective stimulation of beneficial bacterial species. Synbiotics are a combination of probiotics and prebiotics. This synergistic approach offers dual benefits by promoting the growth of beneficial bacteria and directly combating pathogens.
Dietary Fibers (DF)
The water-insoluble fibers are regarded as functional nutrients because of their ability to escape digestion and modulate nutrient digestion. A moderate level of insoluble fiber in poultry diets may increase chyme retention time in the upper part of the GIT, stimulating gizzard development and endogenous enzyme production, improving the digestibility of starch, lipids, and other dietary components (Mateos et al., 2012). The insoluble DF, when used in amounts between 3–5% in the diet, could have beneficial effects on intestinal development and nutrient digestibility.
Dietary fibers influence the development of the gizzard in poultry birds. A well-developed gizzard is a must for good gut health. Jiménez-Moreno & Mateos (2012) noted that coarse fiber particles are selectively retained in the gizzard, ensuring a complete grinding and a well-regulated feed flow. Secretion of digestive juices regulates GIT motility and feed intake. Including insoluble fibers in adequate amounts improves the gizzard function and stimulates HCl production in the proventriculus, thus helping control gut pathogens.
Toxin Risk Management
Mycotoxins may have a detrimental impact on the mucosal barrier function in animals (Akbari et al., 2017; Antonissen et al., 2015; Basso, Gomes and Bracarense, 2013; Pierron, Alassane-Kpembi and Oswald, 2016). Mycotoxins like Aflatoxin B1, Ochratoxin A, and deoxynivalenol (DON) not only suppress immune responses but also induce inflammation and even increase susceptibility to pathogens (Yuhang et al., 2023). To avoid intestinal health problems, poultry producers need to emphasize avoiding levels of mycotoxins in feedstuffs and rancid fats that exceed recommended limits (Murugesan et al., 2015; Grenier and Applegate, 2013).
Fusarium mycotoxin
Bacterial lipopolysaccharides (LPS), also known as endotoxins, are the main components of the outer membrane of all Gram-negative bacteria and are essential for their survival. In stress situations, the intestinal barrier function is impaired, allowing the passage of endotoxins into the bloodstream. When the immune system detects LPS, inflammation sets in and results in adverse changes in gut epithelial structure and functionality. Dietary Intervention to bind these endotoxins in the GIT can help mitigate the negative impact of LPS on animals. Given this, toxin risk management with an appropriate binding agent able to control both mycotoxins and endotoxins appears to be a promising strategy for reducing their adverse effects. Further, adding antioxidants and mycotoxin binders to feed can reduce the effects of mycotoxins and peroxides and is more necessary in ABF programs (Yegani and Korver, 2008).
Essential oils/Phytomolecules
Essential oils (EOs) are important aromatic components of herbs and spices and are used as natural alternatives for replacing antibiotic growth promoters (AGPs) in poultry feed. The beneficial effects of EOs include appetite stimulation, improvement of enzyme secretion related to food digestion, and immune response activation (Krishan and Narang, 2014)
Essential oils (EOs), raw extracts from plants (flowers, leaves, roots, fruit, etc.), are an unpurified mix of different phytomolecules. The raw extract from Oregano is a mix of various phytomolecules (Terpenoids) like carvacrol, thymol and p-cymene. Whereas the phytomolecules are active ingredients of essential oils or other plant materials. Phytomolecule is clearly defined as one active compound.
These botanicals have received increased attention as possible growth performance enhancers for animals in the last decade via their beneficial influence on lipid metabolism, and antimicrobial and antioxidant properties (Botsoglou et al., 2002), ability to stimulate digestion (Hernandez et al., 2004), immune enhancing activity, and anti-inflammatory potential (Acamovic and Brooker, 2005). Many studies have been reported on supplementing poultry diets with some essential oils that enhanced weight gain, improved carcass quality, and reduced mortality rates (Williams and Losa, 2001). The use of some specific EO blends has been shown to have efficacy towards reducing the colonization and proliferation of Clostridium perfringens and controlling coccidia infection and, consequently, may help to reduce necrotic enteritis (Guo et al., 2004; Mitsch et al., 2004; Oviedo-Rondón et al., 2005, 2006a, 2010).
Salmonella
Antimicrobial properties of phytomolecules hinder the growth of potential pathogens. Thymol, eugenol, and carvacrol are structurally similar and have been proven to exert synergistic or additive antimicrobial effects when combined at lower concentrations (Bassolé and Juliani, 2012). In in-vivo studies, essential oils used either individually or in combination have shown clear growth inhibition of Clostridium perfringens and E. coli in the hindgut and ameliorated intestinal lesions and weight loss than the challenged control birds (Jamroz et al., 2006; Jerzsele et al., 2012; Mitsch et al., 2004). One well-known mechanism of antibacterial activity is linked to their hydrophobicity, which disrupts the permeability of cell membranes and cell homeostasis with the consequence of loss of cellular components, influx of other substances, or even cell death (Brenes and Roura, 2010; Solórzano-Santos and Miranda-Novales, 2012; Windisch et al., 2008; O’Bryan et al., 2015).
Apart from use in feed, the liquid phytomolecules preparations for drinking water use can prove to be beneficial in preventing and controlling losses during challenging periods of bird’s life (feed change, handling, environmental stress, etc.). Liquid preparations can potentially reduce morbidity and mortality in poultry houses and thus the use of therapeutic antibiotics. Barrios et al. (2021) suggested that commercial blends of phytomolecule preparations may ameliorate the impact of Necrotic Enteritis on broilers. Further, they hypothesized that the effects of liquid preparation via drinking water were particularly important in improving overall mortality.
In modern, intensive poultry production, the imminent threat of resistant Eimeria looms large, posing a significant challenge to the sustainability of broiler operations. Eimeria spp., capable of developing resistance to traditional anticoccidial drugs, has become a pressing global issue for poultry operators. The resistance of Eimeria to traditional drugs, coupled with concerns over drug residue, has necessitated a shift towards natural, safe, and effective alternatives. It was found that if a drug to which the parasite has developed resistance is withdrawn from use for some time or combined with another effective drug, the sensitivity to that drug may return (Chapman, 1997).
