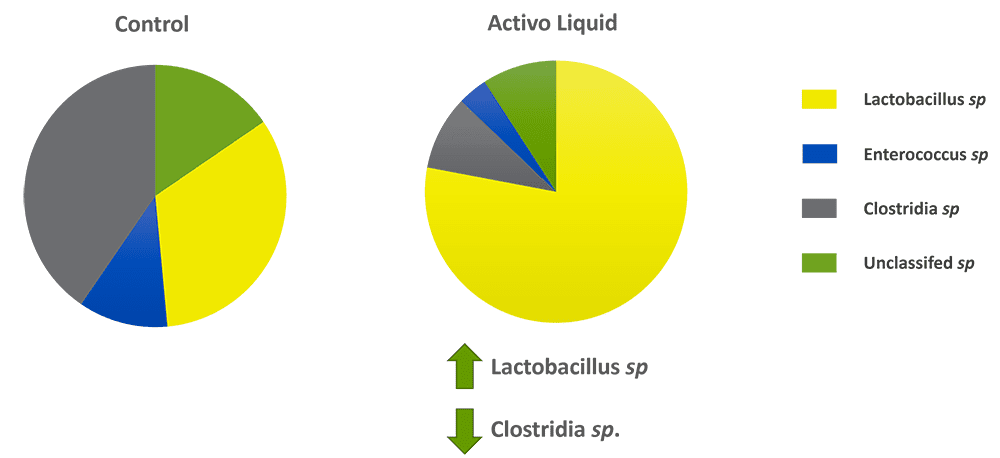How nutritionists can adjust feed formulation costs – and still keep all value
By Technical Team EW Nutrition
More than five months after the start of the war in Ukraine, we are facing severe challenges in many aspects of our daily lives. The livestock industry is not excepted: the conflict affects the availability and prices of grains, fertilizers, oil, and biofuel, with the latter two having a direct impact on freight rate and shipping time.
Livestock producers face challenging times and must be rational and creative to continue managing profitable operations.
Increased prices for feed materials and energy
Which strategies are available?
Maintaining performance and profitability during price hikes generally requests a combination of different nutritional strategies to, at least partially, compensate for the higher costs:
using alternative feed ingredients such as by-products (while considering their limitations in terms of inclusion and quality)
eliminating or reducing safety bands (especially to already used by-products)
revising the use of feed additives according to current challenges in feed ingredients and at farms
following on the previous one: strategic uses of enzymes, emulsifiers, and feed additives that may help the animals to improve their FCR.
By-products can partly compensate for high feed-ingredient prices
Over the last few decades, industrial and agricultural by-products from crops, vegetables, and fruit processing have been widely evaluated and used in livestock. As a result, many studies have been conducted to determine the nutritional composition of straws and residues from food manufacturing.Now more than ever, by-products are needed since more of the leading agricultural products will be taken to feed the human population instead of animals.
Agricultural by-products can be classified into various types: straws, brans, midds, cakes, meals, and industry residues, among others. Their price is typically lower than traditional energy and protein sources, making them suitable alternative ingredients to potentially reduce the overall cost of the diet.
Wheat milling by-product
The use of by-products – to what must be paid attention?
To guarantee safety and effectiveness, just as with any other feedstuff, it is necessary to check the nutrient composition of the alternative ingredients using feed composition tables and laboratory analysis. Besides the composition and nutrient concentration, the availability of these nutrients and palatability are critical parameters to consider. The feed/animal producer, when purchasing by-products, should:
try to find his by-product sources close to the feed production site to reduce costs with logistics and transportation
collect and test samples right after the by-products are delivered
check if the feed mill is ready to handle and process those ingredients, especially when they are bulky or have flowability issues;
compare the difference in animal performance and cost per unit produced when using traditional grain-soybean meal diets vs. by-product ingredients.
Processing for improving by-products’ quality
The safety and nutritional availability of by-products can be improved by chemical, physical, and biological treatments. Physical processes, such as drying, grinding, peeling, pelleting, extruding, and expanding, increase surface area and can deactivate certain anti-nutritional factors. Biological processes include the use of enzymes and microbial fermentation to tackle anti-nutritional factors and increase the nutritional value and digestibility of by-products.
Be aware of the possible risks of by-products!
With all the economic benefits and positive impacts on animal performance, we must not lose sight of the possible risks and health issues that by-products might imply.
Mycotoxins can be a problem
Most agricultural by-products have a higher moisture content than traditional ingredients. High-fiber and high-moisture materials can quickly become contaminated by molds that produce mycotoxins, undermining animal performance or even leading to death (Juan et al., 2017; Peng et al., 2018).
Fusarium ssp. produce mycotoxins such as fumonisins, trichothecenes (DON, T-2 toxin) or zearalenone
Anti-nutritional factors negatively impact animal health
In addition, some agro-industrial by-products contain anti-nutritional factors (glycoalkaloids, tannins). These substances impair feed digestibility and affect animal performance (Jimenez-Moreno et al., 2019). Also, a high fiber content in the diet containing by-products limits the performance (Pereira et al., 2019).
Additives can help with cost reduction
Due to the increase in feed prices, it is also necessary to review the strategies for using feed additives in animal production. Enzymatic complexes or packages, mannanases, phytases, and xylanases, among others, might be a helpful option to maximize the yield of existing diets. For example, Edward et al. (2000) reviewed the benefits of using phytase for better phosphorus utilization in the diet (a raw material that also suffers from price increases since much of it is imported from China). However, the enzymes must be used properly. Nutritionists trying to create profitable formulations must check the availability of the substrate before including the enzyme in their formulation.
Other feed additives such as toxin binders reduce the exposure of animals to possible increased levels of mycotoxins. Gut health-improving additives such as pro/pre-biotics, phytomolecules, and MCFAs support gut health and performance, achieving similar levels as traditional diets.
These applications should be thoroughly evaluated as the return from their application may be interesting in increased by-products diets.
Using by-products in poultry means balancing several factors
In poultry feeds, using by-products to increase sustainability and cost-reduction is supported by ample research and practice, especially in feed for broilers and laying hens. Research focuses on finding the risks of the inclusion of various by-products and thus the levels at which their inclusion doesn’t hurt health and performance.
In summary, to use by-products in poultry diets, their cost, availability, nutritional composition, anti-nutritional factors, quality, as well as interaction and cost-effectiveness with feed additives (e.g., enzymes, toxin binders) must be considered to avoid or diminish the factors hindering animal health and performance.
Several factors must be considered when using by-products in poultry nutrition
DDGS are a valuable source of proteins – but limited inclusion
DDGS provide protein, energy, water-soluble vitamins, xanthophylls, and linoleic acid (Abd El-Hack et al., 2015). However, it also contains anti-nutritional factors such as non-starch polysaccharides (NSP) (Pedersen et al., 2014). A further disadvantage is their high danger of mycotoxin contamination (Schaafsma et al., 2009), even though Wang et al. (2007) and Damasceno (2020) indicate that up to 16% of DDGS can be included in broilers’ diets without negatively affecting health, performance, and meat characteristics.
Rice brans are one of the main available grain by-products
Rice brans constitute 10% of the paddy rice and thus represent a considerable global volume of the available grain by-products. As a feed ingredient, it is rich in protein, starch, fat, vitamins, and some trace minerals (Sanchez et al., 2019). Due to their susceptibility to oxidation (rancidity) and anti-nutritional factors such as phytase and trypsin inhibitors (Gallinger et al., 2004), the limit recommended for this by-product in poultry is around 10% (Hosseini et al., 2020; Sanchez et al., 2019).
Wheat by-products – optimally used with enzymes
Wheat by-products can also be a substitute for whole grains in poultry feeds; however, their NSP content can affect the viscosity of the digesta (Knudsen, 2014). When combined with enzymes (e.g., xylanase), wheat midds can be included in broiler and layer diets up to 30% without changes in performance (Abudabos, 2011; Salami et al., 2018). Dietary fiber has gained special attention due to its various beneficial effects on poultry. In this direction, moderate amounts of wheat bran – a source of insoluble fiber – have shown improved antioxidant status, gizzard development, intestinal digestive enzyme activities, and morphology in broilers (Shang et al., 2020).
By-products support pigs’ performance
When by-products are fed to pigs, swine nutritionists have reported that many of them can support pig growth and finishing performance and meat quality as well as immune response, milk yield, and milk quality in reproductive animals, among other productive parameters (Yang et al., 2021). For instance, Dong et al. (2019) concluded that, from a nutritional perspective, ingredients such as highland barley, buckwheat, glutinous broomcorn millet, non-glutinous broomcorn millet, and Chinese naked oat could potentially substitute corn in livestock feeding. Or as another example, Liu et al. (2019) suggest in their study that mulberry leaf can contribute to improvements in meat quality, with no adverse effects on the growth performance of finishing pigs. (Dong et al., 2019).
Pigs are susceptible to antinutritional factors
Especially pigs are susceptible to anti-nutritional factors
There are many different types of anti-nutritional factors that work in various ways. In swine feed, common anti-nutritional factors lower protein and amino acid digestibility and increase endogenous amino acid losses (Souffrant, 2001). This effect causes reductions in carcass yield for finishing pigs (Soto et al., 2019).
Options are available to compensate for the higher feed prices
Nutritionists have several options to optimize animal performance in the context of price increases. However, it is necessary to have a more holistic view of the business to know which of all the alternatives are the most suitable for each system. Understanding the strengths and weaknesses of each ingredient and feed additive and considering them in the light of literature and field data will yield the best understanding of how to use them effectively in successful animal production.
Stop endotoxins from decreasing animal performance
By Technical Team EW Nutrition
Find out why endotoxemia threatens animal production and how intelligent toxin mitigation solution SOLIS MAX can support endotoxin management.
Figure 1: Structure of Lipopolysaccharide
The quick guide to endotoxins (LPS) and what to do about them
Lipopolysaccharides (LPS) are a constant challenge for animal production. LPS, which are also known as endotoxins, are the major building blocks of the outer walls of Gram-negative bacteria (see figure 1). Throughout its life cycle, a bacterium releases these molecules upon cell death and lysis. When endotoxins are released into the intestinal lumen of chickens or swine, or in the rumen of polygastric animals, they can cause serious damage to the animal’s health and performance by over-stimulating their immune system.
LPS may induces inflammation and fever, lowering feed intake, and redirecting nutritional resources to the immune response, which results in hindered animal performance.
Endotoxins depress animal performance
One of the biggest issues caused by endotoxemia is that animals reduce their feed intake and show a poor feed conversion rate (FCR). Why does this happen? The productive performance of farm animals (producing milk, eggs, or meat) requires nutrients. An animal also requires a certain baseline amount of nutrients for maintenance, that is, for all activities related to its survival.
As a result of inflammation, endotoxemia leads to a feverish state. Maintenance needs to continue; hence, the energy required for producing heat will be diverted from the nutrients usually spent on production of milk, eggs, meat, etc., and performance suffers. This is amplified because the immune reaction also requires resources (e.g., energy, amino acids, etc. to produce more immune cells).
The inflammation response can result in mitochondrial injury to the intestinal cells, which alter the cellular energy metabolism. This is reflected in changes to the levels in adenosine triphosphate (ATP), the energy “currency” of living cells. A study by Li et al. (2015) observed a respective reduction of 15% and 55% in the ATP levels of the jejunum and ileum of LPS-challenged broilers, compared to the unchallenged control group.
A piglet study by Huntley, Nyachoti, and Patience (2017) found that LPS-challenged pigs retained 15% less of the available metabolizable energy and showed 25% less nutrient deposition (figure 2). These results illustrate how animal performance declines during endotoxemia.
Control treatment (CON) = Pigs fed by a basal diet
Immune system stimulation treatment (ISS) = Pigs given LPS (E. coli serotype 055:B5) injection
A loss of energy retained due to a reduction in available metabolizable energy leads to losses in performance as the amount of energy available for muscle production and fat storage will be lower. Furthermore, the decrease in feed intake creates a further energy deficit concerning production needs.
Endotoxin tolerance
The repeated exposure to LPS leads to the production of anti-inflammatory cytokines, as a reaction of the body to prevent tissue damage due to the excessive inflammation. This immunosuppression during stress may lead to an increased risk of secondary infection and poor vaccination titers.
LPS tolerance, also known as CARS (compensatory anti-inflammatory response syndrome) essentially depresses the immune system to control its activity. This “regulation” can be extremely dangerous as an excessive depression of the immune system leaves the organism exposed to the actual pathogens.
The way forward: Natural endotoxin mitigation with SOLIS MAX
The quantity of Gram-negative bacteria in an animal intestine is considerable; therefore, the danger of immune system over-stimulation through endotoxins cannot be taken lightly. Stress factors – that are not uncommon in animal production – affect the microbiome (favoring gram-negative bacteria) and also decrease the intestinal barrier function, which leads to the passage of LPS into the bloodstream
Animals suffering from endotoxemia are subject to severe metabolic dysfunctions. If they do not perish from septic shock (and most of them do not), they are still likely to show performance losses. Moreover, they at great risk of immunosuppression caused by CARS, the immune system “overdrive” discussed above.
Fortunately, research shows that EW Nutrition’s SOLIS MAX effectively binds bacterial toxins, helping to prevent these scenarios.
In vitro trial shows SOLIS MAX’ effectiveness against bacterial endotoxins
Binding endotoxins in the gastrointestinal tract, especially during stress situations in animal production, can help to mitigate the negative impact of LPS on the animals. It reduces the endotoxins passing into the bloodstream and entering the organism.
SOLIS MAX is a synergistic combination of natural plant extracts, yeast cell walls, and natural clay minerals. An in vitro study conducted at a research facility in Germany evaluated its binding performance for LPS derived from E. coli.
To test the efficacy of SOLIS MAX in binding endotoxins, 0.1% (w/v) of SOLIS MAX was resuspended in endotoxin-free water, with and without a challenge of 25,2568 EU/ml. After one hour, the solutions were centrifuged and the supernatants tested for LPS using Endo-LISA test kits.
The results show that 1 mg of SOLIS MAX adsorbs 20 endotoxin units (EU) of E. coli endotoxin, which corresponds – for this challenge – to an 80% adsorption rate (figure 3).
Figure 3: SOLIS MAX effectively adsorbs E. coli endotoxins
Endotoxin solution SOLIS MAX: Stabilize gut health, support performance
The detrimental impact of LPS can be mitigated by using a high-performance solution such as SOLIS MAX. To prevent negative health and performance outcomes for the animal it is important to stabilize the challenged intestinal barrier and to support the balance of the gut microbiome. Binding endotoxins before they can exert their damaging impact is the primary objective, which SOLIS MAX achieves through the intelligent interaction of natural plant extracts. This can be expected to yield positive results in terms of production levels and the prevention of secondary infections, preserving animal health and farms’ economic viability.
References
Adib-Conquy, Minou, and Jean-Marc Cavaillon. “Compensatory Anti-Inflammatory Response Syndrome.” Thrombosis and Haemostasis 101, no. 01 (2009): 36–47. https://doi.org/10.1160/th08-07-0421.
Huntley, Nichole F., C. Martin Nyachoti, and John F. Patience. “Immune System Stimulation Increases Nursery Pig Maintenance Energy Requirements.” Iowa State University Animal Industry Report 14, no. 1 (2017). https://doi.org/10.31274/ans_air-180814-344.
Li, Jiaolong, Yongqing Hou, Dan Yi, Jun Zhang, Lei Wang, Hongyi Qiu, Binying Ding, and Joshua Gong. “Effects of Tributyrin on Intestinal Energy Status, Antioxidative Capacity and Immune Response to Lipopolysaccharide Challenge in Broilers.” Asian-Australasian Journal of Animal Sciences 28, no. 12 (2015): 1784–93. https://doi.org/10.5713/ajas.15.0286.
Efficient microbiome modulation with phytomolecules
By Dr. Inge Heinzl, Editor, EW Nutrition
From day 1, young animals are confronted with the pathogens of their environment. Feed and feed ingredients also significantly increase exposure to microbes. This article will look closely at three critical bacteria in poultry production. The trials of phytomolecules-based products shared in this article prove the unique benefit of lowering harmful pathogens while simultaneously sparing health-promoting microbes. The targeted selection of the blend’s phytomolecules contributes to this distinctive mode of action.
E. coli can be valuable… and dangerous
E.coli are commensal bacteria that usually belong to the natural gut flora. However, there are several E. coli strains that, due to certain virulence factors, can cause disease. These bacteria are called avian pathogenic E. coli or APEC. The disease ‘Colibacillosis’ can occur in different forms:
Omphalitis – a noncontagious infection of the navel and/or yolk sac in young poultry
peritonitis – inflammatory response on “internal laying” (yolk material in the peritoneum)
salpingitis – inflammation of the oviduct
cellulitis – discoloration and thickening of the skin, inflammation of the subcutaneous tissues
synovitis – lameness with swollen joints
coligranuloma (Hjärre disease) – lesions similar to tuberculosis, not of economic importance
meningitis, and
septicemia or blood poisoning.
Since some of the E. coli strains can sometimes be transmitted vertically to offspring, it is crucial to keep the pathogenic pressure in the parent generation as low as possible (Mc Dougal, 2018).
