Skuteczne fitomolekuły łączą w sobie doskonałą stabilność obróbki i silne działanie u zwierzęcia
Przez Dr. Inge Heinzl, redaktor i dr Ruturaj Patil, Global Product Manager – Phytogenics, EW Nutrition
Przez tysiąclecia rośliny były wykorzystywane do celów leczniczych w medycynie ludzkiej i weterynaryjnej oraz jako przyprawy w kuchni. Od czasu wprowadzenia zakazu stosowania antybiotykowych stymulatorów wzrostu w 2006 r. przez Unię Europejską, znalazły one również nacisk na żywienie zwierząt. Ze względu na ich właściwości trawienne, przeciwdrobnoustrojowe i promujące zdrowie jelit, wydawały się idealną alternatywą dla zrekompensowania zmniejszonego stosowania antybiotyków w krytycznych okresach, takich jak zasiedlenie, zmiana paszy lub stres związany z jelitami.
Aby zoptymalizować korzyści płynące z fitomolekuł, ważne jest, aby
Poziomy fitomolekuł są standaryzowane w celu uzyskania spójnych wyników i synergii
wykazują najwyższą stabilność podczas rygorystycznej obróbce paszy; Będąc często bardzo lotnymi substancjami, nie powinny ulegać destabilizacji w wysokich temperaturach i ciśnieniu
Fitomolekuły są korzystnie całkowicie uwolnione i dostępne u zwierzęcia, aby osiągnąć najlepszą skuteczność.
Pierwszy krok: Standaryzowane fitomolekuły
Olejki eteryczne i inne fitogeniki pochodzą z roślin. Skład roślin zasadniczo zależy od odmienności genetycznej w akcesjach, pochodzenia roślin, warunków terenowych, takich jak pogoda, gleba, zbiorowisko i czas zbiorów, ale także procesów suszenia, przechowywania i ekstrakcji próbek (Sadeh i in., 2019; Yang i in., 2018; Ehrlinger, 2007). Na przykład oliwa ekstrahowana z tymianku może zawierać od 22 do 71% odpowiedniego tymolu fenolu (Soković i in., 2009; Shabnum i Wagay, 2011; Kowalczyk i wsp., 2020).
Nowoczesna technologia umożliwia produkcję standaryzowanych fitomolekuł o najwyższym stopniu czystości i najniższej możliwej zmienności między partiami dla produktów wysokiej jakości. Oferuje również większy zrównoważony rozwój środowiskowy i ekonomiczny dzięki niezawodnej i opłacalnej technologii pozyskiwania.
Zastosowanie tak wysoce standaryzowanych fitomolekuł umożliwia produkcję fitogenicznych suplementów paszowych o niezmiennie wysokiej jakości.
Krok drugi: Wybór najbardziej odpowiednich fitomolekuł
Fitomolekuły mają różne podstawowe cechy. Niektóre wspomagają trawienie (Cho et al., 2006, Oetting, 2006; Hernandez, 2004); inne działają przeciwko patogenom (Sienkiewitz i in., 2013; Smith-Palmer i inni, 1998; Özer i in., 2007) lub są przeciwutleniaczami (Wei i Shibamoto, 2007; Cuppett i Hall, 1998). Aby zoptymalizować zdrowie jelit w produkcji zwierzęcej, jednym z głównych obiecujących mechanizmów jest redukcja patogenów przy jednoczesnym wzroście pożytecznych drobnoustrojów. Zmniejszenie patogenów w jelitach nie tylko zmniejsza ryzyko występowania zapalenia jelit, ale także eliminuje niewygodnych konkurentów dla paszy.
Aby znaleźć najlepszą kombinację służącą zamierzonemu celowi, należy najpierw ocenić dużą liczbę różnych fitomolekuł pod kątem ich struktury, właściwości chemicznych i aktywności biologicznej. Dostępność i koszt substancji to kolejne czynniki, które należy wziąć pod uwagę. Przy wyborze najbardziej odpowiednich fitomolekuł produkowane i testowane są różne mieszaniny pod kątem ich skuteczności. W tym przypadku istotne jest uwzględnienie efektów synergicznych lub antagonistycznych.
Aby uzyskać skuteczną i wydajną mieszankę fitomolekuł, konieczne jest wiele etapów selekcji i testów – w wyniku czego prawdopodobnie tylko kilka mieszanin może spełnić wymagania.