Several phytogenic compounds, including saponins, tannins, essential oils, flavonoids, alkaloids, and lectins, have been the subject of rigorous study for their anticoccidial properties. Among these, saponins and tannins in specific plants have emerged as powerful tools in the fight against these resilient protozoa. Botanicals and natural identical compounds are well renowned for their antimicrobial and antiparasitic activity so that they can represent a valuable tool against Eimeria (Cobaxin-Cardenas, 2016). The mechanisms of action of these molecules include degradation of the cell wall, cytoplasm damage, ion loss with reduction of proton motive force, and induction of oxidative stress, which leads to inhibition of invasion and impairment of Eimeria spp. development (Abbas et al., 2012; Nazzaro et al., 2013). Natural anticoccidial products may provide a novel approach to controlling coccidiosis while meeting the urgent need for control due to the increasing emergence of drug-resistant parasite strains in commercial poultry production (Allen and Fetterer, 2002).
Role of Feed Enzymes Beyond Feed Cost Reduction
Feed enzymes have traditionally been associated with improving feed efficiency and reducing feed costs by enhancing nutrient digestibility. However, their role can extend well beyond economic benefits, profoundly impacting gut health and supporting reduced antimicrobial use in poultry production. Exogenous enzymes reduce microbial proliferation by reducing the undigestible components of feed, the viscosity of digesta, and the irritation to the gut mucosa that causes inflammation. Enzymes also generate metabolites that promote microbial diversity which help to maintain gut ecosystems that are more stable and more likely to inhibit pathogen proliferation (Bedford, 1995; Kiarie et al., 2013).
High dietary levels of non-starch polysaccharides (NSPs) can increase the viscosity of digesta. This leads to an increase in the retention time of the digesta, slows down the nutrient digestion and absorption rate, and may lead to an undesired increase in bacterial activity in the small intestine (Langhout et al., 2000; Smits et al., 1997). Further the mucosal barrier function may also be adversely affected. To solve this problem, exogenous enzymes, such as arabinoxylanase and/ or ß-glucanase, are included in feed to degrade viscous fibre structures (Bedford, 2000). The use of xylanase and ß-glucanase may also cause oligosaccharides and sugars to be released, of which certain, for example, arabinoxylan oligosaccharides, may have prebiotic properties (De Maesschalck et al., 2015; Niewold et al., 2012).
New generation xylanases coming from family GH-10 are known to effectively breakdown both soluble and insoluble arabinoxylans into a good mixture of smaller fractions of arabino-xylo-oligosaccharides (AXOS) and xylo-oligosaccharides (XOS), which exert a prebiotic effect in the GIT. Awati et.al. (2023) observed that a novel GH10 xylanase contributed to positive microbial shift and mitigated the anti-nutritional gut-damaging effects of higher fiber content in the feed. With a substantial understanding of the mode of action and technological development in enzyme technology, nutritionists can reliably consider new-generation xylanases for gut health optimization in their antibiotic reduction strategy.
Conclusions
The challenge of mitigating antimicrobial resistance (AMR) in poultry production necessitates a multidimensional approach, with gut health at its core. Dietary interventions, such as organic acids, probiotics, prebiotics, phytomolecules, toxin binders, and feed enzymes, promote gut resilience, enhance immune responses, and reduce reliance on antimicrobials. These strategies not only support the health and productivity of poultry but also address critical global issues of AMR and food safety.
While no single solution fits all circumstances, integrating these dietary tools with robust biosecurity measures, sound management practices, and continued research on species-specific and field-applicable strategies can pave the way for sustainable, antibiotic-free poultry production. The transition to such systems aligns with regulatory requirements and consumer expectations while contributing to global efforts against AMR.
Ultimately, embracing holistic and innovative dietary strategies ensures a resilient gastrointestinal environment, safeguarding poultry health and productivity while protecting public health and environmental sustainability for future generations.
References: The references can be made available upon request to the author.
Rearing pigs without antibiotics
Conference Report
Holistic management is essential for successfully rearing pigs, particularly in systems that aim to minimize antibiotics. The method emphasizes the interconnectedness of various factors contributing to sustainable pig health and productivity. Some of the key components of this holistic management were discussed by Dr. Seksom.
Sow lifetime productivity
Suggested targets for sow lifetime productivity are
>70 marketed fattening pigs
At least 6 parities with at least 10.5 pigs marketed per parity
25 fattening pigs/sow/year (2.4 parities/year x 10.5 fattening pigs)
To achieve these targets, we need 29.2 born alive piglets/sow/year (or 12.2 born alive piglets/parity), and it is essential to control losses during each production period: <10% pre-weaning, <3% during nursery, and <2% in fattening.
Since the occurrence of African swine fever (ASF), with improved genetics, we can now produce pigs with 120 kg+ bodyweight at slaughter without carcass problems and reach about 3 tons of bodyweight/sow/year, compared to around 2 tons before.
Modern pig genetics and subsequent problems
Despite the advancements in modern pig genetics leading to improved production and bigger litters, several ensuing problems have emerged:
Less average body weight of piglets at birth
Large number of piglets born with less than 1.0 kg (target <5%)
High pre-weaning mortality
High post-weaning mortality and morbidity
Dr. Seksom highlighted that birthweights decrease with increasing sow prolificacy. He stated that “piglets should be divided into groups with similar body weights at weaning” and that “a key objective for successful weaning is a piglet that weighs a minimum of 6-6.5 kg at three weeks of age, and that less than 25% of the piglets have a weight of ≤5.9 kg.”
Sow body condition
Sows should be fed to feed to body condition score (BCS), not a fixed amount of feed. Ideally, the sows have a BCS of 2.75 (the sow’s backbone is visible, and the tips of the short ribs can be felt but are smooth) or 3.0 (well-rounded appearance, hips, and spine can only be felt with firm pressure) at 12 weeks of pregnancy, so we can feed more in the last month to achieve a BCS of 3-3.25 at farrowing. This is essential to ensure that sows have sufficient energy reserves for lactation and overall health.