Due to the, mostly in young chicks, common use of antibiotics, E. coli strains resistant to ß-lactam antibiotics (ESBL-producing E. coli) or fluoroquinolones (e.g., Enrofloxacin) have developed.
Clostridium perfringens: the cause of necrotic enteritis
Clostridium perfringens belong to the normal caecal flora. However, its overgrowth in the intestine is linked to necrotic enteritis, causing estimated losses of up to USD 6 billion yearly in global poultry production, which corresponds to USD 0.0625 per bird (Wade and Keyburn, 2015). Necrotic enteritis can occur in a clinical and a subclinical form.
In the case of clinical necrotic enteritis, the birds suffer from diarrhea resulting in wet litter and increased flock mortality of up to 1 % per day (Ducatelle and Van Immerseel, 2010). Mortality rates sometimes sum up to 50 % (Van der Sluis, 2013). If birds die without clinical signs, it may be peracute necrotic enteritis.
The subclinical version, however, is more critical. Due to the lack of symptoms, it often remains undetected and, therefore, not treated. Mainly through the impaired utilization of feed, representing 65-75 % of the total costs in broiler production, subclinical necrotic enteritis permanently impacts production efficiency (Heinzl et al., 2020).
Salmonella enterica: a zoonosis relevant for birds and humans
Most concerning in (non-typhoid) salmonellosis is that it can be transferred to humans. The transmission occurs via direct contact with an infected animal, consuming contaminated animal products such as meat or eggs, contact with infected vectors (insects or pets) or contaminated equipment, or cross-contamination in the kitchen. Frozen or raw chicken products, as well as the eggs, are frequent causes of animal-origin Salmonella infections in humans.
Salmonella is the more critical the younger the birds. If the hatching eggs already carry salmonellae, the hatchability dwindles. During their first weeks of life, infected chicks show higher mortality and systemic infections.
Adult animals usually do not die from salmonellosis; often, the infection remains unnoticed. During an acute salmonella outbreak, the animals might show weakness and diarrhea. They lose weight, resulting in decreased egg production in layers.
Trials with phytomolecules show promising results
To check if phytomolecules-based products can effectively influence gut flora, a product specially designed for gut health (Ventar D) was tested for its antimicrobial activity. Additionally, the extent to which the same blend impacted the beneficial bacteria, such as Lactobacilli, was evaluated.
Trial 1: phytomolecules act against E. coli and Salmonella enterica
The in vitro study using the agar dilution method was conducted at a German laboratory.
The bacteria (Salmonella typhimurium and ESBL-producing E. coli) stored at -80°C were reactivated by cultivating them on Agar Mueller Hinton overnight. After this incubation, some colonies were picked and suspended in 1 ml 0.9% NaCl solution. 100 µl of the suspension were pipetted and evenly spread (plate spread technique) on new Agar Mueller Hinton containing different concentrations of a phytomolecules-based product (Ventar D): 0 µg/mL – control; 500 µg/mL; 900 µg/mL; 1.250 µg/mL and 2.500 µg/mL. After 16-20 h incubation at 37°C, growth was evaluated. The results can be seen in pictures 1 and 2:
Figure 1: E. coli exposed to different concentrations of Ventar D (upper row from left to right: control 0 µg/ml, 500 µg/ml, 900 µg/ml; lower row from left to right: 1250 µg/ml and 2500 µg/ml)
E. coli colonies exposed to 900 µg/mL of Ventar D’s phytogenic formulation were smaller than the control colonies. At 1250 µg/mL, fewer colonies were detected, and at 2500 µg/mL, growth couldn’t be seen anymore.
The salmonella colonies showed a similar picture; however, the reduction could be seen from a concentration of 1.250 µg/ml of Ventar D onwards (picture 2).
Figure 2: Salmonella enterica exposed to different concentrations of Ventar D (upper row from left to right: control 0 µg/ml, 500 µg/ml, 900 µg/ml; lower row from left to right: 1250 µg/ml and 2500 µg/ml)
Trial 2: Phytomolecules inhibit Clostridium perfringens and spare Lactobacilli
In this trial, the bacteria (Clostridium perfringens,Lactobacillus agilis S73, and Lactobacillus plantarum) were cultured under favorable conditions (RCM, 37°C, anaerobe for Clostr. perfr., and MRS, 37°C, 5 % CO2 for Lactobacilli) and exposed to different concentrations of Ventar D (0 µg/ml – control, 500 µg/ml, 750 µg/ml, and 1000 µg/ml).
The results are shown in figures 3a-d.
Figure 3a: control, 0 µg/ml
Figure 3b: 500 µg/ml
Figure 3c: 750 µg/ml
Figure 3d: 1000 µg/m
In the case of Clostridium perfringens, a significant reduction of colonies could already be observed at a concentration of 500 µg/ml of Ventar D. At 750 µg/ml, only a few colonies remained. At a Ventar D concentration of 1000 µg/ml, Clostridium perfringens could no longer grow.
In contrast to Clostridium, the Lactobacilli showed a different picture: only at the higher concentration (1250 µg/ml of Ventar D), Lactobacillus plantarum and Lactobacillus agilis S73 showed a slight growth reduction (figures 4 and 5).
Figure 4: Lactobacillus plantarum exposed to 0 (left) and 1250 µg/ml (right) of Ventar D
Figure 5: Lactobacillus agilis S73 exposed to 0 (left) and 1250 µg/ml (right) of Ventar D
Improve gut health by positively influencing the intestinal flora
The experiments show that even at lower concentrations, phytomolecules impair the growth of harmful bacteria while sparing the beneficial ones. Phytomolecule-based products can be regarded as a valuable tool for controlling relevant pathogens in poultry and influencing the microflora composition in a positive way.
The resulting better gut health is the best precondition to reducing antibiotics in animal production.
Global mycotoxin report: Jan-June 2022 | Find the pain points
By Technical Team, EW Nutrition
The pressure of climate change is taking a severe toll – not just on weather-dependent industries, but already on society in general. For feed and food, the impact is already dramatic. Extreme weather events, increased temperatures, and rising carbon dioxide levels are facilitating the growth of toxigenic fungi in crops, severely increasing the risk of mycotoxin contamination. Once feed is contaminated, animal health can be impacted, with chain reactions affecting productivity for animal farming, as well as, ultimately, the quality and availability of food.
*** Download the full report for an analysis of mycotoxin contamination risks around the world
Acidifiers support piglets after weaning
By Dr. Inge Heinzl, Editor, EW Nutrition
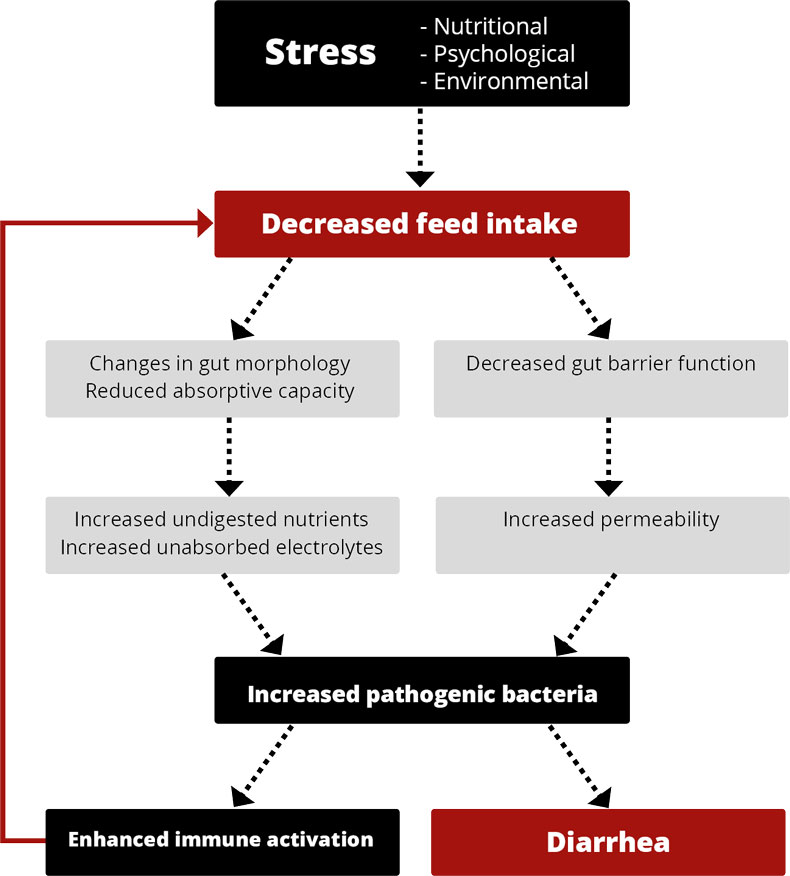
In piglet production, high productivity, meaning high numbers of healthy and well-performing piglets weaned per sow and year, is the primary target. Diarrhea around weaning often gets in the way of achieving this goal.
Up to the ban of antibiotic growth promoters in 2006, antibiotics were often applied prophylactically to help piglets overcome this critical time. Zinc oxide (ZnO) application is another measure that cannot be used anymore to prevent piglet diarrhea. Effective alternatives are required.
Weaning – a critical point in piglets’ life
Weaning stress is well-known to have a negative impact on the balance of the intestinal microflora and gastrointestinal functions (Miller et al., 1985). Suckling piglets have a limited ability to produce hydrochloric acid, but nature has a solution to compensate for this inadequacy. The lactobacilli present in the stomach can use the lactose in the sow’s milk to produce lactic acid (Easter, 1988). In nature, the piglets would start to eat small amounts of solid feed at about three weeks when the sow’s milk production no longer covers their nutrient demand. By increasing the feed intake, the piglets stimulate hydrogen chloride (HCl) production in their stomachs.
In piglet production, where weaning occurs at three or four weeks of age, the piglets are still not eating considerable amounts of solid feed. It is often the case that 50 % of the piglets take feed at the earliest after 24 h, and 10 % accept the first feed only after 48 h (Brooks, 2001). Additionally, hard grains in the diet can physically damage the small intestine wall, reducing villus height and crypt depth (Kim et al., 2005).
Only a minor production of HCl, no more lactose supply for the lactobacilli, varying feed intake, and high buffering capacity of the feed lead to a pH of >5 in the stomach.
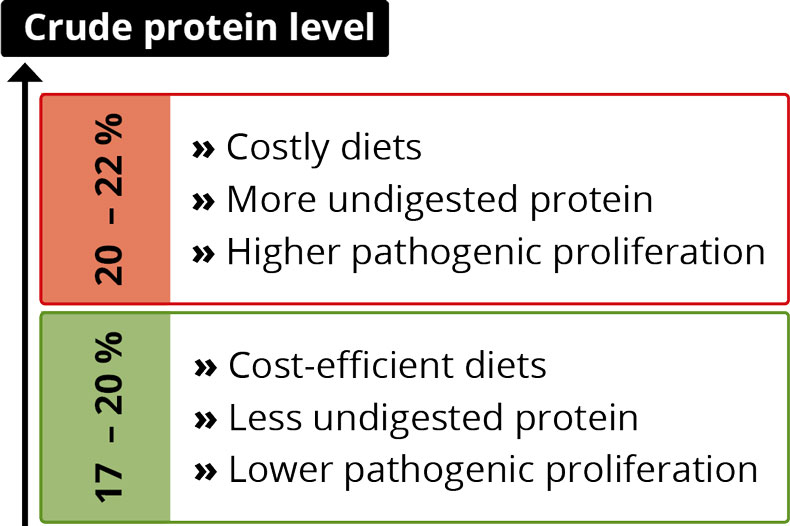
The higher stomach pH is partly responsible for problems after weaning
A pH higher than 5, besides causing direct effects on the microflora in the stomach, has consequences for the whole digestive tract and digestion.
A high pH is favorable for certain microorganisms, including coliforms (Sissons, 1989) and other acid-sensitive bacteria such as Salmonella typhimurium, Salmonella typhi, Campylobacter jejuni, and V. cholerae (Smith, 2003).
Lower activity of proteolytic enzymes
In the stomach, the conversion of pepsinogen to pepsin, which is responsible for protein digestion, is catalyzed under acid conditions (Sanny et al., 1975). Pepsin works optimally at two pH levels: pH 2 and pH 3.5 (Taylor, 1959). With increasing pH, the activity decreases; at pH 6, it stops. Therefore, a high pH can lead to poor digestion and undigested protein arriving in the intestine. There, it can be used as “feed” for harmful bacteria, leading to their proliferation. Barrow et al. (1977) found higher counts of coliforms in piglets’ intestinal tract two days after weaning, while the number of lactobacilli was depressed.
In the lower parts of the gastrointestinal tract (GIT), the final products of the pepsin protein digestion are needed to stimulate the secretion of pancreatic proteolytic enzymes. If no final products arrive, the enzymes are not activated, and the inadequate protein digestion continues. Additionally, gastric acid is the main stimulant for bicarbonate secretion in the pancreas, neutralizing gastric acid and providing an optimal pH environment for the digestive enzymes working in the duodenum.
Expedited digesta transport
The stomach pH also influences the transport of digesta. The acidity of the chyme leaving the stomach and arriving in the small intestine is decisive for the amount of digesta being transferred from the stomach to the small intestine. Acid-sensitive receptors provide feedback regulation to prevent the stomach from emptying until the duodenal chyme can be neutralized by pancreatic or other secretions (Pohl et al., 2008). Therefore, a higher pH in the stomach leads to a faster transport of the digesta, resulting in worse feed digestion.
Proliferation of microorganisms
A high pH is favorable for certain microorganisms, including coliforms (Sissons, 1989) and other acid-sensitive bacteria such as Salmonella typhimurium, Salmonella typhi, Campylobacter jejuni, and V. cholerae (Smith, 2003).
Elevated stomach pH + incomplete immune system = diarrhea
Acidifiers can mitigate the adverse effects of weaning on piglets
To overcome this critical time of weaning and maintain performance, acidifiers can be a helpful tool. They improve gut health, stimulate immunity, and serve as nutrient sources – while also positively affecting feed and water hygiene.
What are acidifiers?
Acidifiers’ role in pig nutrition has evolved from feed preservatives to stomach pH stabilizers, compensating for young pigs’ reduced digestive capacity (Ferronato and Prandini, 2020). They are now used to replace antibiotic growth promoters and ZnO, which were applied for a long time to mitigate the negative effects of weaning.
In general, both organic and inorganic acids and their salts feature in animal nutrition. They can be added to the feed or the water.
Organic acids: Commonly used with good results
Feed acidifiers are usually organic acids, including fatty and amino acids. Their carboxyl functional group is responsible for their acidic specificity as feed additives (Pearlin et al.,2019). Their pKa, the pH where 50 % of the acid occurs in a dissociated form, is decisive for their antimicrobial action. In animal nutrition, acids with pKa 3-5 are typically used (Kirchgeßner and Roth, 1991).
Organic acids used as feed additives can be divided into three groups:
Simple monocarboxylic acids such as formic, acetic, propionic, and butyric acid
Carboxylic acids with a hydroxyl group such as lactic, malic, tartaric, and citric acid
Short-chain carboxylic acids with double bonds – fumaric and sorbic acid.
The primary acids for pig nutrition are acetic, fumaric, formic, lactic, benzoic, propionic, sorbic, and citric acids (Roth and Ettle, 2005).
Inorganic acids – the low-cost version
Inorganic acids are cheaper than organic acids, but their only effect is to decrease the pH. Additionally, they are extremely corrosive and dangerous liquids due to their strong acidity in a pure state (Kim et al., 2005).
Salts are easier to handle
The advantage of salts over free acids is that they are generally odorless and easier to handle in the feed manufacturing process due to their solid and less volatile form. Higher solubility in water is a further advantage compared to free acids (Huyghebaert and Van Immerseel, 2011; Roth and Ettle, 2005; Partanen and Mroz, 1999). The better handling and higher palatability make acid salts a more user-friendly method to apply acids to feed and water without compromising their efficacy (Luise et al., 2020).
The salts are mainly produced with calcium, potassium, and sodium. They include calcium formate, potassium diformate, sodium diformate, and sodium fumarate.
Blends
A mixture of diverse acidifiers combines the different characteristics of these substances. Perhaps, there may be synergistic effects. Acid blends are more and more used as feed additives. They have a wider-ranging action than single substances.
Roth et al. (1996) showed that a combination of formic acid with various formats is more effective than the application of formic acid alone.
The main effects of acidifiers
Acidifiers support piglets during the critical time after weaning through different modes of action. The final results are:
Improvement in gut health
Increase in growth performance
Stabilization of the immune system.
1. Improvement in gut health
As shown in figure 1, the improvement in gut health relies on the antimicrobial effect of organic acids and the decrease in the stomach’s pH.
1.1 Organic acids directly attack bacteria
Organic acids not only act through their pH-decreasing effect but also directly attack pathogens. Due to their lipophilic character, organic acids can pass the bacterial cell membrane when they are in their undissociated form (Partanen in Piva et al., 2001). The lower the external pH, the more undissociated acid can pass the membrane.
Within the cell, the pH is higher. Hence, the organic acid dissociates and releases hydrogen ions, reducing the cytoplasmic pH from alkaline to acid. Cell metabolism is depressed at lower pH. Therefore, the bacterial cell needs to expel protons to get the cytoplasmic pH back to normal. As this is an energy-consuming process, more prolonged exposure to organic acids kills the bacterium. Additionally, the anions staying within the cell disturb the cell’s metabolic processes and participate in killing the bacterium.