Krok trzeci: Ochrona składników
Wiele fitomolekuł jest z natury bardzo niestabilnych. Tak więc tylko standaryzowana zawartość fitogeników w produkcie nie może zapewnić pełnej dostępności fitomolekuł, gdy są stosowane w paszy dla zwierząt. Niektóre części składników mogą już zagubić się w młynie paszowym ze względu na rygorystyczny proces higienizacji paszy stosowany przez młynarzy paszowych w celu zmniejszenia obciążenia chorobotwórczego. Ogrzewanie jest poważnym wyzwaniem dla wysoce lotnych składników w produkcie na bazie fitomolekuł. Tak więc ochrona tych fitomolekuł staje się niezbędna, aby zagwarantować, że fitomolekuły wprowadzone do paszy dotrą do zwierzęcia.
Wymagana jest delikatna równowaga, aby zapewnić dostępność i aktywność fitomolekuł we właściwym miejscu w jelitach. Fitomolekuły nie mogą zostać utracone podczas przetwarzania paszy, ale muszą być również uwalniane w jelicie. Nośnik z kapilarnym wiązaniem fitomolekuł wraz z powłoką ochronną może być jednym z dostępnych skutecznych rozwiązań. Chroni składniki podczas przetwarzania paszy i zapewnia uwalnianie u zwierzęcia.
Badania wykazują doskonałą stabilność Ventar D w trudnych warunkach
Ventar D to najnowszej generacji rozwiązanie oparte na fitomolekułach do optymalizacji zdrowia jelit wprowadzone przez EW Nutrition, GmbH. Przeprowadzono badanie naukowe w celu porównania stabilności Ventar D w procesie granulowania z dwoma wiodącymi fitogenicznymi konkurencyjnymi suplementami paszowymi.
W tym badaniu pasza z różnymi dodanymi fitogenicznymi suplementami paszowymi musiała zostać poddana procesowi kondycjonowania i granulacji. Składniki aktywne analizowano przed i po procesie peletyzacji. Wszystkie testowane fitogeniczne suplementy paszowe zostały dodane do standardowej paszy dla brojlerów przy zalecanym przez producenta współczynniku włączenia. Testy przeprowadzono w czasach kondycjonowania 45, 90 i 180 sekund oraz temperaturach granulowania 70, 80 i 90 ° C (158, 176 i 194 ° F). Po schłodzeniu zebrano i przeanalizowano trzy próbki. Odpowiednią substancję znacznikową analizowano za pomocą chromatografii gazowej/spektrometrii masowej (GC/MS) w celu zmierzenia szybkości odzysku w gotowej paszy. Zawartość fitomolekuł w paszy zacieru (przed granulacją) znaleziona przez laboratorium została wykorzystana jako punkt odniesienia i ustawiona na 100% odzysku. Wskaźniki odzysku paszy granulowanej oceniano w odniesieniu do tego poziomu wyjściowego.
Wyniki przedstawiono na rysunku 1. Ventar D wykazał najwyższą stabilność składników aktywnych ze współczynnikami odzysku 90% w temperaturze 70°C/45 sek. lub 80°C/90 s i 84% w temperaturze 90°C/180 s. Nowoczesna technologia produkcji zastosowana w Ventar D zapewnia, że składniki aktywne są dobrze chronione w całym procesie granulacji.
Rysunek 1: Stabilność fitomolekuł w warunkach przetwarzania, w stosunku do linii podstawowej zacieru (100%)
Kolejną próbę przeprowadzono w wytwórni pasz w USA. Do tej próby zebrano dziesięć próbek z różnych partii paszy zacieru, w której dodano Ventar D w dawce 110 g / t. Kondycjonowanie zacieru było w temperaturze 87,8 ° C (190 ° F) przez 6 minut i 45 sekund. Po procesie granulowania pobrano dziesięć próbek z granulowanej paszy z ciągłego przepływu z 5-minutową przerwą między próbkami w celu określenia odzysku Ventar D.
Średni odzysk uzyskany dla Ventar D wyniósł 92%.
Badania wykazują poprawę wydajności wzrostu
Wstępne próby wykazały całkowite uwalnianie Ventar D w modelach trawienia. Aby zbadać korzyści w warunkach in vivo, Ventar D testowano u brojlerów z szybkością włączenia 100 g / MT.