Target body condition score – 2.75 at three months of gestation
Feed intake must be increased gradually during the last month of gestation as most fetal growth and mammary gland development occur during this period. This may involve raising energy-dense feeds or adjusting protein levels as needed.
Dr. Seksom stressed that “nutrition is not just the feed; it’s about feeding as well. To feed sows to BCS, assessments of BCS should be done regularly throughout gestation, ideally every 2-4 weeks. This allows for timely adjustments in feeding based on individual sow’s needs. Ensure that staff are trained one-on-one to accurately assess the body condition of sows. This includes recognizing the visual and tactile indicators of different scores and understanding how BCS impacts reproductive performance, longevity, and overall farm profitability.”
After farrowing, the sows must be monitored closely for any signs of excessive weight loss and feeding strategies adjusted accordingly to support recovery and lactation needs.
Piglet diarrhea
Many factors cause diarrhea and must be thoroughly investigated. For bacteria-caused diarrhea, Dr. Seksom advised a good hygiene program, whereas for viral causes, a vaccination program is essential. However, he emphasized that “for a vaccination program, you can’t just copy from another farm; it needs to be created specifically using the titers for diseases on your farm.”
Swine influenza is an often-overlooked cause of diarrhea in piglets. While it is primarily recognized for causing respiratory issues, the virus can also lead to scours in the first two weeks of piglets’ life. So, sows should be checked for symptoms of swine influenza (such as nasal discharge, sneezing and coughing, and inappetence) before farrowing. If necessary, they must be treated with paracetamol to reduce fever and symptoms.
Main disease causes of pre-weaning diarrhea
Nursery period
Mortality level
Days 1-3
Days 3-7
Days 7-14
Days 14-21
Agalactia
√
√
√
√
Moderate
Clostridia
√
√
√
High
Coccidiosis
√
√
√
Low
E. coli
√
√
√
Moderate
PED
√
√
√
Variable
PRRS
√
√
√
√
Variable
Rotavirus
√
√
Low
TGE
√
√
√
√
High
Influenza
√
√
Low
Ensuring colostrum intake
The intake of an adequate quantity of colostrum is crucial for piglets to be protected during the first days of life. Best practices to ensure that piglets get 250 mL of colostrum include:
Teat access – if a sow has a large litter or is unable to nurse all her piglets effectively, consider split suckling by separating larger, more vigorous piglets from the litter for a couple of hours after birth. This allows smaller or weaker piglets better access to the udder without competition. Syringe-feeding colostrum to smaller piglets is also effective.
Early access – six hours after farrowing, the quality of colostrum begins to decline significantly. Additionally, the piglet can only absorb intact large IgG molecules, the major source of passive immunity, during the first 24 h after birth, prior to gut closure. In any case, by this time, the sow will start producing milk and not colostrum.
Sow behavior – if a sow experiences pain or discomfort from injuries caused by her piglets’ teeth, she may become less willing to allow them to nurse, leading to delays in colostrum intake. Genetic background influences maternal behavior significantly. For example, some breeds exhibit stronger maternal instincts and better nursing
behaviors than others. Selecting sows with proven good maternal traits can lead to improved outcomes in piglet survival and growth.
Drafts – newborn piglets are born with low fat reserves and are highly susceptible to hypothermia. Drafts significantly impact the effective temperature experienced by piglets.
Staff training – Staff must be trained to recognize signs of distress in both sows and piglets; training in techniques enables them to assist with nursing and feeding, which is crucial for timely interventions.
Weaning is a process, not just a one-time event
Research has shown that heavier piglets at weaning have better lifetime performance than lighter ones. Weaning weight is a more accurate indication of post-weaning growth than either birth weight or age. It is, therefore, important to establish the weaner immediately post-weaning to maintain growth rates, reduce pen variation, and lessen the amount of ‘tail-enders’ at the point of sale.
Dr. Seksom emphasized that “viewing weaning merely as a single event, rather than a process, overlooks the complexities involved in ensuring a smooth transition for the animals. He advocated for a comprehensive approach to weaning that includes the shown well-planned steps to support piglets during this critical phase. If the weaning process is managed effectively, you can significantly reduce the need for antibiotics.”
Conclusion
“By integrating these holistic management strategies, pig producers can effectively raise pigs without antibiotics while promoting animal health, improving productivity, and addressing consumer concerns about antibiotic use in livestock production,” summarized Dr. Seksom.
EW Nutrition’s Swine Academy took place in Ho Chi Minh City and Bangkok in October 2024. Dr. Seksom Attamangkune, a leading expert in the nutrition and management of pigs in tropical conditions and former Head of the Animal Science Department and Dean of the Faculty of Agriculture at Kamphaeng Saen, Kasetsart University, was a reputable guest speaker at this event.
Antimicrobial resistance in animal production workers, a serious challenge
With 73% of human-use antibiotics also used in food-animal production, antimicrobial resistance (AMR) is a pressing global health concern, particularly in contexts where humans and animals are in close proximity, such as in animal production facilities. This issue is exacerbated by the widespread use of antibiotics in livestock farming, which not only promotes resistance in bacteria but also poses direct risks to farm workers.
Antimicrobial resistance in farm workers in Denmark
In Denmark, a country renowned for its robust agricultural monitoring systems, significant strides have been made in tracking AMR. A comprehensive report from 2015 emphasized the occurrence of antimicrobial-resistant bacteria, particularly in livestock-associated methicillin-resistant Staphylococcus aureus (LA-MRSA). The Danish Integrated Antimicrobial Resistance Monitoring and Research Program (DANMAP) highlighted that farm workers frequently came into contact with these resistant pathogens, which posed occupational hazards and public health challenges (Bager et al., 2015). The program found that 88% of pigs carried LA-MRSA, and farm workers had significantly elevated exposure risks, particularly in intensive swine operations (DANMAP 2015 Report).