Studies from Van Immerseel et al. (2006) revealed that many fermentative bacteria could let their intracellular pH decline and prevent increased acid penetration. Bacteria with a neutrophil pH, however, react more sensitively.
There is a direct effect of pH on the microflora. Some pathogenic bacteria are susceptible to low pH. The proliferation of, e.g., E. coli, Salmonella, and Clostridium perfringens is minimized at a pH<5. Acid-tolerant bacteria such as lactobacilli or bifidobacteria, however, survive. Many lactobacilli can produce hydrogen peroxide, which inhibits, e.g., Staphylococcus aureus or Pseudomonas spp. (Juven and Pierson, 1996).
Already Fuller (1977) showed in in vitro experiments that certain bacteria such as Streptococci, Salmonella, and B. cereus don’t grow in an environment with pH 4.5 or even die (Micrococcus). In contrast, Lactobacilli are not so susceptible to this low pH. Using the same binding sites as harmful bacteria, they suppress coliforms, for example. Kirchgeßner et al. (1997) found a stronger reduction of E. coli than Lactobacilli and Bifidobacteria in different gut segments when exposed to 1.25 % formic acid.
1.2 Recovery of eubiosis through reduction of substrate
The reduction of the pH through organic acids maintains or stimulates the secretion of proteolytic enzymes in the stomach (pepsin) and pancreatic enzymes. Additionally, the acid leaving the stomach is partly responsible for regulating gastric emptying (Ravindran and Kornegay, 1993;Mayer, 1994). Both effects by improving protein digestion, reduce the fermentable substrates arriving in the hindgut. This decreases the quantity of fermentable substrate arriving in the intestine and, therefore, the growth of undesired pathogens.
2. Promotion of growth
2.1 Enhanced digestion of macronutrients
As explained above, the acidity in the stomach is responsible for the activation and stimulation of enzymes. Additionally, the lower pH keeps the feed in the stomach for longer. Both result in better digestion.
The improved utilization of nutrients leads to higher daily gain and better feed conversion. In pigs, the growth-promoting effect of organic acids is particularly pronounced during the first few weeks after weaning (Roth and Ettle, 2005). Some examples of the growth-promoting effect of formic and propionic acid feature in table 1.
Table 1: Influence of two commonly used organic acids in animals on growth performance
Varying results are mainly due to the character of the organic acid, the dosage, the buffering capacity, and the possible reduction of feed intake in case of a high dosage (Roth and Ettle, 2005).
2.2 Improved utilization of minerals
Minerals are essential for metabolic processes and, thus, healthy growth. Chelated minerals show a higher digestibility. Acidic anions of the acidifiers form complexes (chelates) with cationic minerals such as Ca, Zn, P, and Mg. The resulting higher digestibility and absorption lead to decreased excretion of supplemented minerals and, therefore, to a lower environmental burden. Kirchgeßner and Roth (1982), e.g., reported an improved absorption and retention of Ca, P, and Zn with the addition of fumaric acid. However, there are also trials showing no effect of acidification of the diet on mineral balance (Radecki et al., 1988).
Phytic acid
Another factor influencing the absorption of minerals, mainly phosphorus, is the amount of intrinsic or microbial phytase in the diet (Rutherfurd et al., 2012). The enzyme phytase releases phosphorus out of phytic acid and increases its bioavailability. Partanen and Mroz(1999) showed that organic acids improve the performance of phytase and, therefore, the bioavailability of phosphorus in the diet.
Besides a better utilization by the animal, improved absorption of minerals means preserving the environment and direct cost-saving, as mineral supplements are expensive.
2.3 Stimulation of gut and stomach mucosal morphology
An intact gut mucosa with a preferably high surface is vital for efficient nutrient absorption. Many trials show that organic acids improve the condition of the mucosa:
Organic acids stimulate cell proliferation
In an in vitro trial with pig hindgut mucosa, butyric acid stimulated epithelial cell proliferation in a dose-dependent manner (Sakata et al., 1995).
Blank et al. reported that fumaric acid, being a readily available energy source, may have a local trophic effect on the small intestines’ mucosa. Due to faster recovery of the gastrointestinal epithelial cells after weaning, this trophic effect may increase the absorptive surface and digestive capacity in the small intestines.
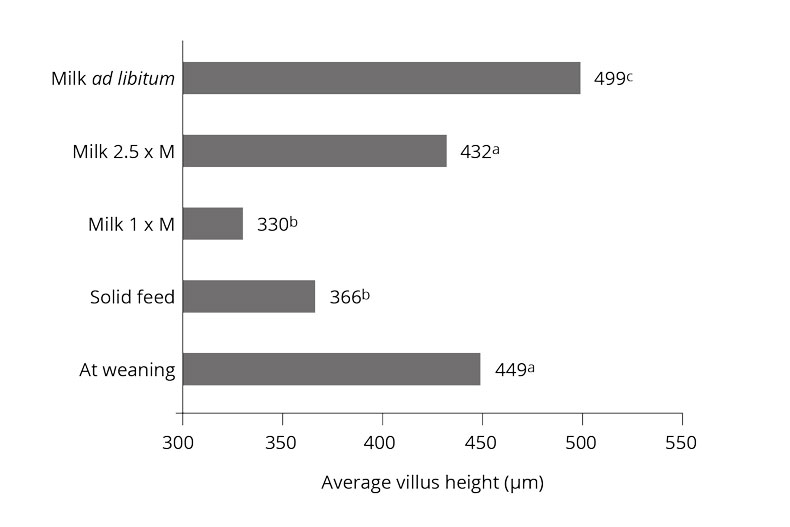
Organic acids influence villi length and crypt depth in the gut
Ferrara et al. (2016) observed a trend toward longer villi with a mixture of short-chain organic acids and mid-chain fatty acids, compared to the negative control.
The addition of Na-butyrate to the feed leads to increased crypt depth, villi length, and mucosa thickness in the distal jejunum and ileum, according to Kotunia et al. (2004). However, the villi length and mucosa thickness were reduced in the duodenum.
According to Gálfi and Bokori (1990), a diet with 0.17% sodium butyrate increased the length of ileal microvilli and the depth of caecal crypts in pigs weighing between 7 and 102 kg.
Organic acids strengthen stomach mucosa
Mazzoni et al. (2008) reported that sodium butyrate applied orally at a low dose influenced gastric morphology and function (thickening the mucosa), presumably due to its action on mucosal maturation and differentiation.
2.4 Pigs can use organic acids as an energy source
Organic acids are usually added to the feed in small doses. As some organic acids are intermediary products of the citric acid cycle, they are an energy source after being absorbed through the gut epithelium by passive diffusion. Their gross energy can be fully metabolized (Pearlin et al., 2019; Roth and Ettle, 2005; Suiryanrayna and Ramana, 2015).
The gross energy supply varies according to the acid. Roth and Ettle (2005) determined values between 6 kJ/g (formic acid) and 27 kJ/g (sorbic acid). Pearlin et al. (2019) calculated that 1 M of fumaric acid generates 1.340 kJ or 18 M ATP; this is comparable to the energy provision of glucose. Citric acid’s energy provision is similar; acetic and propionic acid require 18 and 15 % more energy to generate 1 M ATP.
Acidifiers improve immune response
The immune system, especially at the sensitive life stage of weaning, plays an essential role for the piglet. Acidifiers have been shown to stimulate or support the immune system. Ahmed et al. (2014) showed that citric acid (0.5 %) and a blend of acidifiers composed of formic, propionic, lactic, phosphoric acid + SO2(0.4 %) significantly increased the level of serum IgG. IgG are part of the humoral immune system. They mark foreign substances to be eliminated by other defense systems.
In a trial conducted by Ren et al. (2019), piglets receiving a mixture of formic and propionic acid showed lower concentrations of plasma tumor necrosis factor-α, regulating the activity of diverse immune cells. Furthermore, interferon-γ and interleukin (Il)-1ß were lower than in the control group after the challenge with E. coli (ETEC). In this trial, the addition of organic acids to the feed alleviated the inflammatory response in a way comparable to antibiotics.
In a nutshell
Organic acids are no longer seen as pure acidifiers but as growth promoters and potential antibiotic substitutes due to their positive effect on the gastrointestinal tract. Their main effect, the decrease of pH, entails consequences from inhibiting pathogenic bacteria and improved digestion to enhanced health and growth.
Research indicates that acidifiers can be a viable alternative to antibiotic growth promoters and ZnO for ensuring healthy piglet production after weaning.
Price hikes = more cereal byproducts in animal feed. What about mycotoxin risk?
By Technical Team, EW Nutrition
Most grains used in feed are susceptible to mycotoxin contamination, causing severe economic losses all along feed value chains. As skyrocketing raw material prices force producers to include a higher proportion of economical cereal byproducts in the feed, the risks of mycotoxin contamination likely increase. In this article, we review why mycotoxins cause the damage they do – and how effective toxin-mitigating solutions prevent this damage.
Mycotoxin contamination of cereal byproducts requires solutions
Cereal byproducts may become more important feed ingredients as grain prices increase. But also from a sustainability point of view and considering population growth, using cereal byproducts in animal feed makes a lot of sense. Dried distiller’s grains with solubles (DDGS) are a good example of how byproducts from food processing industries can become high-quality animal feed.
Figure 1: Byproducts are a crucial protein source (data from FEFAC Feed & Food 2021 report)
Still, research on what happens to mycotoxins during food processing shows that mycotoxins are concentrated into fractions that are commonly used as animal feed (cf. Pinotti et al., 2016 + link to article IH+MC ). To safeguard animal health and performance when feeding lower-quality cereals, it is essential to monitor mycotoxin risks through regular testing and to use toxin-mitigating solutions.
Problematic effects of mycotoxins on the intestinal epithelium
Most mycotoxins are absorbed in the proximal part of the gastrointestinal tract. This absorption can be high, as in the case of aflatoxins (ca. 90%), but also very limited, as in the case of fumonisins (< 1%); moreover, it depends on the species. Importantly, a significant portion of unabsorbed toxins remains within the lumen of the gastrointestinal tract.
Importantly, studies based on realistic mycotoxin challenges (e.g., Burel et al., 2013) show that the mycotoxin levels necessary to trigger damaging processes are lower than the levels reported as safe by EFSA, the Food Safety Agency of the European Union. The ultimate consequences range from diminished nutrient absorption to inflammatory responses and pathogenic disorders in the animal (Figure 2).
Figure 2: Mycotoxins’ impact on the GIT and consequences for monogastric animals
Alteration of the intestinal barrier‘s morphology and functionality
Several studies indicate that mycotoxins such as aflatoxin B1, DON, fumonisin B1, ochratoxin A, and T2, can increase the permeability of the intestinal epithelium of poultry and swine (e.g. Pinton & Oswald, 2014). This is mostly a consequence of the inhibition of protein synthesis.
As a result, there is an increase in the passage of antigens into the bloodstream (e.g., bacteria, viruses, and toxins). This increases the animal’s susceptibility to infectious enteric diseases. Moreover, the damage that mycotoxins cause to the intestinal barrier entails that they are also being absorbed at a higher rate.
Impaired immune function in the intestine
The intestine is a very active immune site, where several immuno-regulatory mechanisms simultaneously defend the body from harmful agents. Immune cells are affected by mycotoxins through the initiation of apoptosis, the inhibition or stimulation of cytokines, and the induction of oxidative stress.
For poultry production, one of the most severe enteric problems of bacterial origin is necrotic enteritis, which is caused by Clostridium perfringens toxins. Any agent capable of disrupting the gastrointestinal epithelium – e.g. mycotoxins such as DON, T2, and ochratoxin – promotes the development of necrotic enteritis.
It is known that fumonisin B1 may induce changes in the balance of sphingolipids at the cellular level, including for gastrointestinal cells. This facilitates the adhesion of pathogenic bacteria, increases in their populations, and prolongs infections, as has been shown for the case of E. coli. The colonization of the intestine of food-producing animals by pathogenic strains of E. coli and Salmonella also poses a risk for human health.
Interaction with bacterial toxins
When mycotoxins induce changes in the intestinal microbiota, this can lead to an increase in the endotoxin concentration in the intestinal lumen. Endotoxins promote the release of several cytokines that induce an enhanced immune response, causing inflammation, thus reducing feed consumption and animal performance, damage to vital organs, sepsis, and death of the animals in some cases.
The synergy between mycotoxins and endotoxins can result in an overstimulation of the immune system. The interaction between endotoxins and estrogenic agents such as zearalenone, for example, generates chronic inflammation and autoimmune disorders because immune cells have estrogen receptors, which are stimulated by the mycotoxin.
Increased mycotoxin risks through byproducts? Invest in mitigation solutions
To prevent the detrimental consequences of mycotoxins on animal health and performance, proactive solutions are needed that support the intestinal epithelium’s digestive and immune functionality and help maintain a balanced microbiome in the GIT. As the current market conditions will likely engender a long-term shift towards the inclusion of more cereal byproducts in animal diets, this becomes even more important.
Trial data shows that EW Nutrition’s toxin-mitigating solution SOLIS MAX provides effective protection against feedborne mycotoxins. The synergistic combination of ingredients in SOLIS MAX mycotoxins from damaging the animals’ gastrointestinal tract and entering the blood stream:
In-vitro study shows SOLIS MAX’ strong mitigation effects against wide range of mycotoxins
Animal feed is often contaminated with two or more mycotoxins, making it important for an anti-mycotoxin agent to be effective against a wide range of different mycotoxins. A dose response evaluation of SOLIS MAX was conducted a at an independent laboratory in Spain, for inclusion levels of 0.10%, 0.15%, and 0.20% (equivalent to 1 kg, 1.5 kb, and 2 kg per ton of feed). A phosphate buffer solution at pH 7 was prepared to simulate intestinal conditions in which a portion of the mycotoxins may be released from the binder (desorption).
Each mycotoxin was tested separately by adding a challenge to buffer solutions, incubating for one hour at 41°C, to establish the base line (see table). At the same time a solution with the toxin challenge and SOLIS MAX was prepared, incubated, and analyzed for the residual mycotoxin. All analyses were carried out by high performance liquid chromatography (HPLC) with standard detectors.
Figure 3: SOLIS MAX adsorption capacity against different mycotoxins (%)
The results demonstrate that SOLIS MAX is a very effective solution against the most common mycotoxins found in raw materials and animal feed, showing clear dose-response effects.
Mycotoxin risk management for better animal feed
A healthy gastrointestinal tract is crucial to animals’ overall health: it ensures that nutrients are optimally absorbed, it provides effective protection against pathogens through its immune function, and it is key to maintaining a well-balanced microflora. Even at levels considered safe by the European Union, mycotoxins can compromise different intestinal functions, resulting in lower productivity and susceptibility to disease.
The globalized feed trade, which spreads mycotoxins beyond their geographical origin, climate change and raw material market pressures only escalates the problem. On top of rigorous testing, producers should mitigate unavoidable mycotoxin exposures through the use of solutions such as SOLIS MAX – for stronger animal health, welfare, and productivity.
References
Antonissen, Gunther, An Martel, Frank Pasmans, Richard Ducatelle, Elin Verbrugghe, Virginie Vandenbroucke, Shaoji Li, Freddy Haesebrouck, Filip Van Immerseel, and Siska Croubels. “The Impact of Fusarium Mycotoxins on Human and Animal Host Susceptibility to Infectious Diseases.” Toxins 6, no. 2 (January 28, 2014): 430–52. https://doi.org/10.3390/toxins6020430.
Burel, Christine, Mael Tanguy, Philippe Guerre, Eric Boilletot, Roland Cariolet, Marilyne Queguiner, Gilbert Postollec, et al. “Effect of Low Dose of Fumonisins on Pig Health: Immune Status, Intestinal Microbiota and Sensitivity to Salmonella.” Toxins 5, no. 4 (April 23, 2013): 841–64. https://doi.org/10.3390/toxins5040841.
Burton, Emily J., Dawn V. Scholey, and Peter E. Williams. “Use of Cereal Crops for Food and Fuel – Characterization of a Novel Bioethanol Coproduct for Use in Meat Poultry Diets.” Food and Energy Security 2, no. 3 (September 19, 2013): 197–206. https://doi.org/10.1002/fes3.30.
Ghareeb, Khaled, Wageha A. Awad, Josef Böhm, and Qendrim Zebeli. “Impacts of the Feed Contaminant Deoxynivalenol on the Intestine of Monogastric Animals: Poultry and Swine.” Journal of Applied Toxicology 35, no. 4 (October 28, 2014): 327–37. https://doi.org/10.1002/jat.3083.
Mani, V., T. E. Weber, L. H. Baumgard, and N. K. Gabler. “Growth and Development Symposium: Endotoxin, Inflammation, and Intestinal Function in livestock1,2.” Journal of Animal Science 90, no. 5 (May 1, 2012): 1452–65. https://doi.org/10.2527/jas.2011-4627.