Kilka badań in vitro wykazało aktywność przeciwdrobnoustrojową leku Ventar D. Jeden test potwierdza również, że Ventar D może wykazywać zróżnicowaną aktywność przeciwdrobnoustrojową poprzez silniejszą aktywność przeciwko powszechnym bakteriom enteropatogennym, oszczędzając jednocześnie korzystne (Heinzl, 2022). Co więcej, działanie przeciwutleniające i przeciwzapalne Ventar D wspiera lepsze funkcjonowanie bariery jelitowej. Lepsze zdrowie jelit prowadzi do wyższej wydajności wzrostu i lepszej konwersji paszy, co można wykazać w kilku badaniach z brojlerami (ryc. 2 i 3). W testach grupę otrzymującą Ventar D porównano z grupą kontrolną bez takiego suplementu paszowego lub grupami dostarczanymi z produktami konkurencyjnymi w zalecanych dawkach włączenia.
W porównaniu z negatywną grupą kontrolną, grupa Ventar D konsekwentnie wykazywała wyższy średni dzienny przyrost 0,3-4,1 g (0,5-8,5%) i o 3-4 punkty lepszą konwersję paszy. W porównaniu z produktami konkurencji, Ventar D zapewnił o 1-1,7 g (2-3%) wyższy średni dzienny przyrost i o 3 punkty lepszy / 1 punkt wyższy FCR niż konkurenci 2 i 1.
Rysunek 2: Średni przyrost dobowy (g) – wyniki kilku badań przeprowadzonych z brojlerami
Rysunek 3: FCR – wyniki kilku badań przeprowadzonych z brojlerami
Standaryzacja i nowe technologie dla wyższej rentowności
Kilka badań in vitro i in vivo dowiodło, że Ventar D przenosi “moc fitomolekuł” na wyższy poziom: Połączenie standaryzowanych fitomolekuł i optymalnej ochrony składników aktywnych prowadzi do doskonałej stabilności produktu podczas przetwarzania paszy. Większa ilość składników aktywnych docierających do jelit poprawia zdrowie jelit i zwiększa wydajność produkcyjną zwierząt. Ventar D pokazuje, w jaki sposób możemy efektywniej wykorzystywać fitomolekuły i czerpać korzyści z wyższej rentowności gospodarstwa.
References:
Cho, J. H., Y. J. Chen, B. J. Min, H. J. Kim, O. S. Kwon, K. S. Shon, I. H. Kim, S. J. Kim, and A. Asamer. “Effects of Essential Oils Supplementation on Growth Performance, IGG Concentration and Fecal Noxious Gas Concentration of Weaned Pigs”. Asian-Australasian Journal of Animal Sciences 19, no. 1 (2005): 80–85. https://doi.org/10.5713/ajas.2006.80.
Cuppett, Susan L., and Clifford A. Hall. “Antioxidant Activity of the Labiatae”. Advances in Food and Nutrition Research 42 (1998): 245–71. https://doi.org/10.1016/s1043-4526(08)60097-2.
Ehrlinger, M. “Phytogenic Additives in Animal Nutrition.” Dissertation, Veterinary Faculty of the Ludwig Maximilians University, 2007.
Heinzl, I. “Efficient Microbiome Modulation with Phytomolecules”. EW Nutrition, August 30, 2022. https://ew-nutrition.com/pushing-microbiome-in-right-direction-phytomolecules/.
Hernández, F., J. Madrid, V. García, J. Orengo, and M.D. Megías. “Influence of Two Plant Extracts on Broilers Performance, Digestibility, and Digestive Organ Size.” Poultry Science 83, no. 2 (2004): 169–74. https://doi.org/10.1093/ps/83.2.169.
Kowalczyk, Adam, Martyna Przychodna, Sylwia Sopata, Agnieszka Bodalska, and Izabela Fecka. “Thymol and Thyme Essential Oil—New Insights into Selected Therapeutic Applications.” Molecules 25, no. 18 (2020): 4125. https://doi.org/10.3390/molecules25184125.
Lindner, , U. “Aromatic Plants – Cultivation and Use.” Düsseldorf: Teaching and Research Institute for Horticulture Auweiler-Friesdorf, 1987.
Oetting, Liliana Lotufo, Carlos Eduardo Utiyama, Pedro Agostinho Giani, Urbano dos Ruiz, and Valdomiro Shigueru Miyada. “Efeitos De Extratos Vegetais e Antimicrobianos Sobre a Digestibilidade Aparente, O Desempenho, a Morfometria Dos Órgãos e a Histologia Intestinal De Leitões Recém-Desmamados.” Revista Brasileira de Zootecnia 35, no. 4 (2006): 1389–97. https://doi.org/10.1590/s1516-35982006000500019.
Sadeh, Dganit, Nadav Nitzan, David Chaimovitsh, Alona Shachter, Murad Ghanim, and Nativ Dudai. “Interactive Effects of Genotype, Seasonality and Extraction Method on Chemical Compositions and Yield of Essential Oil from Rosemary (Rosmarinus Officinalis L”.).” Industrial Crops and Products 138 (2019): 111419. https://doi.org/10.1016/j.indcrop.2019.05.068.