Antimicrobial resistance in farm workers in the US
Studies in the United States have revealed even more alarming statistics. Farm workers in intensive animal farming environments were found to be 32 times more likely to develop antibiotic-resistant infections than the general population. This increased risk was attributed to prolonged exposure to resistant bacteria and antibiotic residues in animal feed and the environment (Silbergeld et al., 2008). The close interaction between humans and animals in confined spaces fosters the transfer of resistant genes, making these workers a vulnerable group.
Mechanisms of resistance spread
The spread of AMR from livestock to humans can occur through several pathways:
Direct contact: Handling animals and exposure to manure or bodily fluids.
Contaminated food: Consumption of undercooked or improperly handled meat products.
Environmental contamination: Water and soil contaminated with antibiotics or resistant bacteria.
What can be done?
Even in countries where antimicrobials reduction legislation has been in place for almost two decades, such as Germany or Sweden, new resistance cases are constantly discovered. In supermarkets around the world, meat contaminated with antibiotic-resistant superbugs is still a common occurrence. And in antibiotic resistance hot spots, “from 2000 to 2018, P50 increased from 0.15 to 0.41 in chickens—meaning that 4 of 10 antibiotics used in chickens had resistance levels higher than 50%. P50 rose from 0.13 to 0.43 in pigs and plateaued between 0.12 and 0.23 in cattle” (Dall, 2019). These hot spots are spread across the globe, from south and northeast India, northeast China, north Pakistan, Iran, and Turkey, to the south coast of Brazil, Egypt, the Red River Delta in Vietnam, and areas surrounding Mexico City, Johannesburg, and more recently Kenya and Morocco.
Globally, antimicrobial use in animals is projected to increase by 67% by 2030, especially in low- and middle-income countries where regulatory frameworks are weaker. Denmark provides a successful model for mitigating these risks. Policies such as the “Yellow Card” scheme have reduced antibiotic use in pigs by promoting alternative husbandry practices and strict monitoring. This approach has also reduced the prevalence of resistant bacteria in animal populations, offering a replicable strategy for other nations (Alban et al., 2017).
Recommendations for mitigation
Strengthening surveillance: Programs like DANMAP should be implemented globally to monitor antibiotic usage and resistance trends in animals and humans.
Reducing antibiotic use: Phasing out non-therapeutic uses of antibiotics, particularly as growth promoters, and avoiding Critically Important Antimicrobials for Human Medicine.
Protecting workers: Providing personal protective equipment (PPE) and regular health screenings for farm workers.
Public awareness: Educating communities about the risks of AMR and promoting safe food handling practices.
The evidence from Denmark and the U.S. underscores the urgent need to address AMR in animal production settings. Protecting farm workers from AMR not only safeguards their health but also prevents the spread of resistant pathogens across the wider public.
References
Bager, F., et al. (2015). DANMAP 2014: Use of antimicrobial agents and occurrence of antimicrobial resistance in bacteria from food animals, food, and humans in Denmark. Retrieved from DANMAP Report.
Silbergeld, E. K., Graham, J., & Price, L. B. (2008). Industrial food animal production, antimicrobial resistance, and human health. Annual Review of Public Health, 29(1), 151-169.
Alban, L., et al. (2017). Assessment of the risk to public health due to use of antimicrobials in pigs—An example of pleuromutilins in Denmark. Frontiers in Veterinary Science, 4, 74. DOI.
Global antimicrobial use in livestock farming: A revised estimate
Antimicrobial resistance (AMR) poses a significant threat to global health, driven by the overuse and misuse of antibiotics in both human medicine and livestock farming. In livestock farming, antimicrobials are still used extensively for therapeutic and non-therapeutic purposes. However, estimates of the quantities used per species are notoriously hard to derive from fragmented, incomplete, or unstandardized data around the world.
A recent article (“Global antimicrobial use in livestock farming: an estimate for cattle, chickens, and pigs”, Animal, 18(2), 2024) attempts to update the figures by estimating global biomass at treatment of cattle, pigs, and chickens, considering distinct weight categories for each species in biomass calculation, and using the European Medicines Agency’s weight standards for the animal categories. With these more refined calculations, authors Zahra Ardakani, Maurizio Aragrande, and Massino Canali aim to provide a more accurate estimate of global antimicrobial use (AMU) in cattle, chickens, and pigs. Understanding these patterns is crucial for addressing AMR and developing strategies for sustainable livestock management.
Key Findings
The study estimates that the global annual AMU for cattle, chickens, and pigs amounts to 76,060 tons of antimicrobial active ingredients. This is a significant revision from previous estimates due to a more detailed evaluation of animal weights and categories:
1. Cattle: 40,697 tons (53.5% of total AMU)
2. Pigs: 31,120 tons (40.9% of total AMU)
3. Chickens: 4,243 tons (5.6% of total AMU)
Figure 1: Distribution of global antimicrobial use among cattle, pigs, and chickens.
Methodology
The study utilizes the concept of Population Correction Units (PCU) to estimate antimicrobial usage, taking into account the weight and category of livestock at the time of treatment. This method differs from previous approaches that relied on live weight at slaughter, providing a more accurate representation of AMU.
The PCU is calculated by multiplying the number of animals by their average weight during treatment. This approach allows for differentiation by age and sex, which is particularly important for species like cattle and pigs.
Figure 2: (a) Changes in global PCU (million tonnes), (b) changes in global antibiotic use in mg per PCU, and (c) changes in global AMU (thousand tonnes) for cattle, chickens, and pigs; between 2010 and 2020. Abbreviations: PCU = Population Correction Unit; AMU = Antibiotic Use.
Figure 2: (a) Changes in global PCU (million tonnes), (b) changes in global antibiotic use in mg per PCU, and (c) changes in global AMU (thousand tonnes) for cattle, chickens, and pigs; between 2010 and 2020. Abbreviations: PCU = Population Correction Unit; AMU = Antibiotic Use.
Study shows lower AMU than previous estimates
The study highlights a significant shift in AMU patterns, with chickens showing a remarkable decrease in antimicrobial use despite increased production. This is indicative of improved management and more responsible use of antibiotics in the poultry industry.
The lower AMU in cattle and pigs, compared to previous estimates, underscores the importance of considering animal age and weight at treatment. These findings align closely with World Organization for Animal Health (WOAH) estimates, validating the methodology.