Obremski, K. “The Effect of in Vivo Exposure to Zearalenone on Cytokine Secretion by Th1 and Th2 Lymphocytes in Porcine Peyer’s Patches after in Vitro Stimulation with LPS.” Polish Journal of Veterinary Sciences 17, no. 4 (2014): 625–32. https://doi.org/10.2478/pjvs-2014-0093.
Oswald, I. P., C. Desautels, J. Laffitte, S. Fournout, S. Y. Peres, M. Odin, P. Le Bars, J. Le Bars, and J. M. Fairbrother. “Mycotoxin Fumonisin B1 Increases Intestinal Colonization by Pathogenic Escherichia Coli in Pigs.” Applied and Environmental Microbiology 69, no. 10 (2003): 5870–74. https://doi.org/10.1128/aem.69.10.5870-5874.2003.
Pinotti, Luciano, Matteo Ottoboni, Carlotta Giromini, Vittorio Dell’Orto, and Federica Cheli. “Mycotoxin Contamination in the EU Feed Supply Chain: A Focus on Cereal Byproducts.” Toxins 8, no. 2 (February 15, 2016): 45. https://doi.org/10.3390/toxins8020045.
Pinton, Philippe, and Isabelle Oswald. “Effect of Deoxynivalenol and Other Type B Trichothecenes on the Intestine: A Review.” Toxins 6, no. 5 (May 21, 2014): 1615–43. https://doi.org/10.3390/toxins6051615.
The risk of using by-products as feed alternatives – and the solutions
By Technical Team and Dr. Inge Heinzl, Editor, EW Nutrition
Consistently rising feed prices compel feed producers to resort to alternative feed ingredients. By-products of milling and ethanol distillation would be good options. The following article shows what should be paid attention to when using these feeds.
Keeping high-quality animal protein affordable requires cost-efficient alternatives
For a high percentage of consumers, the price of food products is one of the most decisive purchase factors; however, quality and sustainable use of resources are also of high importance. So, to comply with market requirements, meat producers must find cost-efficient and sustainable sources of feed ingredients. Feed prices already increased during the COVID-19 pandemic. Shortage of workforce and high shipping costs led to discontinuity in the supply chain, long delivery times, and increased costs for certain raw materials. Due to the Ukrainian crisis, there is no improvement to be seen. Alternatives must be considered more vigorously to compensate for this limited feed supply.
Grain by-products are an option
The use of grain by-products occurring at milling or ethanol production can cover a part of animal nutritional demands. Additionally, it contributes to sustainable usage of the available sources, as the remains of the production of human consumables are put back into the food chain.
However, increasing levels of by-products in the feed also have their sticking points. The raw materials grains or corn are often contaminated with mycotoxins, impacting the quality of this kind of feed.
Milling processes reduce mycotoxins in food
Before the whole process of milling, the grains are sorted and cleaned. Kernels with extensive mold growth, broken kernels, fine materials, and dust are removed.
When it comes to reducing mycotoxins by sorting and cleaning, the results vary a lot. They are influenced by several factors, including the initial condition of the grains, the type and level of contamination, and the type and efficiency of the cleaning process (Pinotti et al., 2016). The cleaning process has been shown to remove from 5 to 80 % of DON and NIV, 5 to 40 % of ZON (Schaarschmidt & Fauhl-Hassek, 2018), and 50 to 60 % of T2/HT2 contamination in wheat (Pascale et al., 2011). Debranning, the mechanical process by which the outer layers of wheat grains are removed, further reduces mycotoxin content in wheat grain from 15 to 80% of the initial contamination (Aureli et al., 2007; Rios et al., 2009). However, neither the cleaning and debranning nor the milling process include a step that destroys mycotoxins.
In white flour for human consumption, mycotoxin levels typically range from 50 to 70% of the wheat grain (Cheli et al., 2013).
The milling of maize shows a reduction factor of about 4 for aflatoxins and about 10 for zearalenone from the grain to the final human products. Contrarily, concentration triplicates for both aflatoxins and zearalenone in the case of the by-products such as germs, bran, and animal flour.
Milling processes concentrate mycotoxins in animal-feed fractions
The milling and pre-milling processes reduce the content of mycotoxins in products for human consumption, but what about the parts removed and normally used in animal feeds? Several studies (Tibola et al., 2015; Hoffmans et al., 2022) indicate that the concentration of mycotoxins is higher in the wheat fractions intended for animal feeds such as bran, flour shorts screenings, and middlings. However, their level in feedstuffs is variable and affected by several factors such as the type of mycotoxins, the level and extent of fungal contamination, and the complexity of the cereal processing technology.
Compared to the concentration in wheat grain, these concentrations in by-products may be up to 800 % but more typically range from 150 % to 340 % (Cheli et al., 2013). EW Nutrition’s worldwide mycotoxin survey shows a similar trend (Figures 1 and 2), in which DON levels are nine times higher in wheat midds than wheat grains, and fumonisin is eight times higher in wheat bran.
Figure 1 + 2: Mycotoxins levels in grain and by-products
Highest concentrations in germ and bran fraction
After corn milling, animal feed fractions such as germ and bran have a low yield ranging from 5 to 7 % and are mostly composed of the outer parts of the kernels; as a consequence, an important concentration of mycotoxins occurs in these fractions (Schollenberger et al., 2008). When taking corn grains as the base, the contamination of aflatoxins goes up to three times in corn germ and up to nine times in bran (Brera et al., 2006; Pietri et al., 2009). For fumonisins, a double concentration can be expected (Brera et al., 2004), and for zearalenone, up to four times (Brera et al., 2006). Recently, Park and collaborators (2018) evaluated the distribution of 12 mycotoxins during wet milling of corn and found higher concentrations in corn gluten feed and corn bran.
Milling is a crucial step in the post-production of rice, in which the husk and the bran layers are removed. Rice bran is a common ingredient for animal feeds, in which aflatoxin is a common contaminant. It is believed that most of the aflatoxin contamination in rice bran occurs due to non-optimal storage conditions (Takahashi et al., 1989); however, a concentration of the toxin during milling of stored paddy rice also occurs, and the levels can triplicate compared with the grains (Trucksess et al., 2011).
The concentration of mycotoxins in DDGS during the ethanol production
Destillers’ dried grains with solubles (DDGS), a by-product of ethanol production, is a valuable feed ingredient, particularly as a source of protein for ruminants and monogastric animals at a competitive price.
Also here, mycotoxin contamination raises concerns with regard to their use in animal feeds. Mycotoxins are not destroyed during the ethanol fermentation process or during the production of DDGS. Moreover, a concentration of DON, ZEA, and fumonisin from corn to DDGS of 2–3.5 times has been reported for industrial ethanol production (Bennett et al., 1981; Schaafsma et al., 2009; Bowers & Munkvold, 2014).
In summary, studies on the fate of mycotoxins during food processing have shown that mycotoxins are concentrated in the fractions commonly used as animal feed. Moreover, high variability in mycotoxin contamination of cereal by-products has been evidenced, representing barriers to an increased acceptance of several food by-products as feed ingredients.
Feed formulation: Consider the mycotoxin contamination in by-products
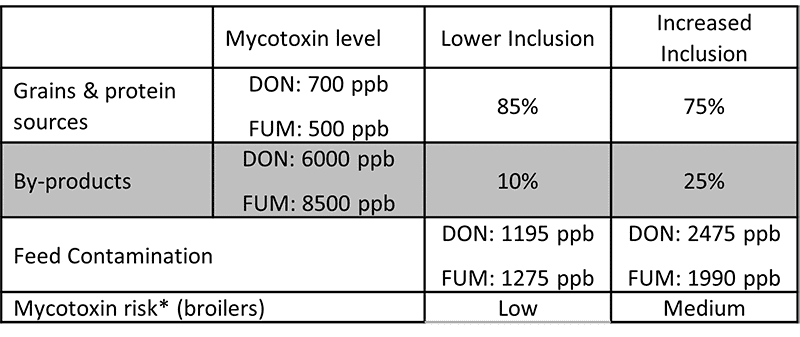
Higher inclusions of cereals have an impact on their safe use in feeds. To evaluate this impact, we can simulate two different scenarios with different inclusions of by-products:
Table 1: Different levels of by-products’ inclusion rates
In the first lower inclusion scenario, the risk for broilers in the starting phase considers the low inclusion of raw materials; the losses related to the contamination (without management) are mild. When increasing the levels of by-products, the risk category also increases. The losses are more important for the operation, ranging from gut barrier alterations with impaired production parameters to alterations in the immune response and increased susceptibility to disease.
Mycotoxins in by-products effective toxin risk management can help!
Given the pros of including cereal by-products in animal feeds, such as their saving potential and their link with sustainability of resources, their utilization is advisable; however, understanding how mycotoxin distribution and concentration change during grain processing is critical. Today’s knowledge is limited to a few mycotoxins in cereal milling.
Therefore, when considering using these by-products in the animal feed, we must bear in mind that:
modified mycotoxins and mycotoxin co-contamination can be present, contributing to additive/synergistic effects on animal health.
toxin risk management strategies, including analysis, risk evaluation, and risk mitigation must be pursued to prevent those undesired effects.
References:
Aureli, G., and M.G. D’Egidio. “Efficacy of Debranning on Lowering of Deoxynivalenol (DON) Level in Manufacturing Processes of Durum Wheat.” Tecnica Molit. 58 (2007): 729–33.
Bennett, G. A., A. A. Lagoda, O. L. Shotwell, and C. W. Hesseltine. “Utilization of Zearalenone- Contaminated Corn for Ethanol Production.” Journal of the American Oil Chemists’ Society 58, no. 11 (1981): 974–76. https://doi.org/10.1007/bf02659774.
Bowers, Erin, and Gary Munkvold. “Fumonisins in Conventional and Transgenic, Insect-Resistant Maize Intended for Fuel Ethanol Production: Implications for Fermentation Efficiency and DDGS Co-Product Quality.” Toxins 6, no. 9 (2014): 2804–25. https://doi.org/10.3390/toxins6092804.
Brera, Carlo, Carla Catano, Barbara de Santis, Francesca Debegnach, Marzia de Giacomo, Elena Pannunzi, and Marina Miraglia. “Effect of Industrial Processing on the Distribution of Aflatoxins and Zearalenone in Corn-Milling Fractions.” Journal of Agricultural and Food Chemistry 54, no. 14 (2006): 5014–19. https://doi.org/10.1021/jf060370s.
Brera,Carlo, Francesca, Debegnach, Silvana Grossi, and Marina Miraglia. “Effect of Industrial Processing on the Distribution of Fumonisin B1 in Dry Milling Corn Fractions.” Journal of Food Protection 67, no. 6 (2004): 1261–66. https://doi.org/10.4315/0362-028x-67.6.1261.
Cheli, Federica, Luciano Pinotti, Luciana Rossi, and Vittorio Dell’Orto. “Effect of Milling Procedures on Mycotoxin Distribution in Wheat Fractions: A Review.” LWT – Food Science and Technology 54, no. 2 (2013): 307–14. https://doi.org/10.1016/j.lwt.2013.05.040.
Park, Juhee, Dong-Ho Kim, Ji-Young Moon, Jin-Ah An, Young-Woo Kim, Soo-Hyun Chung, and Chan Lee. “Distribution Analysis of Twelve Mycotoxins in Corn and Corn-Derived Products by LC-MS/MS to Evaluate the Carry-over Ratio during Wet-Milling.” Toxins 10, no. 8 (2018): 319. https://doi.org/10.3390/toxins10080319.
Pascale, Michelangelo, Miriam Haidukowski, Veronica Maria Lattanzio, Marco Silvestri, Roberto Ranieri, and Angelo Visconti. “Distribution of T-2 and HT-2 Toxins in Milling Fractions of Durum Wheat.” Journal of Food Protection 74, no. 10 (2011): 1700–1707. https://doi.org/10.4315/0362-028x.jfp-11-149.
Pietri, A., M. Zanetti, and T. Bertuzzi. “Distribution of Aflatoxins and Fumonisins in Dry-Milled Maize Fractions.” Food Additives & Contaminants: Part A 26, no. 3 (2009): 372–80. https://doi.org/10.1080/02652030802441513.
Pinotti, Luciano, Matteo Ottoboni, Carlotta Giromini, Vittorio Dell’Orto, and Federica Cheli. “Mycotoxin Contamination in the EU Feed Supply Chain: A Focus on Cereal Byproducts.” Toxins 8, no. 2 (2016): 45. https://doi.org/10.3390/toxins8020045.
Ríos, G., L. Pinson-Gadais, J. Abecassis, N. Zakhia-Rozis, and V. Lullien-Pellerin. “Assessment of Dehulling Efficiency to Reduce Deoxynivalenol and Fusarium Level in Durum Wheat Grains.” Journal of Cereal Science 49, no. 3 (2009): 387–92. https://doi.org/10.1016/j.jcs.2009.01.003.
Schaafsma, Arthur W, Victor Limay-Rios, Diane E Paul, and J David Miller. “Mycotoxins in Fuel Ethanol Co-Products Derived from Maize: A Mass Balance for Deoxynivalenol.” Journal of the Science of Food and Agriculture 89, no. 9 (2009): 1574–80. https://doi.org/10.1002/jsfa.3626.
Schaarschmidt, Sara, and Carsten Fauhl-Hassek. “The Fate of Mycotoxins during the Processing of Wheat for Human Consumption.” Comprehensive Reviews in Food Science and Food Safety 17, no. 3 (2018): 556–93. https://doi.org/10.1111/1541-4337.12338.
Schollenberger, M., H.-M. Müller, M. Rüfle, S. Suchy, and W. Drochner. “Redistribution of 16FusariumToxins during Commercial Dry Milling of Maize.” Cereal Chemistry Journal 85, no. 4 (2008): 557–60. https://doi.org/10.1094/cchem-85-4-0557.
Takahashi, H., H. Yazaki, M. Manabe, S. Matsuura, and S. Kimura. “Distribution of Citrinin and Aflatoxins in Steamed Milled Rice Kernels Inoculated with Penicillium Citrinum and Aspergillus Flavus.” Mycotoxins 1990, no. 31 (1989): 49–53. https://doi.org/10.2520/myco1975.1990.49.
Trucksess, M.W., H.K. Abbas, C.M. Weaver, and W.T. Shier. “Distribution of Aflatoxins in Shelling and Milling Fractions of Naturally Contaminated Rice.” Food Additives & Contaminants: Part A 28, no. 8 (2011): 1076–82. https://doi.org/10.1080/19440049.2011.576441.
Improve health and productivity in breeders with phytomolecules
By Dr.Inge Heinzl, Editor, and Technical Team, EW Nutrition
Careful management of the breeders isa must to get their best reproductive efficiency. In today’s hatching egg production, factors such as stress, inflammation, body weight, and altered mating behaviorlead to decreased performance, meaning fewer hatchable eggs and, therefore, fewer day-old chicks per hen (Grandhaye, 2020). The use of antibiotics to increase performance in farm animals is no longer allowed in many countries, and, since it may lead to the development of resistance, it is also not recommended. So, also in breeders, alternatives are requested to maintain animal health, welfare, and a high level of performance.
Optimal gut health is the cornerstone for breeder performance
As the organ responsible for digestion of the incoming feed, the absorption of nutrients, and the defense of the organism against pathogens or toxins, a healthy gut is a pre-condition for optimal performance (Shini and Bryden, 2021). A healthy gut, according to Bailey (2018), has optimally developed gut tissues, a well-functioning gut immune system, and well-balanced gut microbiota. It shows efficient functionality in terms of digestion and absorption and protects the organism against harmful agents.
The gut directly or indirectly provides the elements for egg production
Efficient feed digestion and absorption of nutrients are essential for the breeder hen to obtain the “material” for maintenance, growth, and egg production. Gut health is crucial since dysbacteriosis and diarrhea, characteristics of gut health challenges, increase dirty eggs, creating favorable conditions for pathogens to enter the egg and infect the embryo.
Egg yolks consist of water (70%), proteins (10%), and lipids (20%). The yolk lipids are lipoproteins rich in triglycerides, built up in the liver and transported to the ovary. Cholesterol carried via lipoproteins to the egg yolk is also built up there, thus showing the importance of the liver in egg production. The gut plays a crucial role in protecting the liver from damage, constituting a barrier against harmful pathogens and toxins, potentially passing into the bloodstream and reaching this vital organ.
Phytomolecules support performance in different ways
Phytomolecules, are an excellent tool to support gut health and animal performance. Phytomolecules are plant-derived secondary metabolites that exert insect-attracting or defensive functions in the plant. They are used in their natural but also nature-identical forms in humans and animals to exert their digestive, immune-modulating, antimicrobial effects.
Phytomolecules support gut health by balancing the gut microbiome
Diverse examples can be found in the scientific literature, where phytomolecules improve the gut microbiome, resulting in better performance of layer and breeder hens. This support happens in two ways:
Promoting beneficial bacteria
Rabelo-Ruiz and co-workers (2021), asserted that adding garlic and onion extracts to the diet of layers led to more eggs with a bigger size, accompanied by an increase in Lactococci in the ileum and Lactobacilli in the cecum. Another example is provided by Park et al. (2016). When supplementing the diet of layers with a fermented phytogenic feed additive, egg production and weight raised with increasing dosage of the additive, and a higher number of Lactobacilli could be observed in the cecum.