Shabnum, Shazia, and Muzafar G. Wagay. “Essential Oil Composition of Thymus Vulgaris L. and Their Uses”. Journal of Research & Development 11 (2011): 83–94.
Sienkiewicz, Monika, Monika Łysakowska, Marta Pastuszka, Wojciech Bienias, and Edward Kowalczyk. “The Potential of Use Basil and Rosemary Essential Oils as Effective Antibacterial Agents.” Molecules 18, no. 8 (2013): 9334–51. https://doi.org/10.3390/molecules18089334.
Smith-Palmer, A., J. Stewart, and L. Fyfe. “Antimicrobial Properties of Plant Essential Oils and Essences against Five Important Food-Borne Pathogens”. Letters in Applied Microbiology 26, no. 2 (1998): 118–22. https://doi.org/10.1046/j.1472-765x.1998.00303.x.
Soković, Marina, Jelena Vukojević, Petar Marin, Dejan Brkić, Vlatka Vajs, and Leo Van Griensven. “Chemical Composition of Essential Oils of Thymus and Mentha Species and Their Antifungal Activities”. Molecules 14, no. 1 (2009): 238–49. https://doi.org/10.3390/molecules14010238.
Wei, Alfreda, and Takayuki Shibamoto. “Antioxidant Activities and Volatile Constituents of Various Essential Oils.” Journal of Agricultural and Food Chemistry 55, no. 5 (2007): 1737–42. https://doi.org/10.1021/jf062959x.
Yang, Li, Kui-Shan Wen, Xiao Ruan, Ying-Xian Zhao, Feng Wei, and Qiang Wang. “Response of Plant Secondary Metabolites to Environmental Factors”. Molecules 23, no. 4 (2018): 762. https://doi.org/10.3390/molecules23040762.
Özer, Hakan, Münevver Sökmen, Medine Güllüce, Ahmet Adigüzel, Fikrettin Şahin, Atalay Sökmen, Hamdullah Kiliç, and Özlem Bariş. “Chemical Composition and Antimicrobial and Antioxidant Activities of the Essential Oil and Methanol Extract of Hippomarathrum Microcarpum (Bieb.) from Turkey”. Journal of Agricultural and Food Chemistry 55, no. 3 (2007): 937–42. https://doi.org/10.1021/jf0624244.
EW Nutrition wprowadza na rynek Ventar D, optymalizator zdrowia jelit nowej generacji
VISBEK, 18 października – EW Nutrition ogłasza wprowadzenie na rynek najlepszego w swojej klasie modyfikatora zdrowia jelit nowej generacji. Ventar D to innowacyjna, zastrzeżona mieszanka fitomolekuł z nowatorskim mechanizmem dostarczania.
Ventar D odpowiada na kluczowe wymagania branży żywienia zwierząt. Produkt został opracowany, aby wspierać zdrowie jelit i poprawiać wydajność, co skutkuje wzrostem rentowności dla klienta. Ventar D jest wynikiem zintegrowanego, wspólnego wysiłku zespołów badawczych, rozwoju, produkcji, sprzedaży i usług firmy EW Nutrition.
Michael Gerrits, dyrektor zarządzający EW Nutrition, podkreśla sukces wewnętrznych, holistycznych procesów badawczo-rozwojowych firmy: „EW Nutrition angażuje się w dostarczanie najwyższej klasy rozwiązań w zakresie zdrowia jelit, aby zmniejszyć zależność branży żywienia zwierząt od antybiotyków. Począwszy od dogłębnego zrozumienia potrzeb klientów, 100% zintegrowane podejście wsteczne pozwala na bezproblemowe wsparcie ze strony EW Nutrition podczas całej podróży klienta.”
Ruturaj Patil, Menedżer Produktu EW Nutrition Ventar D, mówi o korzyściach, jakie Ventar D przynosi swoim klientom: „Skuteczność każdego skutecznego rozwiązania dla zdrowia jelit polega na jego składzie, stabilności i dostarczaniu do przewodu pokarmowego. Ventar D oferuje opatentowaną formułę, najlepszą w swojej klasie stabilność granulowania i innowacyjny system dostarczania. Cieszymy się, że możemy zaoferować naszym klientom to nowatorskie rozwiązanie i być częścią ich podróży, aby uczynić produkcję zwierzęcą bardziej zrównoważoną, przy jednoczesnym zwiększeniu rentowności”.