However, the study also acknowledges limitations, including reliance on European standards for average weight at treatment, which may not reflect global variations. Additionally, the lack of comprehensive global data on veterinary antibiotics presents challenges in creating fully accurate estimates.
Corrected estimate highlights improved production advances
This study provides a revised and potentially more accurate estimate of global antimicrobial use in livestock. By accounting for the weight and treatment categories of animals, it offers insights that could guide policy and management practices to mitigate the spread of antimicrobial resistance.
The article also indicates that the industry may have over-estimated antimicrobial usage in livestock and, just as importantly, that antimicrobial use has been kept in check or even reduced, despite increases in farmed animal headcounts. The lower usage is likely due to regulatory oversight and improvements in alternative methods to control and mitigate health challenges.
Can phytogenics have a meaningful effect in coccidiosis control?
by Madalina Diaconu, Global Manager Gut Health, EW Nutrition
Coccidiosis, caused by Eimeria spp., is a major challenge in poultry production, leading to significant economic losses. Historically, control strategies have relied on chemical anticoccidials and ionophores. However, the emergence of drug-resistant Eimeria strains and consumer concerns about chemical residues necessitate alternative solutions. Phytogenics, especially tannins and saponins, offer promising natural solutions to be included in programs for coccidiosis control. More and more independent research highlights the potential of these natural compounds to enhance poultry health and productivity.
Efficacy of Tannins and Saponins in Coccidiosis Control
Phytogenics are plant-derived bioactive compounds known for their antimicrobial, antioxidant, and immunomodulatory properties. Among these, tannins and saponins have shown particular promise in supporting coccidiosis control.
The challenge: Preventing the spread of infections and mitigating subclinicial coccidiosis before it reaches this stage.
Tannins
Tannins are polyphenolic compounds found in various plants. They exhibit strong antimicrobial activity by binding to proteins and metal ions, disrupting microbial cell membranes, and inhibiting enzymatic activity.
Anticoccidial Activity: Tannins have been shown to interfere with the life cycle of Eimeria. Studies demonstrate that tannins can reduce oocyst shedding and intestinal lesion scores in infected birds (Abbas et al., 2017).
Immune Modulation: Tannins enhance immune responses by promoting the proliferation of lymphocytes and the production of antibodies, which help in the clearance of Eimeria infections (Redondo et al., 2021).
Saponins
Saponins are glycosides with surfactant properties, capable of lysing cell membranes of pathogens. They also stimulate immune responses, enhancing the host’s ability to fight infections.
Membrane Disruption: Saponins disrupt the cell membranes of Eimeria, leading to reduced parasite viability and replication (Githiori et al., 2004).
Immune Enhancement: Saponins stimulate the production of cytokines and enhance the activity of macrophages, improving the overall immune response against coccidiosis (Zhai et al., 2014).
Independent Research Evidences Phytogenics’s Role in Supporting Programs for Coccidiosis Control
Numerous studies have evaluated the efficacy of phytogenics in coccidiosis control. Here, we highlight key findings from peer-reviewed research:
Abbas et al. (2012): This study reviewed various botanicals and their effects on Eimeria species in poultry. The authors concluded that tannins and saponins significantly reduce oocyst shedding and lesion scores, comparable to conventional anticoccidials.
Allen et al. (1997): The authors investigated the use of dietary saponins in controlling Eimeria acervulina infections. The study found that saponin-treated birds exhibited lower oocyst counts and improved weight gain compared to untreated controls.
Masood et al. (2013): This study explored the role of natural antioxidants, including tannins, in controlling coccidiosis. The results indicated that tannins reduced oxidative stress and improved intestinal health, leading to better performance in broiler chickens.
Idris et al. (2017): The researchers assessed the potential of saponin-rich plant extracts against avian coccidiosis. The findings demonstrated significant reductions in oocyst output and lesion severity, highlighting the potential of saponins as effective anticoccidials.
Hailat et al. (2023): The researchers studied three phytogenic formulations against a control group with chemical drugs. The study concluded that phytogenic blends can be safely used as alternatives to the chemically synthesized drugs, either alone or in a shuttle program, for the control of poultry coccidiosis.
El-Shall et al. (2021): This review article highlights research findings on phytogenic compounds which showed preventive, therapeutic, or immuno-modulating effects against coccidiosis.
Despite initial skepticism, the growing body of evidence supports the efficacy of phytogenics in supporting coccidiosis control. Tannins and saponins, in particular, have shown significant potential in reducing parasite load, improving intestinal health, and enhancing immune responses. These natural compounds offer several advantages over traditional chemical treatments, including lower risk of resistance development and absence of harmful residues in meat products.
Challenges and Promises
While the efficacy of phytogenics is well-supported, challenges remain, especially with lower-quality products that may display variability in plant extract composition, in their standardization of doses, and in ensuring consistent quality. At the same time, these compounds are not silver bullets, and no producer should make unreasonable claims.
As far as the mode of action is concerned, the evidence is becoming clear: phytogenics, particularly tannins and saponins, are effective in mitigating gut health challenges and supporting bird performance when challenged. Their natural origin, coupled with potent antimicrobial and immunomodulatory properties, makes them suitable for sustainable poultry production. As the poultry industry seeks to reduce reliance on chemical drugs, phytogenics represent a viable and promising solution.