Phytomolecules can promote the growth of certain beneficial bacteria and therefore act like prebiotics. As these changes took place in the lower gut, they assumed an improved digestibility of the feed.
Lowering pathogenic bacteria
In the study by Park et al. (2016) and in an in vitro study by Ghazanfari et al. (2019),E. coli in the cecum was reduced.
According to Burt (2007b), several essential oils / phytomolecules, amongst them, carvacrol, thymol, eugenol, and cinnamaldehyde, are effective against pathogens such as Listeria, Salmonella, E. coli, Shigella, and Staphylococcus. The hydrophobic essential oils can partition the lipids of the cell membranes. The resulting permeability of the membrane enables the leakage of cell content.
Changing virulence factors
Another mode of action is the change of virulence factors. Carvacrol, e.g., is known to decrease the motility of Campylobacter jejuni (Van Alphen et al., 2012); oregano and thyme oil reduced the motility of E. coli by inhibiting the synthesis of flagellin (Burt, 2007a). Vidanarachchi et al. (2005) mentioned that the hydrophobicity of microbes increases when some plant extracts are present, affecting their virulence characteristics. Also, the inhibition of defense measures such as efflux pumps in Gram-negative bacteria has been researched (Savoia, 2012).
Phytomolecules support gut health by improving digestion
For many years, phytomolecules have been studied and known for their digestive characteristics. In poultry and other animals, they influence feed digestion in two main ways.
Stimulating enzyme secretion
Platel and Srinivasan (2004) described different spices promoting not only the salivary flow, gastric juice and bile secretion but also the stimulation of the activity of enzymes such as pancreatic lipase, amylase, and proteases in rats. Hashemipour et al. (2013) saw the same effect in broilers supplemented with carvacrol and thymol in the diet. Research has also concluded on a higher nutrient digestibility: Hernandez et al. (2004) and Basmacioğlu Malayoğlu, 2010 noticed that supplementing plant extracts or essential oils improved apparent whole-tract and ileal digestibility of different nutrients.).
Maintaining gut integrity and enlarging the digestion area
An intact gut with a large area for digestion guarantees optimal utilization of nutrients. Different researchers found that adding plant extracts or essential oils (Khalaji et al., 2011; Ghazanfari et al., 2015; Chowdhury et al., 2018) promotes intestinal gut morphology, reflected in higher villi and deeper crypts, which might lead to higher nutrient absorption.
Concerning gut integrity, thymol and carvacrol showed protecting effects and mitigated gut lesions in broilers challenged with C. perfringens (Du et al., 2016). Probably, the lower pathogenic pressure due to the antimicrobial activity of phytogenic substances leads to minor damage to the gut wall and, in the end, to better absorption of the nutrients.
Phytomolecules mitigate the effects of stress
Environmental stress in breeders may decrease performance: the heat-stress-induced disruption of the tight junctions often leads to higher gut permeability, poor nutrient absorption, and higher electrolyte and water secretion (Abdelli, 2021). Sahin et al. (2010) achieved a linear improvement in egg production in quails when applying two doses of green tea catechin.
Cold-stressed layers also reacted positively to supplementation of oregano essential oil, improving egg production compared to a non-supplemented control (Migliorini, 2019).
Positive influence of phytomolecules results in higher performance
As described, phytomolecules improve gut health and support the animal in multiply ways, allowing better utilization of resources for growth and production. Literature provides many articles showing the promoting effects of these substances on the performance of layers or breeders, some of them summarized in Table 1.
Table 1: Benefits of phytomolecules in layers and breeders
Compounds
Reference
Main effects: Improved egg weight, egg mass, and higher hen-day-egg production
In-feed and in-water phytomolecules-based products show efficacy
Much of the research done with phytomolecules focuses on essential oils (with variable inclusions of the active compounds or on single plant extracts. EW Nutrition is a research-driven company proposing phytomolecule-based solutions for the animal production industry. These products combine selected, synergistically acting phytomolecules to achieve optimal results.
EW Nutrition has tested the combined use of
a microencapsulated blend of phytomolecules (Activo) for the feed and designed to maintain a good gut-health status during the whole life-cycle of the breeders, and
Activo Liquid, a liquid combination of phytomolecules and organic acids, which is conveniently applied on the farm via the waterline.
A trial conducted at the University of Central Queensland (Australia) showed that phytomolecules enhance beneficial bacteria such as Lactobacilli and, on the other hand, repress harmful bacteria such as Clostridium perfringens.
For the trial, caecal microbiota of layers was used. They were grown with and without Activo Liquid in vitro, and the changes in microbiota were monitored.
Result: The in vitro study clearly shows that Activo Liquid increases the number of lactobacilli and decreases clostridia and Enterococcus sp.
Cie Chart
Figure 1: Shifting intestinal balance with phytomolecules
2.Three field trials with Activo Liquid showed an increased laying rate in breeders
Many operations started testing phytomolecules in a farm-application-based program to reaffirm the gut health-improving activity of phytomolecules in broiler breeder performance. Especially the flexibility of assisting animals through the water for drinking during stress periods makes phytomolecules an optimal tool to support gut health.
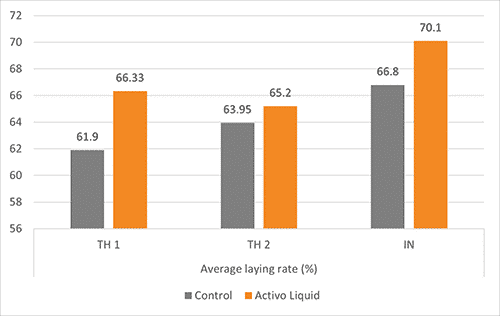
Two broiler breeder farms in Thailand (TH1 and TH2) and one grandparent farm in India (IN) are good examples of the effectiveness of phytomolecules. On each farm, the birds were always divided into two groups. Besides the standard management, feed, and water, one group got 200 ml Activo Liquid per 1,000 L of water. The periods when the birds received Activo in the water differed:
TH1 & TH2: 5 days per week, during weeks 24 – 32
IN: 5 days per week, every third week from weeks 18 to 24 and every fourth week from 28 to 36
The trials lasted for 9 weeks (Thailand 1 and 2) and 30 weeks (India).
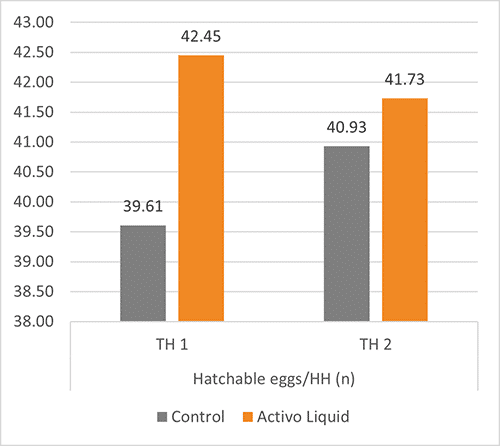
The results are shown in figure 2. The animals supplemented with Activo Liquid showed an up to 4.4 % higher laying rate and up to three more hatchable eggs per hen housed.
Figure 2+3: Results of three trials conducted In Asia concerning laying rate and hatchable eggs
3. Customers tell about lower breeder mortality and more DOCs due to phytomolecules
The benefits of a tailored phytomolecule program have been demonstrated in several broiler breeder operations worldwide. For example, a combination of the in-feed (Activo) and the in-water solution (Activo Liquid) was tested in the Middle East. For the study, 75,000 23-weeks-old broiler breeders were divided into groups: 4 houses with the program, and 6 houses served as control (standard feed and water). The program, tailored to customer needs, was designed as follows:
AC+AL group:
Activo 100 g/ton of feed during the whole trial (weeks 23-41) +
Activo Liquid 250 ml/1000 L water, four days per week, weeks 23-30.
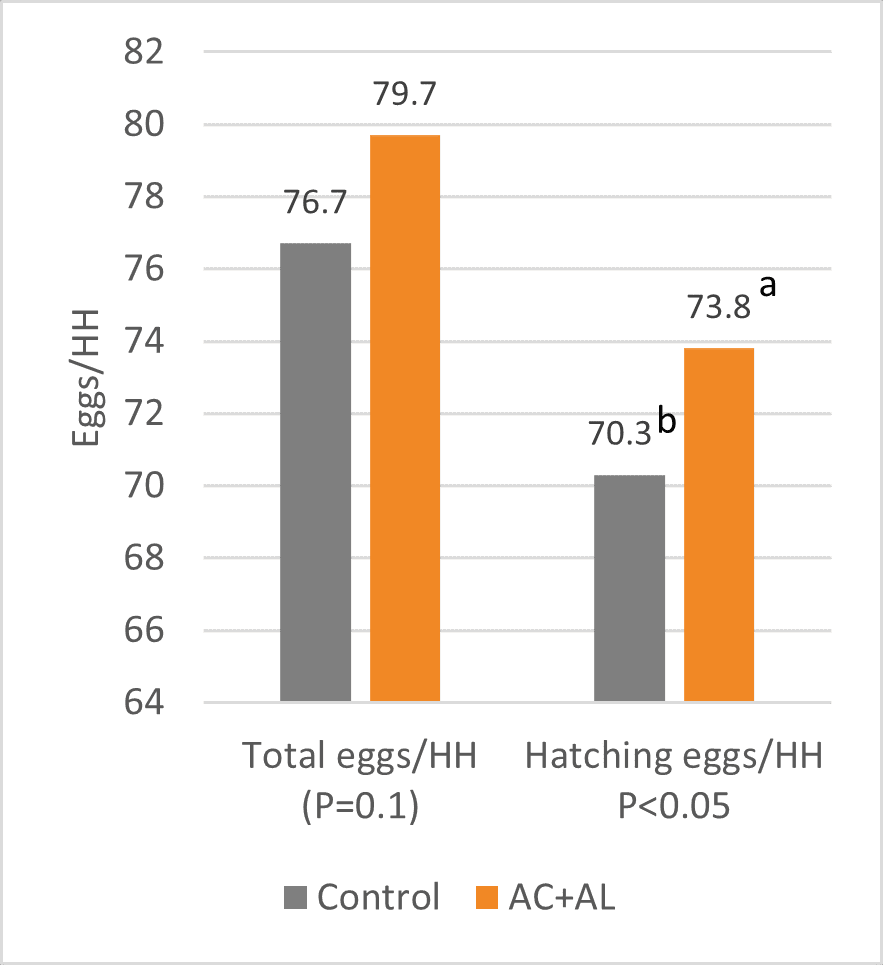
As a result, the peak and average laying rates were higher for the flocks with the program, and laying persistency was also higher. This allowed for a significant difference of 3 total and 3.5 hatching eggs/hen housed at week 41. In both cases, an increase equivalent to 5 % compared to the control group (figure 4) could be observed.
Figure 4: Total eggs and hatching eggs per hen housed
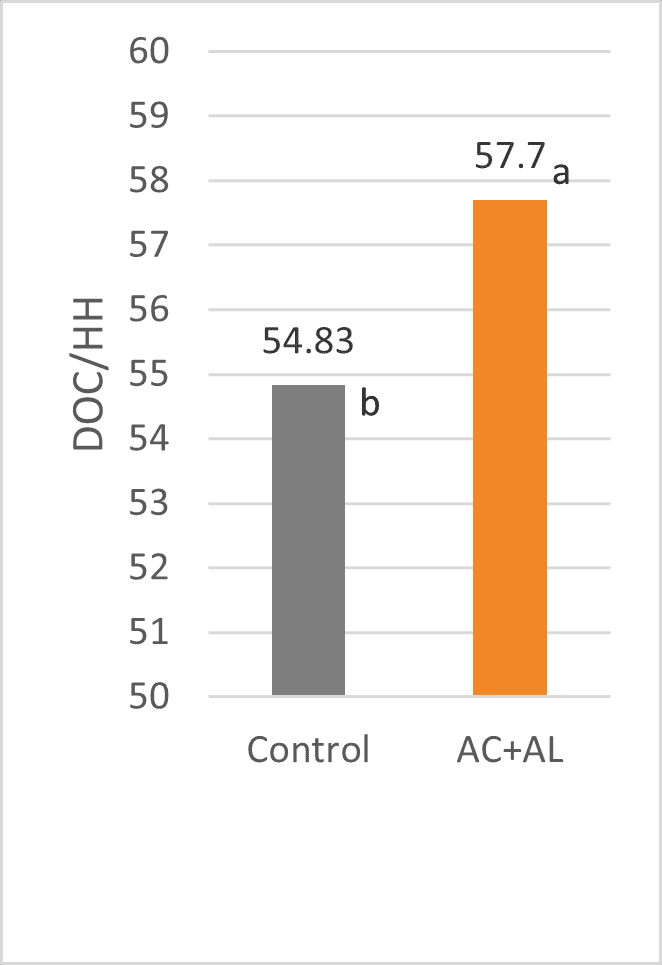
As fertility and hatchability were similar for both groups, the 5 % increase in hatching eggs resulted in a 5 % higher number of day-old chicks per hen housed (figure 5).
Figure 5: Number of DOSs per hen housed
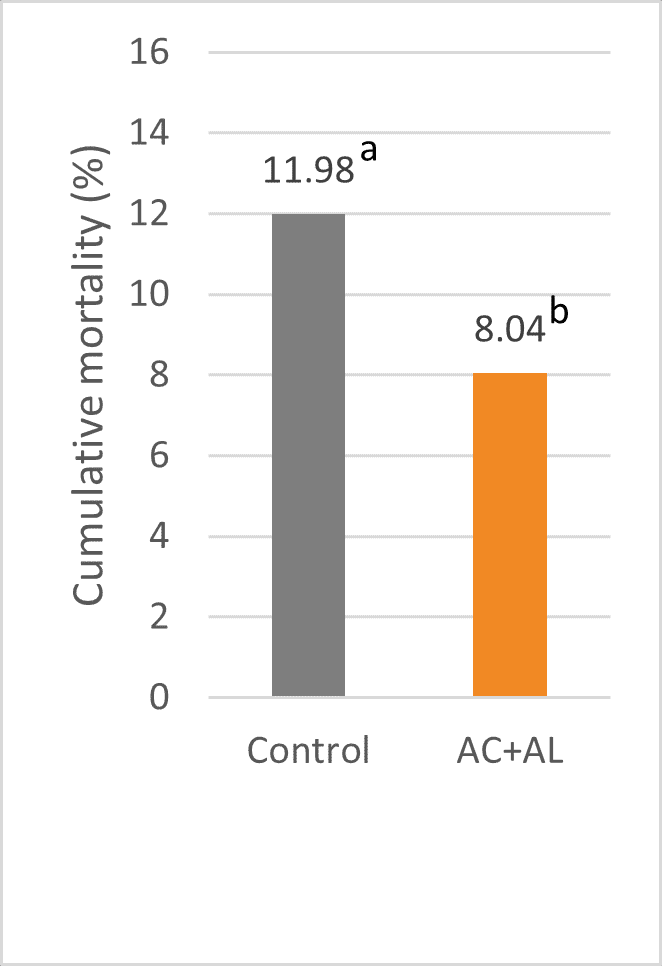
It must be mentioned that during the trial period, at 28 weeks of age, an NDV outbreak was diagnosed on the farm, which negatively impacted the overall results. However, this impact was reduced in the groups receiving the phytomolecule-based products, which also was reflected in a lower mortality rate (figure 6).
Figure 6: Cumulative mortality rate wk 41
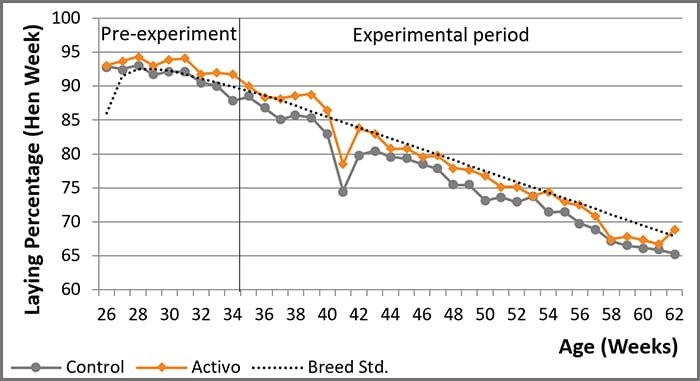
4. Scientific trial shows that Activo can increase post-peak productivity in breeders
When thinking about the use of phytomolecules, most broiler breeder operations would like to consider scientific trial results in this type of animal. For EW Nutrition, it is crucial to accurately evaluate every product that reaches a market. Thus several scientific trials with broiler breeders have been performed. For one of them, Hubbard breeders (JA57 females with 80 M77 males) were divided into 2 treatments, having 5 replicate pens for each. The experiment started after the peak production period, at 34 weeks of age, and ended at week 62. To make the trial fair, the production data of 6 (pre-experimental) weeks was used to allocate the pens for each treatment, resulting in two (statistically) similar groups.
The control group was fed the standard mash diet. For the Activo group, 100g Activo/MT was added to the diet.
With Activo, breeders kept their high productivity after the peak, while the control group showed a steady decline from breed target values. During the experiment, Activo supplemented birds produced 3.6 more eggs than control birds (P=0.06) while consuming a similar amount of feed. As a result, a lower feed consumption per egg produced was achieved (169.9 vs. 173.6 g/egg, respectively).