EW Nutrition oferuje rozwiązania w zakresie żywienia zwierząt dla przemysłu paszowego. Firma koncentruje się na zdrowiu jelit, wspieranym przez inne linie produktów. Firma EW Nutrition bada, rozwija, produkuje, sprzedaje i serwisuje większość sprzedawanych przez siebie produktów. W 50 krajach kluczowi klienci są obsługiwani bezpośrednio przez własny personel EW Nutrition.
Martwicze zapalenie jelit: Pełny przegląd
Inge Heinzl, Marisabel Caballero, Ajay Bhoyar
Eliminacja martwiczego zapalenia jelit z hodowli zaczyna się od dobrego zrozumienia, co to jest, jak temu zapobiegać i jak złagodzić jego wpływ na produkcję drobiu.
Martwicze zapalenie jelit jest chorobą drobiu spowodowaną przerostem Clostridium perfringens typu A i w mniejszym stopniu typu C w jelicie cienkim. Toksyny wytwarzane przez C. perfringens uszkadzają również ścianę jelita. Na ogół występuje u brojlerów w wieku 2-6 tygodni. W postaciach subklinicznych charakteryzuje się zaburzeniami trawienia. Postacie kliniczne prowadzą do poważnych problemów i zwiększonej śmiertelności w stadach w bardzo krótkim czasie.
Martwicze zapalenie jelit jest przyczyną 6 miliardów dolarów strat w światowej produkcji drobiu, a ta dająca się kontrolować choroba, rośnie. Jednym z powodów jest dobrowolne lub wymagane prawem ograniczenie antybiotyków w produkcji zwierzęcej. Tendencja ta jest napędzana przez coraz częstsze występowanie oporności na środki przeciwdrobnoustrojowe, a także zapotrzebowanie konsumentów. Innym powodem jest redukcja jonoforów, które oprócz działania przeciwko kokcydiom, wykazują również skuteczność przeciwko Clostridia. Gdy stosuje się żywe szczepionki przeciw kokcydiozie, zastosowanie tych jonoforów nie jest możliwe i zwiększa się liczba Clostridia / martwiczego zapalenia jelit (Williams, 2005).
Chociaż jest to powszechny problem w przypadku wszystkich rodzajów drobiu, w szczególności w przypadku brojlerów, martwicowe zapalenie jelit i kokcydioza są najważniejszymi problemami zdrowotnymi.
Kliniczne i podkliniczne postacie ne
Postać kliniczna
(c) Rob Moore
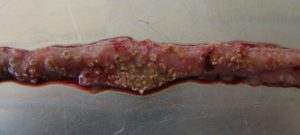
… Charakteryzuje się ostrą, ciemną biegunką powodującą mokrą ściółkę i nagle zwiększającą się śmiertelność stada nawet o 1% dziennie po pojawieniu się pierwszych objawów klinicznych (Ducatelle i Van Immerseel, 2010), czasami sumującą się do śmiertelności 50% (Van der Sluis, 2013). Ptaki mają potargane pióra, brak apetytu i są w letargu.
Sekcja zazwyczaj pokazuje balonowe jelito cienkie z szorstką powierzchnią błony śluzowej, zmianami i brązowawymi (błoniczymi) pseudobłonami. Podczas sekcji zwłok występuje dużo wodnistego brązowego, zabarwionego krwią płynu i nieprzyjemny zapach. Wątroba jest ciemna, opuchnięta i jędrna, a woreczek żółciowy rozdęty (Hofacre i wsp., 2018).
W przypadku nadostrego martwiczego zapalenia jelit ptaki mogą umrzeć bez żadnych objawów wstępnych.
Postać subkliniczna
Kiedy ptaki cierpią na postać subkliniczną, chroniczne uszkodzenie błony śluzowej jelit i zwiększoną ilość śluzu w jelicie cienkim, prowadzi to do upośledzenia trawienia i wchłaniania składników odżywczych, co prowadzi do słabej wydajności wzrostu.
Pogorszenie konwersji paszy i wynikająca z tego obniżona wydajność stają się zauważalne około 35 dnia życia. Ponieważ pasza stanowi około 65-75% kosztów produkcji brojlerów, słaba konwersja paszy zwiększa koszty produkcji i znacząco wpływa na rentowność. Często ze względu na brak wyraźnych objawów ta subkliniczna choroba pozostaje nieleczona i trwale wpływa na wydajność produkcji.
Patogeny
Za martwicę odpowiedzialne są Gram-dodatnie bakterie beztlenowe, specyficzne szczepyClostridium perfringenstypu A oraz w mniejszym stopniu typ C (Keyburn et al., 2008).