References
Abbas, R. Z., Iqbal, Z., Blake, D., Khan, M. N., & Saleemi, M. K. (2011). “Anticoccidial drug resistance in fowl coccidia: the state of play revisited”. World’s Poultry Science Journal, 67(2), 337-350. https://doi.org/10.1017/S004393391100033X
Allen, P. C., Danforth, H. D., & Levander, O. A. (1997). “Interaction of dietary flaxseed with coccidia infections in chickens”. Poultry Science, 76(6), 822-828. https://doi.org/10.1093/ps/76.6.822
El-Shall, N.A., El-Hack, M.E.A., et al. (2022). “Phytochemical control of poultry coccidiosis: a review”. Poultry Science, 101(1) 101542. https://doi.org/10.1016/j.psj.2021.101542
Idris, M., Abbas, R. Z., Masood, S., Rehman, T., Farooq, U., Babar, W., Hussain, R., Raza, A., & Riaz, U. (2017). “The potential of antioxidant rich essential oils against avian coccidiosis”. World’s Poultry Science Journal, 73(1), 89-104. https://doi.org/10.1017/S0043933916000787
Hailat, A.M., Abdelqader, A.M., & Gharaibeh, M.H. (2023). “Efficacy of Phyto-Genic Products to Control Field Coccidiosis in Broiler Chickens”. International Journal of Veterinary Science, 13(3), 266-272. https://doi.org/10.47278/journal.ijvs/2023.099
Masood, S., Abbas, R. Z., Iqbal, Z., Mansoor, M. K., Sindhu, Z. U. D., & Zia, M. A. (2013). “Role of natural antioxidants for the control of coccidiosis in poultry”. Pakistan Veterinary Journal, 33(4), 401-407.
Redondo, L. M., Chacana, P. A., Dominguez, J. E., & Miyakawa, M. E. (2021). “Perspectives in the use of tannins as alternative to antimicrobial growth promoter factors in poultry”. Frontiers in Microbiology, 12, 641949. https://doi.org/10.3389/fmicb.2021.641949
Zhai, H., Liu, H., Wang, S., Wu, J., & Kluenter, A. M. (2014). “Potential of essential oils for poultry and pigs”. Animal Nutrition, 2(4), 196-202. https://doi.org/10.1016/j.aninu.2016.12.004
Sustainable use of veterinary antimicrobials in Europe: EEA report
The European Environment Agency (EEA) has recently published a briefing detailing the impact of veterinary antimicrobials on Europe’s environment. Positive developments are to be applauded, however they do not tell the whole story.
The use of antimicrobials for farmed animals and in aquaculture decreased by around 28% between 2018 and 2022. Nonetheless, the rate of antimicrobial resistance continues to rise around the world, including as an important cause of death in the European Economic Area (the EEA includes all EU countries, as well as Norway, Lichtenstein, and Iceland). At present, antimicrobial-resistant infections are estimated to caused 35,000 deaths per year in the European Union. For reference, in the EU, traffic accidents cause around 20,000 deaths per year.
A large number of EU guidelines, policies, and regulations attempt to control and monitor the use of antimicrobials in food-producing animals. This makes the regulatory landscape somewhat confusing, especially that many implementation details are still left to the states.
Figure 1. Overview of the EU regulatory framework applicable to antimicrobials used in food-producing animals
One of the results of unequal implementation is that there is no standardized way of tracking the actual use of antimicrobials in food-producing animals. To collect the numbers, the EEA has used proxy numbers, especially antimicrobial sales data and self-reported data. With such broad strokes, it is to be expected that the actual figures might be higher.
Lower numbers…
According to the European Surveillance of Veterinary Antimicrobial Consumption (ESVAC) database, in the European Economic Area countries, plus Switzerland and the UK, total antimicrobial consumption for farmed animals and aquaculture was estimated at 73.9 mg/PCU* in 2022. This signifies a 30% reduction over 5 years.
In numbers: 4,458 tons of active substances were sold in one years for farmed animals & aquaculture.
*PCU represents a population correction unit (PCU). The PCU takes into account the population and relative weight of animals and “is used the normalize antimicrobial sales data for the size of the animal population that could potentially be treated with these substances. Using this methodology, 1 PCU corresponds to 1 kilogram of animal biomass”.
…But higher risks
In 2021, total antimicrobial consumption in humans – measured in 28 European countries – was estimated at 125.0 mg/kg. This number has unfortunately not gone down. What is worse: a much larger volume of antimicrobials is sold for food-producing animals than for human medicine. Which means that, relative to the total population, the impact of veterinary-use antimicrobials remains disproportionately large.
Moreover, two outliers (Poland and Lithuania) exhibited a worrying increasing trend, showing that no good development is irreversible. The EEA also highlights this danger, in the context of growing global consumption of animal protein. Increased demand “may put pressure on farmers to adopt intensive production practices that require increased use of antimicrobials”. The use of antimicrobials elsewhere in the world may lead to impacts in Europe, “not just by theoretically exposing consumers to antimicrobial residues but also by contributing to rising global rates of drug-resistant pathogens and infections”.
Figure 2. Sales by EU Member States in 2018 vs. 2022
Because of declining livestock populations in the last few years, while demand remained constant, EU-27 imports of animal products more than doubled between 2002 and 2022. There are no reliable global data on the veterinary use of antimicrobials, however it is generally believed that over 70% of antimicrobials sold globally may be used in for animal protein production.
Data collected by the World Organization for Animal Health (WOAH) from its member countries suggest that, between 2017-2019, “the use of antimicrobials in animals decreased by 25% in the Asia, Far East and Oceania regions, while it increased in Africa (+45%) and the Americas (+5%). Despite these partial improvements, a recent study forecasted that global use of antimicrobials in food-producing animals could rise by 8% in 2030, compared to 2020 levels (Mulchandani et al., 2023)”.
Quick summary
Reduction in Antimicrobial Use: There has been a significant reduction in the use of antimicrobials in farming and aquaculture across the EU. From 2018 to 2022, there was a decrease of approximately 28%, which aligns with the EU’s Farm to Fork strategy targeting a 50% reduction by 2030.
Antimicrobial Resistance (AMR): Despite the decrease in use, antimicrobial resistance remains a severe public health threat, causing an estimated 35,000 deaths annually in the European Economic Area. The resistance is attributed to the use of antimicrobials, which – as has been widely documented and discussed – can promote the evolution of resistant microorganisms.
Environmental Impact: The briefing underscores significant knowledge gaps in monitoring antimicrobial residues, resistant bacteria, and resistance genes in the environment. Improved surveillance could help identify pollution hotspots and assess the impact of reduction measures.
Regulatory and Policy Framework: The EU has implemented several policies to regulate the use of antimicrobials, including banning their use as growth promoters and setting stricter conditions for prescriptions. These measures are crucial for managing the risk of AMR.