As the dietary treatment did not influence hatchability, the 3.6 extra eggs resulted in 2.9 extra day-old chicks per hen during the post-peak period, showing a positive return.
Phytomolecules as gut health and performance promoters– antibiotics can be reduced!
With their gut health-promoting activity, phytomolecules support breeders to better utilize nutrients. They can be invested for maintenance and the production of hatchable eggs, obtaining good quality day-old chicks.
References
Abad, Paloma, Natalia Arroyo-Manzanares, Juan J. Ariza, Alberto Baños, and Ana M. García-Campaña. “Effect of Allium Extract Supplementation on Egg Quality, Productivity, and Intestinal Microbiota of Laying Hens.” Animals 11, no. 1 (2020): 41. https://doi.org/10.3390/ani11010041.
Abdel-Wareth, A.A.A., and J.D. Lohakare. “Effect of Dietary Supplementation of Peppermint on Performance, Egg Quality, and Serum Metabolic Profile of Hy-Line Brown Hens during the Late Laying Period.” Animal Feed Science and Technology 197 (2014): 114–20. https://doi.org/10.1016/j.anifeedsci.2014.07.007.
Abdel-Wareth, A.A.A., Z.S.H. Ismail, and K.-H. Südekum. “Effects of Thyme and Oregano on Performance and Egg Quality Characteristics of Laying Hens.” World’s Poultry Sci. Journal, Supplement, 69 (2013): 1–6.
Abdelli, Nedra, David Solà-Oriol, and José Francisco Pérez. “Phytogenic Feed Additives in Poultry: Achievements, Prospective and Challenges.” Animals 11, no. 12 (2021): 3471. https://doi.org/10.3390/ani11123471.
Abou-Elkhair, Reham, Shaimaa Selim, and Eman Hussein. “Effect of Supplementing Layer Hen Diet with Phytogenic Feed Additives on Laying Performance, Egg Quality, Egg Lipid Peroxidation and Blood Biochemical Constituents.” Animal Nutrition 4, no. 4 (2018): 394–400. https://doi.org/10.1016/j.aninu.2018.05.009.
Akbari, Mohsen, Mehran Torki, and Keyomars Kaviani. “Single and Combined Effects of Peppermint and Thyme Essential Oils on Productive Performance, Egg Quality Traits, and Blood Parameters of Laying Hens Reared under Cold Stress Condition (6.8 ± 3 °C).” International Journal of Biometeorology 60, no. 3 (2015): 447–54. https://doi.org/10.1007/s00484-015-1042-6.
Al-Harthi, M.A. “Responses of Laying Hens to Different Levels of Amoxicillin, Hot Pepper or Green Tea and Their Effects on Productive Performance, Egg Quality and Chemical Composition of Yolk and Blood Plasma Constituents.” Egypt. Poult. Sci. 24, no. IV (2004): 845–68.
Al-Yasiry, A.R.M., B. Kiczorowska, W. Samolińska, E. Kowalczuk-Vasilev, and D. Kowalczyk-Pecka. “The Effect of Boswellia Serrata Resin Diet Supplementation on Production, Hematological, Biochemical and Immunological Parameters in Broiler Chickens.” Animal 11, no. 11 (2017): 1890–98. https://doi.org/10.1017/s1751731117000817.
Ali, Akhtar, Eric N. Ponnampalam, Gamini Pushpakumara, Jeremy J. Cottrell, Hafiz A. Suleria, and Frank R. Dunshea. “Cinnamon: A Natural Feed Additive for Poultry Health and Production—a Review.” Animals 11, no. 7 (2021): 2026. https://doi.org/10.3390/ani11072026.
Bailey, Richard. “Maintaining Gut Health without Antibiotics.” Poultry World, April 19, 2021. https://www.poultryworld.net/health-nutrition/maintaining-gut-health-without-antibiotics/.
Basmacioğlu Malayoğlu, H., Ş. Baysal, Z. Misirlioğlu, M. Polat, H. Yilmaz, and N. Turan. “Effects of Oregano Essential Oil with or without Feed Enzymes on Growth Performance, Digestive Enzyme, Nutrient Digestibility, Lipid Metabolism and Immune Response of Broilers Fed on Wheat–Soybean Meal Diets.” British Poultry Science 51, no. 1 (2010): 67–80. https://doi.org/10.1080/00071660903573702.
Burt, Sara A., Ruurd van der Zee, Ad P. Koets, Anko M. de Graaff, Frans van Knapen, Wim Gaastra, Henk P. Haagsman, and Edwin J. Veldhuizen. “Carvacrol Induces Heat Shock Protein 60 and Inhibits Synthesis of Flagellin in Escherichia Coli O157:H7.” Applied and Environmental Microbiology 73, no. 14 (2007a): 4484–90. https://doi.org/10.1128/aem.00340-07.
Burt, Sara Ann. “Antibacterial Activity of Essential Oils: Potential Applications in Food.” Thesis, S. Burt, 2007b.
Bölükbasi, S. Canan, and M. Kuddusi Erhan. “Effect of Dietary Thyme (Thymus Vulgaris) on Laying Hens Performance and Escherichia Coli (E. Coli) Concentration in Feces.” International Journal of Natural and Engineering Sciences 1, no. 2 (2007): 55–58.
Chowdhury, Subrata, Guru Prasad Mandal, Amlan Kumar Patra, Pawan Kumar, Indranil Samanta, Saktipada Pradhan, and Arup Kumar Samanta. “Different Essential Oils in Diets of Broiler Chickens: 2. Gut Microbes and Morphology, Immune Response, and Some Blood Profile and Antioxidant Enzymes.” Animal Feed Science and Technology 236 (2018): 39–47. https://doi.org/10.1016/j.anifeedsci.2017.12.003.
Dilawar, Muhammad Ammar, Hong Seok Mun, Dhanushka Rathnayake, Eun Ju Yang, Ye Seul Seo, Hyeoung Seog Park, and Chul Ju Yang. “Egg Quality Parameters, Production Performance and Immunity of Laying Hens Supplemented with Plant Extracts.” Animals 11, no. 4 (2021): 975. https://doi.org/10.3390/ani11040975.
Du, Encun, Weiwei Wang, Liping Gan, Zhui Li, Shuangshuang Guo, and Yuming Guo. “Effects of Thymol and Carvacrol Supplementation on Intestinal Integrity and Immune Responses of Broiler Chickens Challenged with Clostridium Perfringens.” Journal of Animal Science and Biotechnology 7, no. 1 (2016). https://doi.org/10.1186/s40104-016-0079-7.
Franciosini, Maria Pia, Patrizia Casagrande-Proietti, Claudio Forte, Daniela Beghelli, Gabriele Acuti, Dario Zanichelli, Alessandro dal Bosco, Cesare Castellini, and Massimo Trabalza-Marinucci. “Effects of Oregano (Origanum Vulgarel.) and Rosemary (Rosmarinus Officinalis l.)Aqueous Extracts on Broiler Performance, Immune Function and Intestinal Microbial Population.” Journal of Applied Animal Research 44, no. 1 (2015): 474–79. https://doi.org/10.1080/09712119.2015.1091322.
Ghazanfari, S, Z Mohammadi, and M Adib Moradi. “Effects of Coriander Essential Oil on the Performance, Blood Characteristics, Intestinal Microbiota and Histological of Broilers.” Revista Brasileira de Ciência Avícola 17, no. 4 (2015): 419–26. https://doi.org/10.1590/1516-635×1704419-426.
Grandhaye, Jeremy, Veronique Douard, Ana Rodriguez-Mateos, Yifan Xu, Alex Cheok, Antonella Riva, Rodrigo Guabiraba, et al. “Microbiota Changes Due to Grape Seed Extract Diet Improved Intestinal Homeostasis and Decreased Fatness in Parental Broiler Hens.” Microorganisms 8, no. 8 (2020): 1141. https://doi.org/10.3390/microorganisms8081141.
Hashemipour, H., H. Kermanshahi, A. Golian, and T. Veldkamp. “Effect of Thymol and Carvacrol Feed Supplementation on Performance, Antioxidant Enzyme Activities, Fatty Acid Composition, Digestive Enzyme Activities, and Immune Response in Broiler Chickens.” Poultry Science 92, no. 8 (2013): 2059–69. https://doi.org/10.3382/ps.2012-02685.
Hernández, F., J. Madrid, V. García, J. Orengo, and M.D. Megías. “Influence of Two Plant Extracts on Broilers Performance, Digestibility, and Digestive Organ Size.” Poultry Science 83, no. 2 (2004): 169–74. https://doi.org/10.1093/ps/83.2.169.
Khalaji, S., M. Zaghari, K.H. Hatami, S. Hedari-Dastjerdi, L. Lotfi, and H. Nazarian. “Black Cumin Seeds, Artemisia Leaves (Artemisia Sieberi), and Camellia L. Plant Extract as Phytogenic Products in Broiler Diets and Their Effects on Performance, Blood Constituents, Immunity, and Cecal Microbial Population.” Poultry Science 90, no. 11 (2011): 2500–2510. https://doi.org/10.3382/ps.2011-01393.
Khan, Sohail Hassan, Muhammad Ashraf Anjum, Abida Parveen, Tabinda Khawaja, and Naeem Mehmood Ashraf. “Effects of Black Cumin Seed (Nigella Sativa) on Performance and Immune System in Newly Evolved Crossbred Laying Hens.” Veterinary Quarterly 33, no. 1 (2013): 13–19. https://doi.org/10.1080/01652176.2013.782119.
Kim, Duk Kyung, Hyun S. Lillehoj, Sung Hyen Lee, Erik P. Lillehoj, and David Bravo. “Improved Resistance to Eimeria Acervulina Infection in Chickens Due to Dietary Supplementation with Garlic Metabolites.” British Journal of Nutrition 109, no. 1 (2012): 76–88. https://doi.org/10.1017/s0007114512000530.
Kothari, Damini, Woo-Do Lee, Kai-Min Niu, and Soo-Ki Kim. “The Genus Allium as Poultry Feed Additive: A Review.” Animals 9, no. 12 (2019): 1032. https://doi.org/10.3390/ani9121032.
Lutful Kabir, S. M. “Avian Colibacillosis and Salmonellosis: A Closer Look at Epidemiology, Pathogenesis, Diagnosis, Control and Public Health Concerns.” International Journal of Environmental Research and Public Health 7, no. 1 (2010): 89–114. https://doi.org/10.3390/ijerph7010089.
Migliorini, Marcos J., Marcel M. Boiago, Lenita M. Stefani, Aline Zampar, Lenilson F. Roza, Mauricio Barreta, Alessandra Arno, et al. “Oregano Essential Oil in the Diet of Laying Hens in Winter Reduces Lipid Peroxidation in Yolks and Increases Shelf Life in Eggs.” Journal of Thermal Biology 85 (2019): 102409. https://doi.org/10.1016/j.jtherbio.2019.102409.
Mohiti-Asli, Maziar, and Moein Ghanaatparast-Rashti. “Comparison of the Effect of Two Phytogenic Compounds on Growth Performance and Immune Response of Broilers.” Journal of Applied Animal Research 45, no. 1 (2017): 603–8. https://doi.org/10.1080/09712119.2016.1243119.
Nadia, L.,, Radwan, R.A. Hassan, E.M. Qota, and H.M. Fayek. “Effect of Natural Antioxidant on Oxidative Stability of Eggs and Productive and Reproductive Performance of Laying Hens.” International Journal of Poultry Science 7, no. 2 (2008): 134–50. https://doi.org/10.3923/ijps.2008.134.150.
Nadia, L.,, Radwan, R.A. Hassan, E.M. Qota, and H.M. Fayek. “Effect of Natural Antioxidant on Oxidative Stability of Eggs and Productive and Reproductive Performance of Laying Hens.” International Journal of Poultry Science 7, no. 2 (2008): 134–50. https://doi.org/10.3923/ijps.2008.134.150.
Park, Jae Hong, Tae Hwa Song, and Inho Kim. “Egg Production, Egg Quality, and Cecal Microbial Populations of Layers fed Diets Supplemented with Fermented Phytogenic Feed Additive.” TURKISH JOURNAL OF VETERINARY AND ANIMAL SCIENCES 40 (2016): 660–66. https://doi.org/10.3906/vet-1512-55.
Platel, Kalpana, and K. Srinivasan. “Digestive Stimulant Action of Spices: a Myth or Reality?” Indian J Med Res. 119, no. 5 (2004): 167–79.
Puvača, Nikola, Erinda Lika, Sonila Cocoli, Tana Shtylla Kika, Vojislava Bursić, Gorica Vuković, Mirela Tomaš Simin, Aleksandra Petrović, and Magdalena Cara. “Use of Tea Tree Essential Oil (Melaleuca Alternifolia) in Laying Hen’s Nutrition on Performance and Egg Fatty Acid Profile as a Promising Sustainable Organic Agricultural Tool.” Sustainability 12, no. 8 (2020): 3420. https://doi.org/10.3390/su12083420.
Rabelo-Ruiz, Miguel, Juan José Ariza-Romero, María Jesús Zurita-González, Antonio Manuel Martín-Platero, Alberto Baños, Mercedes Maqueda, Eva Valdivia, Manuel Martínez-Bueno, and Juan Manuel Peralta-Sánchez. “Allium-Based Phytobiotic Enhances Egg Production in Laying Hens through Microbial Composition Changes in Ileum and Cecum.” Animals 11, no. 2 (2021): 448. https://doi.org/10.3390/ani11020448.
Ren, Hao, Wilfried Vahjen, Temesgen Dadi, Eva-Maria Saliu, Farshad Goodarzi Boroojeni, and Jürgen Zentek. “Synergistic Effects of Probiotics and Phytobiotics on the Intestinal Microbiota in Young Broiler Chicken.” Microorganisms 7, no. 12 (2019): 684. https://doi.org/10.3390/microorganisms7120684.
Sahin, K., C. Orhan, M. Tuzcu, S. Ali, N. Sahin, and A. Hayirli. “Epigallocatechin-3-Gallate Prevents Lipid Peroxidation and Enhances Antioxidant Defense System via Modulating Hepatic Nuclear Transcription Factors in Heat-Stressed Quails.” Poultry Science 89, no. 10 (2010): 2251–58. https://doi.org/10.3382/ps.2010-00749.
Shini, S., and W. L. Bryden. “Probiotics and Gut Health: Linking Gut Homeostasis and Poultry Productivity.” Animal Production Science, 2021. https://doi.org/10.1071/an20701.
van Alphen, Lieke B., Sara A. Burt, Andreas K. Veenendaal, Nancy M. Bleumink-Pluym, and Jos P. van Putten. “The Natural Antimicrobial Carvacrol Inhibits Campylobacter Jejuni Motility and Infection of Epithelial Cells.” PLoS ONE 7, no. 9 (2012). https://doi.org/10.1371/journal.pone.0045343.
Vidanarachchi, J.K., L.L. Mikkelsen, I. Sims, P.A. Iji, and M. Choct. “Phytobiotics: Alternatives to Antibiotic Growth Promoters in Monogastric Animal Feeds.” Recent Advances in Animal Nutrition in Australia 15 (2005): 131–44.
Wang, Xiao-cui, Xiao-hong Wang, Jing Wang, Hao Wang, Hai-jun Zhang, Shu-geng Wu, and Guang-hai Qi. “Dietary Tea Polyphenol Supplementation Improved Egg Production Performance, Albumen Quality, and Magnum Morphology of Hy-Line Brown Hens during the Late Laying period1.” Journal of Animal Science 96, no. 1 (2018): 225–35. https://doi.org/10.1093/jas/skx007.
Zhu, Nianhua, Jun Wang, Longfei Yu, Qiman Zhang, Kai Chen, and Baosheng Liu. “Modulation of Growth Performance and Intestinal Microbiota in Chickens Fed Plant Extracts or Virginiamycin.” Frontiers in Microbiology 10 (2019). https://doi.org/10.3389/fmicb.2019.01333.
Exploding energy prices? Manage moisture to improve feed mill efficiency
By Ivan Ilić, Global Manager Technical Product Applications, EW Nutrition
Modern large-scale feed mills operate extremely efficiently and have few variable costs that could be reduced to lower the total cost of the final feed (Stark, 2012).In light of worrying energy price hikes, feed producers, however, should reduce their electricity use per unit produced, to maintain profitability. Find out how optimizing the feed mill’s moisture management increases feed quality while decreasing the energy required to produce it.
Due to climatic challenges, variability in raw material quality, and technical constraints, it can be challenging for feed producers to stabilize the water content in compound feed across time, raw material batches or even different machinery.
Combined with high temperatures, high moisture in feed can favor the growth of molds. They spoil feed, depleting energy and nutrients and generating reactive oxygen species (ROS) that reduce feed palatability. Even worse, some molds release toxins harm animals’ health and performance. On the other hand, low moisture levels in feed has a negative impact on pellet durability, increasing fines, process loss, and energy consumption while decreasing pellet press yield (Moritz et al., 2002).
What does feed moisture management have to do with a feed mill’s electricity consumption?