Clostridia występują głównie w glebie, w której rozkładają się substancje organiczne, w ściekach oraz w przewodzie pokarmowym zwierząt i ludzi. Bakterie te wytwarzają przetrwalniki, które są niezwykle odporne na wpływ środowiska (ciepło, napromieniowanie, eksykkacja) oraz niektóre środki dezynfekujące i mogą przetrwać kilka lat. W odpowiednich warunkach zarodniki C. perfringens mogą nawet rozmnażać się w paszy lub ściółce.
Clostridium perfringens to naturalny mieszkaniec jelita kur. U zdrowych ptaków, występuje w wielu różnych szczepów na 10 -10 42 CFU / g treści pokarmowej (McDevitt’a i wsp., 2006). Choroba zaczyna się, gdy C. perfringens namnaża się w jelicie cienkim, zwykle z powodu połączenia czynników, takich jak: duża ilość białka, niska odporność i brak równowagi we florze jelitowej. Następnie, ilość zwiększa się do 10 -10 79 CFU / g treści pokarmowej (Dahiya i wsp., 2005).
NetB, kluczowy czynnik zjadliwości dla NE
Aby zadomowić się w gospodarzu, Clostridium Spp. a inne patogeny zależą od czynników wirulencji (patrz infobox). Te czynniki zjadliwości obejmują na przykład „narzędzia” do przywiązywania się, unikania lub tłumienia układu odpornościowego gospodarza, „narzędzia” do pozyskiwania składników odżywczych i „narzędzia” do wchodzenia do komórek jelitowych. Przez lata zakładano, że toksyna α wytwarzana przez C. perfringens jest zaangażowana w rozwój choroby i kluczowy czynnik zjadliwości. W 2008 roku Keyburn i współpracownicy odkryli inny kluczowy czynnik zjadliwości, używając mutanta C. perfringens niezdolnego do wytwarzania toksyny α, ale nadal powodującego martwicze zapalenie jelit.
W ten sposób zidentyfikowano inną toksynę występującą tylko u kurczaków cierpiących na martwicze zapalenie jelit: C. perfringens toksyna podobna do martwiczego zapalenia jelit typu B (NetB). NetB to toksyna tworząca pory. Toksyny tworzące pory to egzotoksyny zwykle wytwarzane przez bakterie chorobotwórcze, ale mogą być również wytwarzane przez inne mikroorganizmy. Te toksyny niszczą integralność błon komórkowych ściany jelita. Wyciekająca zawartość komórek służy jako składniki odżywcze dla bakterii. Jeśli komórki odpornościowe zostaną zniszczone, może to częściowo wpłynąć na reakcję immunologiczną (Los i in., 2013).
Dodatkowo patogenne szczepy C. perfringens wytwarzają bakteriocyny – najważniejszą z nich jest Perfrin (Timbermont et al., 2014) – do zahamowania proliferacji nieszkodliwych Clostridium Spp. szczepów i zastąpienia normalnej flory jelitowej kurczaków (Riaz et al., 2017).
Przykłady czynników zjadliwości
Adhezyny
Umożliwiają patogenowi przyleganie lub przyczepianie się w docelowym miejscu gospodarza, np. przez fimbrię. Pili umożliwiają wymianę RNA lub DNA między patogenami.
Czynniki inwazji
Ułatwiają penetrację i dystrybucję patogenów w tkance żywiciela (inwazja i rozprzestrzenianie się enzymów). Na przykład: hialuronidaza atakująca kwas hialuronowy tkanki łącznej lub wici, umożliwiająca czynny ruch patogenów.
Toksyny
Uszkadzają lub niszczą komórki gospodarza (np. Endotoksyny – lipopolisacharydy, egzotoksyny)
Strategie uników
Umożliwiają patogenowi poddanie się strategiom obronnym układu odpornościowego (np. czynniki antyfagocytozy zapewniają ochronę przed atakiem fagocytów, specyficzne przeciwciała są inaktywowane przez enzymy).
Kurczak o optymalnym zdrowiu jelit może być mniej podatny na NE. Dodatkowe czynniki predysponujące są niezbędne do wydzielenia składników odżywczych i dostosowania środowiska jelitowego do proliferacji tych patogenów, umożliwiając im wywoływanie chorób (Van Immerseel i in., 2008; Williams, 2005).
Czynniki predysponujące
Pasza: skład i wielkość cząstek
Nie należy lekceważyć roli paszy w rozwoju martwiczego zapalenia jelit. Tutaj do gry wchodzą substancje tworzące środowisko jelitowe sprzyjające C. perfringens.