Further efforts are needed to decrease the reliance on antimicrobials in food production. These include enhanced monitoring, promoting alternative practices in animal farming, and better animal welfare and biosecurity measures.
While improvements are clear and commendable in the EU-27 states, increased antimicrobial usage in some EU countries and in various areas around the world represent a significant concern.
For further details on the use of veterinary antimicrobials in Europe’s environment, you can refer to the EEA’s full report.
Organic acids can play a crucial role in zinc oxide replacement
Dr. Inge Heinzl, Editor EW Nutrition & Juan Antonio Mesonero Escuredo, GTM Swine/GPM Organic Acids EW Nutrition
The use of high levels of Zinc Oxide (ZnO) in the EU before 2022 was one of the most common methods to prevent postweaning diarrhea (PWD) in pig production. Pharmacologically high levels of ZnO (2000-3000 ppm) increase growth and reduce the incidence of enteric bacterial diseases such as post-weaning diarrhea (PWD)( Carlson et al., 1999; Hill et al., 2000; Hill et al., 2001; Poulsen & Larsen, 1995; De Mille et al., 2019).
However, ZnO showed adverse effects, such as the accumulation of heavy metal in the environment, the risk for antimicrobial resistance (AMR), and problems of mineral toxicity and adverse growth effects when feeding it longer than 28 days (Jensen et al., 2018; Cavaco et al., 2011; Vahjen, 2015; Romeo et al., 2014; Burrough et al., 2019). To replace ZnO in pig production, let us first look at its positive effects to know what we must compensate for.
ZnO has a multifactorial mode of action
ZnO shows several beneficial characteristics that positively influence gut health, the immune system, digestion, and, therefore, also overall health and growth performance.
Figure 1. Beneficial effects and ZnO mode of action in postweaning piglets
1. ZnO acts as an antimicrobial
Concerning the antimicrobial effects of ZnO, different possible modes of action are discussed:
ZnO in high dosages generates reactive oxygen species (ROS) that can damage the bacterial cell walls (Pasquet et al., 2014)
The death of the bacterial cell due to direct contact of the metallic Zn to the cell (Shearier et al., 2016)
Intrinsic antimicrobial properties of the ZnO2+ ions after dissociation. The uptake of zinc into cells is regulated by homeostasis. A concentration of the ZnO2+ ions higher than the optimal level of 10-7 to 10-5 M (depending on the microbial strain) allows the invasion of Zn2+ ions into the cell, and the zinc starts to be cytotoxic (Sugarman, 1983; Borovanský et al., 1989).
ZnO shows activity against, e.g., Staphylococcus aureus, Pseudomonas aeruginosa, E. coli,Streptococcus pyogenes, and other enterobacteria (Ann et al., 2014; Vahjen et al., 2016). However, Roselli et al. (2003) did not see a viability-decreasing effect of ZnO on ETEC.
2. ZnO modulates the immune system
Besides fighting pathogenic organisms as described in the previous chapter and supporting the immune system, ZnO is an essential trace element and has a vital role in the immune system. ZnO improves the innate immune response, increasing phagocytosis and oxidative bursts from macrophages and neutrophils. It also ameliorates the adaptative immune response by increasing the number of T lymphocytes (T cells) in general and regulatory T lymphocytes (T-regs) in particular. These cells control the immune response and inflammation (Kloubert et al., 2018). Macrophage capacity for phagocytosis (Ercan and Bor, 1991) and to kill parasites (Wirth et al., 1989), and also the killing activity of natural killer cells depends on Zn (Rolles et al., 2018). By reducing bacterial adhesion and blocking bacterial invasion, ZnO disburdens the immune system (Roselli et al., 2003).
ZnO reduces the expression of several proinflammatory cytokines induced by ETEC (Roselli et al., 2003). Several studies have also shown a modulation effect on intestinal inflammation, decreasing levels of IFN-γ, TNF-α, IL-1ß and IL-6, all pro-inflammatory, in piglets supplemented with ZnO (Zhu et al., 2017; Grilli et al., 2015).
3. ZnO improves digestion and promotes growth
Besides protecting young piglets against diarrhea, the goal is to make them grow optimally. For this target, an efficient digestion and a high absorption of nutrients is essential. Stimulating diverse pancreatic enzymes such as amylase, carboxypeptidase A, trypsin, chymotrypsin, and lipase increases digestibility (Hedemann et al., 2006; Pieper et al., 2015). However, Pieper et al. (2015) also showed that a long-term supply of very high dietary zinc triggers oxidative stress in the pancreas of piglets.
By stimulating the secretion of ghrelin at the stomach level and thereby promoting the release of insulin-like growth factor (IGF-1) and cholecystokinin (CCK), ZnO enhances muscle protein synthesis, cell proliferation, and feed intake (Yin et al., 2009; MacDonald et al., 2000)).
The result of improved digestion is increased body weight and average daily gain, which can be seen, e.g., in a study by Zhu et al. (2017).
4. ZnO protects the intestinal morphology
ZnO prevents the decrease of the trans-endothelial electrical resistance (TEER), usually occurring in the case of inflammation, by downregulating TNF-α and IFN-γ. TNF-α, as well as IFN-γ, increase the permeability of the epithelial tight junctions and, therefore, the intestinal barrier (Al-Sadi et al., 2009).
The enterotrophic and anti-apoptotic effect of ZnO is reflected by a higher number of proliferating and PCNA-positive cells and an increased mucosa surface in the ileum (higher villi, higher villi/crypt ratio)(Grilli et al., 2015). Zhu et al. (2017) also saw an increase in villus height in the duodenum and ileum and a decrease in crypt depth in the duodenum due to the application of 3000 mg of ZnO/kg. Additionally, they could notice a significant (P<0.05) upregulation of the mRNA expression of the zonula occludens-1 and occluding in the mucosa of the jejunum of weaned piglets.
In a trial conducted by Roselli et al. (2003), the supplementation of 0.2 mmol/L ZnO prevented the disruption of the membrane integrity when human Caco-2 enterocytes were challenged with ETEC.