Moisture from raw materias can be lost during storage and processing. Silo aeration and enviroment conditions can contribute to moisture loss when the grains are stored at higher than optimal moisture levels (Angelovič, 2018). During feed processing, the intense friction of grinding results in heat and moisture from the grains is lost as vapor. As an optimal level of moisture is critical to ensure production output and feed quality, it must be added back to the system and adequately managed to keep or increase final feed quality.
For pelleted feeds, managing moisture is a two-step process:
Adding moisture in the mixer. This ensures that the mash feed is enters the conditioning process at the right moisture level, facilitating the penetration of steam and increasing the efficiency of the process.
Managing steam during conditioning. Steam added to the conditioner must be dry (meaning saturated with water droplets in suspension), and when this dry steam contacts the feed, it condenses and adds moisture.
However, simply adding water into the mixer does not give optimal results: Pure water does not completely bind to the feed; it mostly “sits on top” of the feed surface, increasing its water activity, and thus increasing the danger of microbial growth. Plus, a high proportion of pure water evaporates again when the feed is cooled.
Surfactants improve moisture retention
Surfactants change the way water behaves: by reducing the surface tension of water, they enable the feed particles to absorb the water and ensure that it is evenly distributed throughout the feed.
Improved moisture retention can:
facilitate the starch gelatinization during conditioning (important making the pellet more durable and the feed more digestible),
minimize feed shrinkage,
reduce friction and hence the energy required for the pellet die (improving milling efficiency), and
curb microbial growth by reducing water activity.
SURF•ACE: Improve throughput and reduce energy requirements
While surfactants contribute to mold control, feed producers also require the help of organic acids such as propionic acid (cf. Smith et al., 1983). The objectives are to optimize the moisture content in feed and to reduce its mold contamination. EW Nutrition’s SURF•ACETM feed mill processing aid combines organic acids and surfactants to achieve the objective of adding moisture without risking either the significant loss of moisture during cooling or the development of mold.
The effect of adding SURF•ACE to diets with different levels of fat was evaluated at more than 40 feed mills, with production capacities ranging from 5 to 20 tons per hour. SURF•ACE is added to water sprayed during mixing. This hydrating solution lubricates the mash feed, improves steam penetration and starch gelatinization, and reduces friction in the pellet dies. The results show that, relative to pure water, the addition of SURF•ACE increases press throughout (t/h) by between 5 and 25 %.
Trial results: SURF•ACE increases press yields while lowering energy consumption
For a trial at a Turkish beef and poultry feed mill, the same feed was run through the pelletizer in two batches, one with a 1 % water and one with 1% water mixed with 200 g of SURF•ACE per ton of feed. Adding SURF•ACE resulted in higher pellet output (6% for beef; 9% for poutry) and reduced energy consumption (13% for both beef and poultry):
In Poland, another trial conducted at a commercial feed mill found that when SURF•ACE was added to 1% mixer-moisture, this lead to a 28.6 % higher feed throughput in the pellet press, 23 % lower energy consumption per unit produced during the pelleting process, and a nearly 1 %-point higher moisture content in finished feed. This resulted in higher profitability: based on the costs in Poland at the time of the trial, an ROI of 2.4:1 was achieved.
A recent trial at an Indian feed mill evaluated the difference between adding 1% moisture to produce crumble feed (control group) and upgrading the water with 200 g of SURF•ACE per ton. The addition of SURF•ACE reduced power consumption by 6% and improved throughput by 18%.
Feed mills must deal with rising energy costs head-on
Operating in a tight margin environment, feed mills always need to prioritize efficiency. The advantages of using SURF•ACE feed mill processing aid are clear: reduced energy consumption, better pellet quality, fewer fines, better PDI, moisture optimization, lower maintenance costs, and higher productivity (throughput). During times of increasingly high ingredient and energy costs, it is even more important to utilize savings opportunities at every production stage. Thanks to its dual surfactant and preservative effects, SURF•ACE enables feed mills to improve feed quality and increase throughput while lowering electricity use.
References
Angelovič, Marek, Koloman Krištof, Ján Jobbágy, Pavol Findura, and Milan Križan. “The effect of conditions and storage time on course of moisture and temperature of maize grains.” BIO Web Conferences 10 (2018): 02001. https://doi.org/10.1051/bioconf/20181002001
Moritz, J. S., K. J. Wilson, K. R. Cramer, R. S. Beyer, L. J. McKinney, W. B. Cavalcanti, and X. Mo. “Effect of Formulation Density, Moisture, and Surfactant on Feed Manufacturing, Pellet Quality, and Broiler Performance.” Journal of Applied Poultry Research 11, no. 2 (2002): 155–63. https://doi.org/10.1093/japr/11.2.155.
Smith, Philip A., Talmadge S. Nelson, Linda K. Kirby, Zelpha B. Johnson, and Joseph N. Beasley. “Influence of Temperature, Moisture, and Propionic Acid on Mold Growth and Toxin Production on Corn.” Poultry Science 62, no. 3 (1983): 419–23. https://doi.org/10.3382/ps.0620419.
Piglet performance with fewer antimicrobials is possible
By Technical Team, EW Nutrition
A variety of stressors simultaneously occur at weaning, making this probably the most challenging period in pig production. During weaning, we commonly see altered gut development and gut microbiome, which increases piglets’ vulnerability to diseases. The most classic clinical symptom resulting from these stressors is the occurrence of post-weaning diarrhea. It is a sign that something went wrong, and piglet development and overall performance may be compromised (Guevarra et al. 2019).
Besides weaning, an unavoidable practice in pig production, the swine industry has been facing other changes. Among them, the increased pressure to reduce the use of antimicrobials stands out. Antimicrobials are often associated with improved piglet performance and health. Their usage has been reduced worldwide, however, due to the threat of antimicrobial resistance that affects not just animal health but also human health (Cardinal et al., 2019).
Reduce antimicrobials and post-weaning diarrhea: can piglet nutrition achieve both?
With these drastic changes for the piglets and the global swine industry, producers must find solutions to keep their farms profitable — especially from a nutritional perspective. Our last article presented two feed additives that can be part of an antibiotic-free concept for post-weaning piglets. This article will highlight a few essential nutritional strategies that swine producers and nutritionists must consider when formulating post-weaning feed without or with reduced amounts of antimicrobials.
What makes weaning so stressful for piglets?
Producers, nutritionists, and veterinarians all agree that weaning is a tough time for piglets (Yu et al., 2019) and, therefore, a challenge to all those involved in the pig production chain. Although there is a global trend towards increasing weaning age, generally speaking, animals are still immature when going through the weaning process. They face several physiological, nutritional, and environmental changes (figure 1).
Figure 1. Factors associated with weaning can compromise piglet well-being and performance
Most of these changes become “stressors” that trigger a cascade of reactions affecting the balance and morphology of the intestinal microbiome (figure 2). The outcome is a decrease in the piglets’ well-being and, in most cases, performance. We need to clearly understand how these stressors affect pigs to develop effective strategies against post-weaning growth impairments, especially when no antimicrobials are allowed.
Figure 2. Schematic diagram illustrating the effects of stress in weaned piglets (adapted from Jayaraman and Nyachoti, 2017)
Weaning support starts before weaning
The use of creep feed has been evaluated and even criticized for many years. Some operations are still reluctant to use such a feed due to its high cost and amount of labor on the farm, with manually providing feed and cleaning feeding trays. In addition, some questions have been raised regarding the ideal composition of the creep feed – how much complexity should we add to this special diet?
Therefore, the benefits of creep feed are under re-evaluation, not only considering piglet physiology per se, but also feed characteristics and different feeding programs. Recent studies have questioned highly complex creep feed formulations. Creep feed is being called “transition feed” (Molist, 2021) – i.e., that meal which is complementary to sows’ milk and not a replicate of it, helping piglets during the period of changing its main source of nutrients. We must, therefore, look at it as a way of making piglets familiar with solid feed, as highlighted by Mike Tokach during the 2020 KSU Swine Day. Dr. Tokach also mentioned that the presence of feeders in the lactation pen could stimulate the exploratory behaviors of the piglets. Combined, these practices can lead to a higher feed intake and performance during the nursery phase.
Towards a pragmatic stance on creep feed
Heo et al. (2018) evaluated three different creep feed types: a highly digestible creep feed, weaning feed as creep feed, and sow feed as creep feed until weaning. Piglets receiving the highly digestible creep had higher feed intake during the second to the last week pre-weaning (14 to 21 days of age) and higher ADG during the last week pre-weaning (21 to 28 days of age). This resulted in a trend for higher weaning weight. However, these benefits did not persist after weaning when all piglets received the weaning feed.
Guevarra et al. (2019) also suggested that the abrupt transition in piglet nutrition to a more complex nutrient source can influence shifts in the gut microbiota, impacting the absorptive capacity of the small intestine. Yang et al. (2016) evaluated 40 piglets from eight litters during the first week after weaning. They found that the change in diet during weaning reduced the proliferation of intestinal epithelial cells. This indicates that this period affects cellular macromolecule organization and localization, in addition to energy and protein metabolism. These results suggest that “similarity” in feed pre- and post-weaning may contribute more to the continuity of nutrient intake post-weaning than a highly complex-nutrient dense creep feed.
Nutritional strategies without antibiotics: focus on pig physiology
As mentioned, it is crucial to avoid a drastic drop in feed/nutrient intake after weaning compared to pre-weaning levels. In a classic study, Pluske et al. (1996) showed the importance of high intake levels on villus weight (used as a reference for gut health, cf. graph 1). Although not desirable, the reduction should be considered “normal” behavior.
Imagine these recently weaned piglets, facing all these stressors, having to figure out within this new group of peers when it is time to eat, where to find food, why water and food now come from two distinct sources… Therefore, management, feeding, water quality, and other aspects play important roles in post-weaning feed intake (figure 3).
Graph 1. Villus height following different levels of feed intake (M = maintenance) post-weaning (a.b.c bars with unlike superscript letters are different at P<0.05). (From Pluske et al., 1996.)
From a nutritional perspective, piglets at weaning experience a transition from milk (a high-fat, low-carbohydrate liquid) to a plant-based diet (a solid, low-fat, and high-carbohydrate diet) (Guevarra et al., 2019). Even when previously introduced to solid feed, it is still difficult for their enzymatic system to cope with grains and beans.
One of the consequences of the lower digestibility capacity is an increase of undigested nutrients. Harmful bacteria thrive and cause diarrhea, reducing even further an already compromised feed intake. This cycle must be broken with the support of formulations based on piglet physiology.
Post-weaning feed must support digestion and nutrient absorption, including the largest possible share possible of high-quality, digestible ingredients, with low anti-nutritional factors. High-performing feed also integrates functional amino acids, functional carbohydrates, and additives to support the intestinal mucosa and gut microbiome.
Figure 3. Supporting piglets with effective solutions
Crude protein – more of the same?
Levels of crude protein in piglet feed have been in the spotlight for quite some time. The topic can be very controversial where the exact percentage of crude protein in the final feed is concerned. Some nutritionists pragmatically recommend maximal levels of 20% in the weaner feed. Others go a bit lower, with some formulations reaching 17 to 18% total crude protein. Levels above 20% will incur high costs and may accentuate the growth of pathogenic bacteria due to a higher amount of undigested protein in the distal part of the small intestine (figure 4).
Figure 4. The dynamics of crude protein levels in piglet feed
What is not open for discussion, however, is the quality of the protein used, in terms of:
digestibility,
the total amount of anti-nutritional factors, and
the correct supply of essential and non-essential amino acids (particularly lysine, methionine, threonine, tryptophane, isoleucine and valine).
The critical role of digestibility
High-digestibility ingredients for piglets need to deliver minimum 85% digestibility. In most cases, to reach high biological values (correlating to high digestibility), these ingredients typically undergo different processing steps, including heat, physical, and chemical treatments. Animal by-products (such as hydrolyzed mucosa, fish meal, spray-dried plasma) and processed vegetable sources (soy protein concentrate, extruded grains, potato protein) can be used in high amounts during this phase. They will notably reduce the total amount of undigested protein reaching the distal part of the intestine, with 2 main benefits:
Less substrate for pathogenic bacteria proliferation (and therefore lower incidence of diarrhea)
Lower nitrogen excretion to the environment
It is common knowledge that certain storage proteins from soybean meal (for instance, glycinin and B-conglycinin) can cause damage to piglets’ intestinal morphology and trigger the activation of the immune system. However, it is normal practice to introduce this ingredient to piglets around weaning so that the animals can develop a certain level of tolerance to such compounds (Tokach et al., 2003). In Europe, where most diets are wheat-barley based, soybean meal is included in levels varying from 3 to 9% in the first 2 diets, with gradual increases during the nursery phase.
Amino acids and protein: manage the balance
When the supply and balance between essential and non-essential amino acids is concerned, reducing total crude protein brings indeed complexity to the formulations. The concept of ideal amino acid should be expanded, ideally, to all 9 essential amino acids (lysine, methionine, tryptophan, threonine, valine, isoleucine, leucine, histidine, and phenylalanine). In most cases, formulations go up to the 5th or 6th limiting amino acid. Lawor et al. (2020) suggest 2 practical approaches to avoid deficiencies when formulating low-protein piglet feed:
Maintain a maximum total lysine to crude protein ratio in the diet of 7.1 to 7.4%
Do not exceed the SID lysine to crude protein ratio of 6.4%
Some conditionally essential amino acids (e.g. arginine, proline, and glutamine) also play critical roles in diets with reduced crude protein levels. Glutamine is especially interesting. When supplemented in the feed, it can be used as a source of energy by the intestinal epithelium and, therefore, prevent atrophy and support nutrient absorption, resulting in better growth post-weaning (Hanczakowska and Niwińska, 2013; Watford et al., 2015)
The importance of the buffer capacity of the feed – supporting the enzymatic system
Given the move towards antibiotic reduction, this topic is more relevant than ever to nutritionists worldwide. The acid-binding capacity (also known as buffering capacity) of the feed directly affects the capacity of the stomach to digest protein. Hence, buffer capacity is of utmost importance in antimicrobial-free diets as it influences the growth of pathogenic bacteria (Lawlor et al., 2005).
In short, the acid-binding capacity is the resistance of an ingredient or complete feed to pH change. For piglet feed/feed ingredients, it is normally measured by the acid-binding capacity at pH4 (ABC-4). A higher ABC-4 equates to a higher buffering capacity. Feed with a high ABC-4 would require large amounts of gastric acid for the pH of the stomach to reach 4 and below. As the post-weaned piglet has limitations on producing and secreting acid, the stomach pH would stay high and, thus, less favorable for protein digestion.
The recommendation is to have a complete feed based on single ingredients with low ABC-4 values and to use additives that further reduce the ABC-4 value (such as organic acids). According to Molist (2020), post-weaning feed must have an ABC-4 that is lower than 250-300 meq/kg.
Talking about fiber
Dietary fibers are also known for regulating intestinal health in both humans and animals. Chen et al. (2020), for example, examined the effects of dietary soluble fibers (inulin) and insoluble fibers (lignocellulose) in weaned piglet diets for four weeks. Results showed that combining those fibers can positively influence nutrient digestibility, gut microbiota composition, intestinal barrier functions, and growth performance (table 1 ).
Table 1. Effects of dietary fiber supplementation on piglet growth performance (adapted from Chen et al., 2020)
How to reduce antimicrobials? Understand the roles of piglet physiology and nutrition
Swine producers might think that “How can I reduce antimicrobial use on my farm?” and “How can I improve the performance of piglets at weaning?” are two separate questions. However, that is not always the case. Answers based on a deep understanding of physiology and nutrition dynamics help piglets overcome the challenges encountered during weaning – and, thus, lessen the need for antimicrobial interventions.
In this article, we have explored the basic principles that are the basis for ensuring the performance and health of the post-weaning piglet. Although we do not have a singular solution for eliminating antimicrobials on our pig farms, we can count on a group of robust and integrated nutritional strategies. By integrating factors ranging from management to feed additives, these solutions can improve piglet health and performance throughout their lives.