Zanieczyszczenie mykotoksynami
Mikotoksyny szkodzą integralności jelit i stwarzają idealne warunki do namnażania sięClostridium perfringens.
Mikotoksyny nie mają bezpośredniego wpływu na proliferację C. perfringens, produkcję toksyn ani transkrypcję NetB. Jednak mykotoksyny zakłócają integralność zdrowotną jelit, tworząc sprzyjające środowisko dla patogenu. Na przykład:
DON zapewnia dobre warunki do proliferacji perfringens poprzez przerwanie bariery jelitowej i uszkodzenie nabłonka. Prawdopodobnie wynikająca z tego przepuszczalność nabłonka i zmniejszone wchłanianie białek pokarmowych mogą prowadzić do większej ilości białek w jelicie cienkim. Białka te mogą służyć jako składniki odżywcze dla patogenu (Antonissen et al., 2014).
DON i inne mykotoksyny zmniejszają liczbę bakterii wytwarzających kwas mlekowy, wskazując na zmianę równowagi mikrobiologicznej (Antonissen i in., 2016.).
Eimeria ssp.
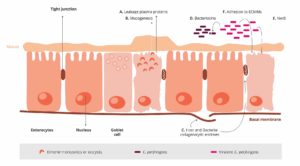
Nienaruszony nabłonek jelitowy jest najlepszą obroną przed potencjalnymi patogenami, takimi jak C. perfringens. Tutaj pojawia się kokcydioza. Moore (2016) wykazał, że przez uszkodzenie nabłonka jelita, gatunki Eimeria dają C. perfringens dostęp do podstawowych domen nabłonka śluzówki jelit. Następnie następuje pierwsza faza procesu patologicznego i stamtąd C. perfringens atakuje blaszkę właściwą. Następuje uszkodzenie nabłonka (Olkowski i in., 2008). Białka osocza wyciekające do jelit i wytwarzany śluz są bogatymi źródłami składników odżywczych (Van Immerseel i in., 2004; Collier i in., 2008). Kolejnym skutkiem kokcydiozy jest zmiana równowagi mikrobiologicznej w jelitach poprzez zmniejszenie liczby np. Candidatus savagella, który aktywuje wrodzoną obronę immunologiczną.
Eimeria wywołuje wyciek białek osocza poprzez zabijanie komórek nabłonka
Wzmacniają produkcję śluzu w jelicie
1 + 2 prowadzą do zwiększenia ilości dostępnych składników odżywczych i tworzą środowisko sprzyjające namnażaniu się C. perfringens
Nie tylko Eimeria Spp., Ale także inne patogeny (np. Salmonella Spp., Larwy Ascarid, wirusy) i czynniki, takie jak mykotoksyny uszkadzające błonę śluzową jelit, mogą utorować drogę do zakażenia C. perfringens. Czynniki predysponujące, takie jak mokra ściółka, której wilgoć jest niezbędna do zarodnikowania Eimeria Spp. oocysty należy również wziąć pod uwagę jako czynniki sprzyjające martwicowemu zapaleniu jelit (Williams, 2005).
Czynniki immunosupresyjne
Oprócz wyjaśnionej już paszy dla influencerów, mykotoksyn i kokcydiów, należy również wspomnieć o innych czynnikach predysponujących. Ogólnie rzecz biorąc, każdy czynnik wywołujący stres u zwierząt zaburza równowagę flory jelitowej. Wynikająca z tego supresja układu odpornościowego przyczynia się do ryzyka martwiczego zapalenia jelit (Tsiouris, 2016). Czynniki te obejmują:
Bakterie : Shivaramaiah i wsp. (2011) zbadali noworodkową infekcję Salmonella typhimurium jako czynnik predysponujący do NE. Wczesna infekcja powoduje znaczne uszkodzenie jelita (Porter i wsp., 21) Dodatkowo Hassan i wsp. (1994) wykazali, że prowokacja Salmonella typhimurium negatywnie wpłynęła na rozwój limfocytów, co może również sprzyjać kolonizacji Clostridium perfringens.
Wirusy : Wiadomo, że zakaźna choroba torebek nasiennych zwiększa nasilenie infekcji salmonellą, gronkowcami, ale także clostridiami. Inne choroby wirusowe wywołujące Clostridię to Gumboro i choroba Marka.
Stres : Przewód pokarmowy jest szczególnie wrażliwy na wszelkiego rodzaju stres. Stres ten może być spowodowany np. zbyt wysokimi temperaturami, dużą gęstością hodowli, nagłą zmianą paszy.