5. ZnO acts antioxidant
The antioxidant effect of ZnO was shown in a study conducted by Zhu et al., 2017. They could demonstrate that the concentration of malondialdehyde (MDA), a marker for lipid peroxidation, decreased on day 14 or 28, and the total concentration of superoxide dismutase (SOD), comprising enzymes that transform harmful superoxide anions into hydrogen peroxide, increased on day 14 (P<0.05). Additionally, Zn is an essential ion for the catalytic action of these enzymes.
Which positive effects of ZnO can be covered by organic acids (OAs)?
1. OAs act antimicrobial
OAs, on the one hand, lower the pH in the gastrointestinal tract. Some pathogenic bacteria are susceptible to low pH. At a pH<5, the proliferation of, e.g., Salmonella, E. coli, and Clostridium is minimized. The good thing is that some beneficial bacteria, such as lactobacilli or bifidobacteria, survive as they are acid-tolerant. The lactobacilli, on their side, can produce hydrogen peroxide, which inhibits, e.g., Staphylococcus aureus or Pseudomonas spp. (Juven and Pierson, 1996).
Besides this more indirect mode of action, a more direct one is also possible: Owing to their lipophilic character, the undissociated form of OAs can pass the bacterial membrane (Partanen and Mroz, 1999). The lower the external pH, the more undissociated acid is available for invading the microbial cells. Inside the cell, the pH is higher than outside, and the OA dissociates. The release of hydrogen ions leads to a decrease in the internal pH of the cell and to a depressed cell metabolism. To get back to “normal conditions”, the cell expels protons. However, this is an energy-consuming process; longer exposure to OAs leads to cell death. The anion remaining in the cell, when removing the protons, disturbs the cell’s metabolic processes and participates in killing the bacterium.
These theoretical effects could be shown in a practical trial by Ahmed et al. (2014). He fed citric acid (0.5 %) and a blend of acidifiers composed of formic, propionic, lactic, and phosphoric acid + SiO2 (0.4 %) and saw a reduction in fecal counts of Salmonella and E. coli for both groups.
2. OAs modulate the immune system
The immune system is essential in the pig’s life, especially around weaning. Organic acids have been shown to support or stimulate the immune system. Citric acid (0.5%), as well as the blend of acidifiers mentioned before (Ahmed et al., 2014), significantly increased the level of serum IgG. IgG is part of the humoral immune system. They mark foreign substances to be eliminated by other defense systems.
Ren et al. (2019) could demonstrate a decrease in plasma tumor necrosis factor-α that regulates the activity of diverse immune cells. He also found lower interferon-γ and interleukin (Il)-1ß values in the OA group than in the control group after the challenge with ETEC. This trial shows that inflammatory response can be mitigated through the addition of organic acids.
3. OAs improve digestion and promote growth
In piglets, the acidity in the stomach is responsible for the activation and stimulation of certain enzymes. Additionally, it keeps the feed in the stomach for a longer time. Both effects lead to better digestion of the feed.
In the stomach, the conversion of pepsinogen to pepsin, which is responsible for protein digestion, is catalyzed under acid conditions (Sanny et al., 1975)group. Pepsin works optimally at two pH levels: pH 2 and pH 3.5 (Taylor, 1959). With increasing pH, the activity decreases; at pH 6, it stops. Therefore, a high pH can lead to poor digestion and undigested protein arriving in the intestine.
These final products of pepsin protein digestion are needed in the lower parts of the GIT to stimulate the secretion of pancreatic proteolytic enzymes. If they do not arrive, the enzymes are not activated, and the inadequate protein digestion continues. Additionally, gastric acid is the primary stimulant for bicarbonate secretion in the pancreas, neutralizing gastric acid and providing an optimal pH environment for the digestive enzymes working in the duodenum.
As already mentioned, the pH in the stomach influences the transport of digesta. The amount of digesta being transferred from the stomach to the small intestine is related to the acidity of the chyme leaving the stomach and arriving in the small intestine. Emptying of the stomach can only take place when the duodenal chyme can be neutralized by pancreatic or other secretions (Pohl et al., 2008); so, acid-sensitive receptors provide feedback regulation and a higher pH in the stomach leads to a faster transport of the digesta and a worse feed digestion.
4. OAs protect the intestinal morphology
Maintaining an intact gut mucosa with a high surface area is crucial for optimal nutrient absorption. Research suggests organic acids play a significant role in improving mucosal health:
Butyric acid promotes epithelial cell proliferation, as demonstrated in an in vitro pig hindgut mucosa study (Sakata et al., 1995). Fumaric acid, serving as an energy source, may locally enhance small intestinal mucosal growth, aiding in post-weaning epithelial cells’ recovery and increasing absorptive surface and digestive capacity (Blank et al., 1999). Sodium butyrate supplementation at low doses influences gastric morphology and function, thickening the stomach mucosa and enhancing mucosal maturation and differentiation (Mazzoni et al., 2008).
Studies show that organic acids affect gut morphology, with a mixture of short-chain and mid-chain fatty acids leading to longer villi (Ferrara et al., 2016) and Na-butyrate supplementation increasing crypt depth and villi length in the distal jejunum and ileum (Kotunia et al., 2004). However, the villi length and mucosa thickness in the duodenum were reduced. Dietary sodium butyrate has been linked to increased microvilli length and cecal crypt depth in pigs (Gálfi and Bokori, 1990).
5. OAs show antioxidant activity
The last characteristic, the antioxidant effect, cannot be provided at the same level as with ZnO; however, Zhang et al. (2019) attest to OAs a certain antioxidant activity. Oxalic, citric, acetic, malic, and succinic acids, which were extracted from Camellia oleifera, also showed good antioxidant activity in a trial conducted by Zhang et al. (2020).
Organic acids are an excellent tool to compensate for the ban on ZnO
The article shows that organic acids have similar positive effects as zinc oxide. They act antimicrobial, modulate the immune system, maintain the gut morphology, fight pathogenic microbes, and also act – slightly – antioxidant. Additionally, they have a significant advantage: they are not harmful to the environment. Organic acids used in the proper pH range and combination are good tools for replacing zinc oxide.