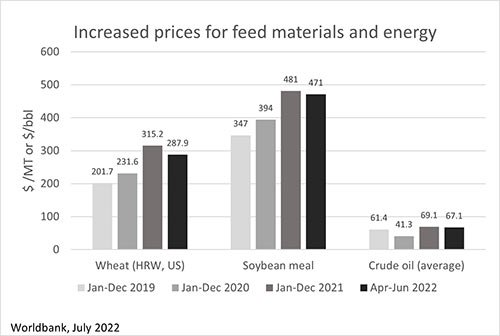 Increased prices for feed materials and energy
Increased prices for feed materials and energy 
 Several factors must be considered when using by-products in poultry nutrition
Several factors must be considered when using by-products in poultry nutrition

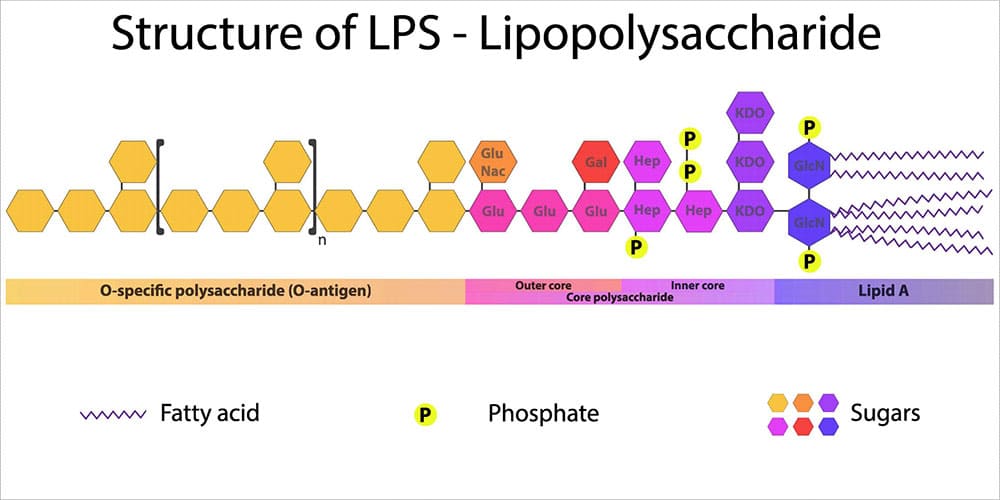

 A
A 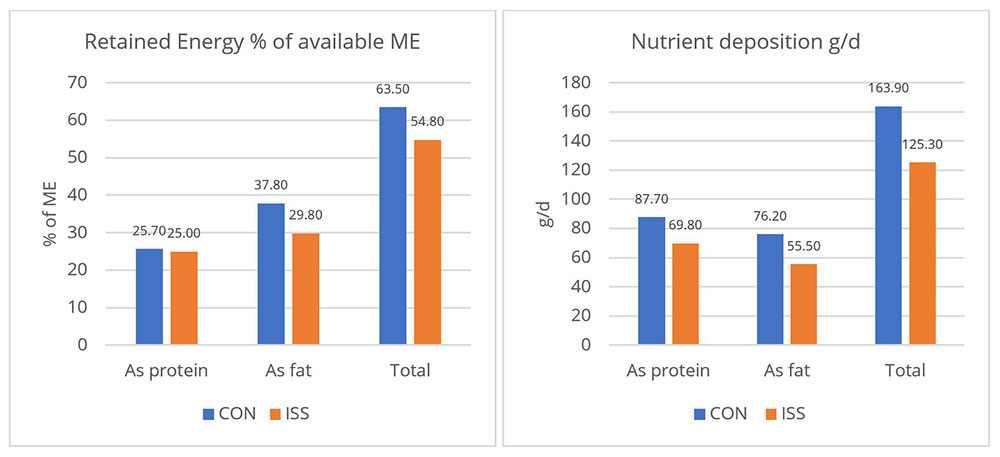

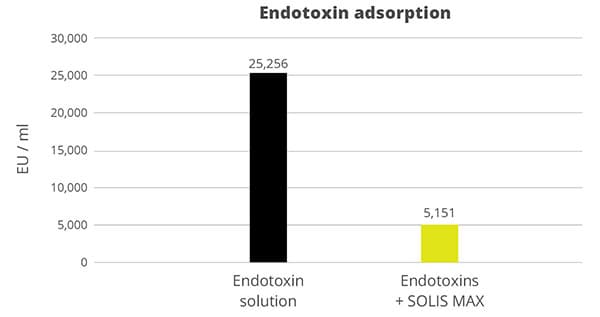



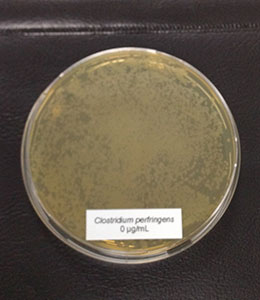
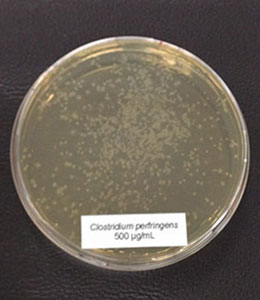
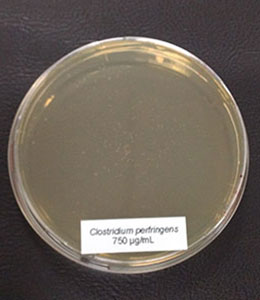

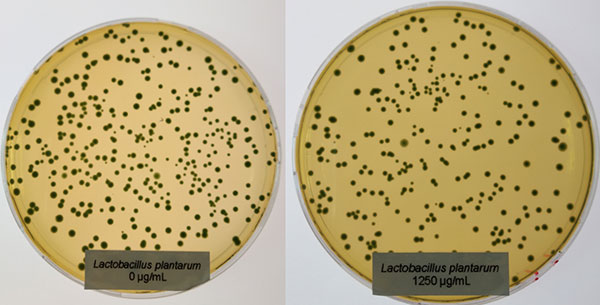
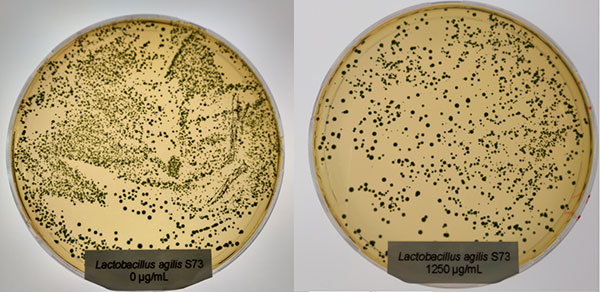
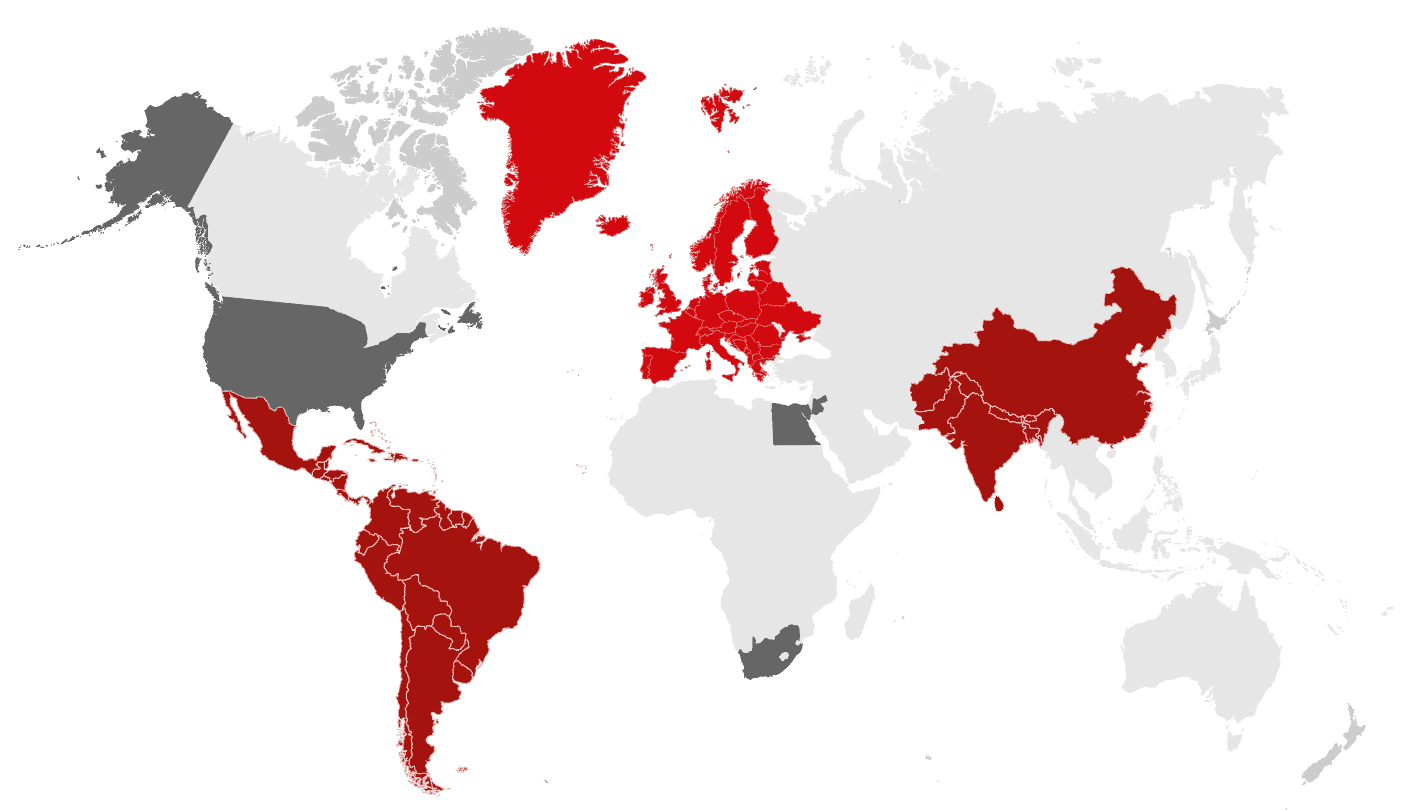
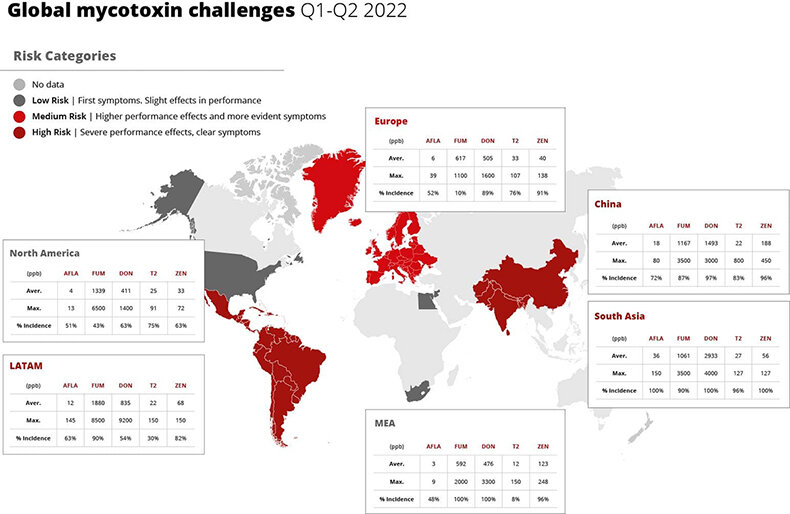


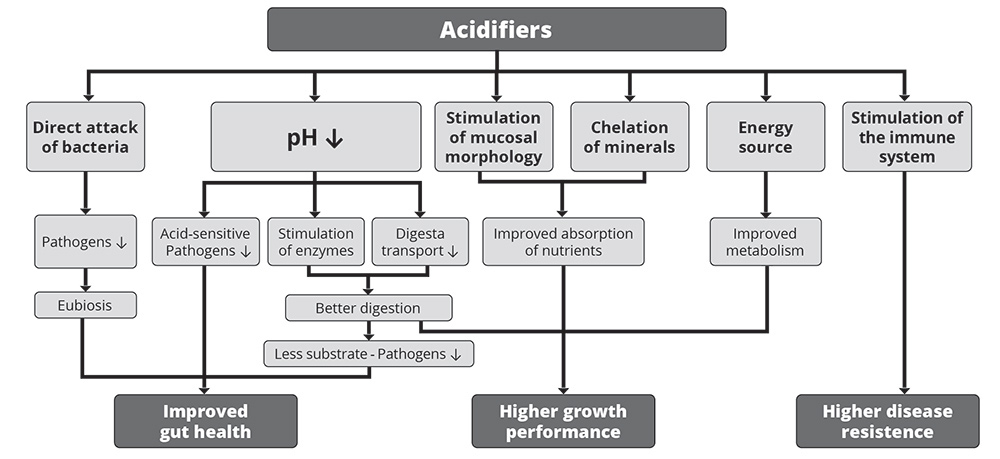

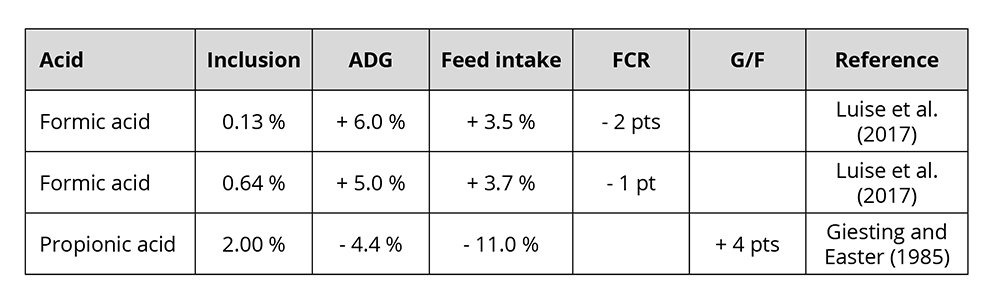



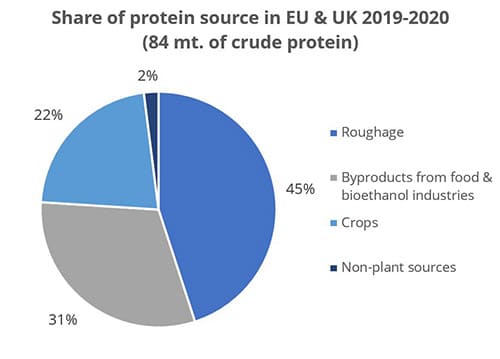
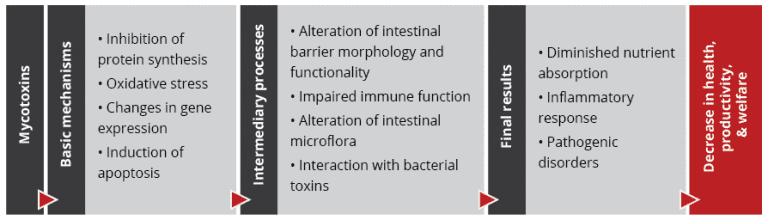
 Recent studies on the effect of various mycotoxins on the intestinal microbiota show that
Recent studies on the effect of various mycotoxins on the intestinal microbiota show that 
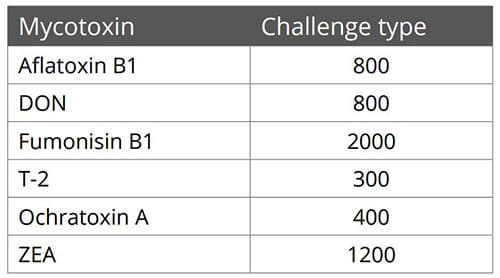
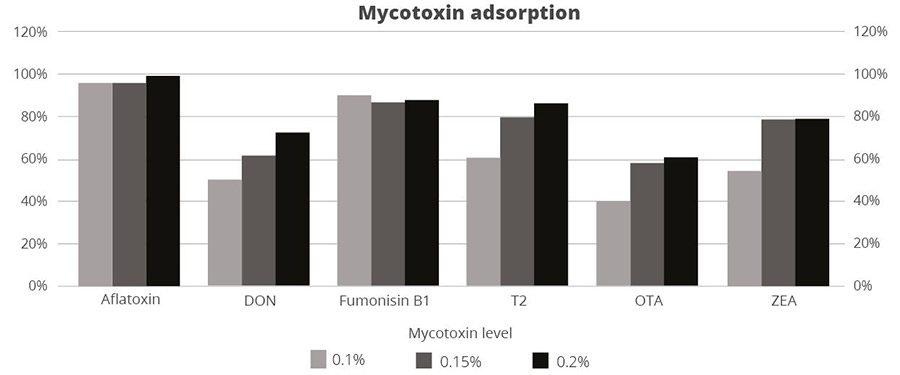


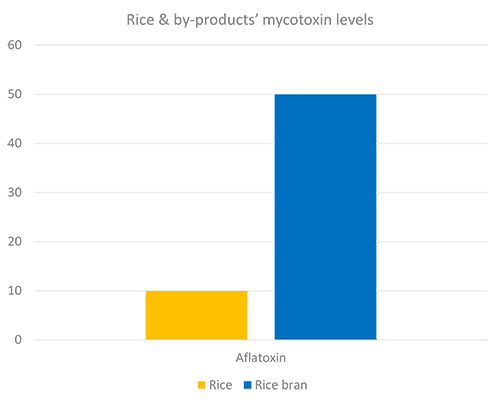
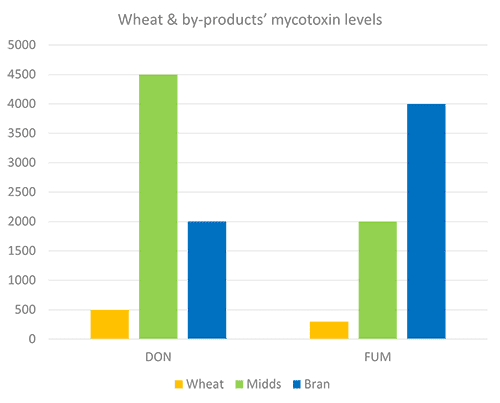
 Destillers’ dried grains with solubles (DDGS), a by-product of ethanol production, is a valuable feed ingredient, particularly as a source of protein for ruminants and monogastric animals at a competitive price.
Destillers’ dried grains with solubles (DDGS), a by-product of ethanol production, is a valuable feed ingredient, particularly as a source of protein for ruminants and monogastric animals at a competitive price.