Leczenie
W ostrych przypadkach rolnik powinien skonsultować się z lekarzem weterynarii i leczyć swoje ptaki.
Należy wspomnieć, że ponieważ obróbka odbywa się za pomocą paszy lub wody, mogą być leczone tylko ptaki, które nadal spożywają wodę lub paszę.
Antybiotyki
W leczeniu ostrej NE powszechnie stosuje się antybiotyki ukierunkowane na bakterie Gram-dodatnie. Wyboru antybiotyku dokonuje lekarz weterynarii, biorąc pod uwagę sposób działania oraz obecność genów oporności w gospodarstwie / stadzie.
Niektóre bakterie są mniej wrażliwe na niektóre antybiotyki z powodu mutacji genetycznych. Są w stanie zrobić:
stymulują produkcję enzymów, które rozkładają lub modyfikują antybiotyki i inaktywują je (1)
eliminują wejścia dla antybiotyków lub promują rozwój pomp, które dymisjonują antybiotyk przed jego działaniem (2)
zmienić lub wyeliminować cząsteczki, z którymi antybiotyk miałby się wiązać (cele dla antybiotyków).
Oznacza to, że gdy stosuje się odpowiednie antybiotyki, bakterie odporne na te antybiotyki przeżywają. Dzięki wyeliminowaniu ich konkurentów są w stanie lepiej się rozmnażać.
Dodatkowo oporność ta może być przenoszona za pomocą „genów odporności”
do komórek potomnych
poprzez ich spożycie z martwych bakterii (3)
poprzez poziomy transfer genów (4)
przez wirusy (5)
Każde zastosowanie antybiotyków sprzyja rozwojowi oporności (Robert Koch Institute, 2019). Krótkotrwałe stosowanie,lepsza bioasekuracjalub aplikacja w małych dawkach dają bakteriom większe szanse na adaptację.
Bakteriofagi
Eksperymentalne stosowanie terapii fagowych okazało się skuteczne w zmniejszaniu progresji choroby i objawów martwiczego zapalenia jelit (Miller i wsp., 2010). Poprzez doustne podanie koktajlu bakteriofagów Miller i współpracownicy mogli zmniejszyć śmiertelność o 92% u brojlerów prowokowanych C. perfringens w porównaniu z brojlerami poza kontrolą.
Mechanizm działania: endolizyny, wysoko rozwinięte enzymy wytwarzane przez bakteriofagi, są zdolne do trawienia ściany komórkowej bakterii w celu uwolnienia potomstwa faga (Fischetti, 2010). Jednak fagi nadal nie są zatwierdzane przez EFSA.
Zapobieganie
Zapobieganie chorobie jest zawsze lepsze – i bardziej opłacalne – niż jej leczenie.
Jak więc należy to zrobić?
Zapobieganie warunkom sprzyjającym proliferacji Clostridium perfringens i wzmacnianie odpowiedzi immunologicznej gospodarza zmniejsza prawdopodobieństwo wystąpienia choroby.
Oprócz wyeliminowania czynników predysponujących, głównymi celami są:
Istnieją dowody na to, że większość szczepów Clostridium wyizolowanych od ptaków cierpiących na martwicze zapalenie jelit może wywołać chorobę eksperymentalnie, podczas gdy szczepy izolowane od zdrowych ptaków nie mogą. Potwierdza to, że tylko określone szczepy są problematyczne (Ducatelle i Van Immerseel, 2010).
Dlatego niezwykle ważne jest, aby unikać wprowadzania tych patogennych szczepów do gospodarstwa.
Oddzielne ubrania, buty i urządzenia do mycia / dezynfekcji rąk w każdym kurniku
Ponad 14 dni przestoju między stadami
Specyficzne środki przeciwko kokcydiozie
Szczepionka
Według parazytologów u kurczaków występuje od 7 do 9 gatunków Eimeria, które nie zapewniają wzajemnej ochrony. Skuteczna szczepionka musi zawierać sporulowane oocysty najbardziej krytycznych patogennych gatunków Eimeria ( E. acervulina, E. maxima, E. tenella, E. necatrix i E. brunetti ). Im więcej gatunków zawartych w szczepionce, tym lepiej. Jednak jeśli nie zostaną zastosowane we właściwy sposób, szczepionki mogą być nieskuteczne lub powodować reakcje u ptaków, które mogą prowadzić do NE (Mitchell, 2017).
Środki przeciw kokcydiozie
Aby uniknąć rozwoju oporności, ważne jest naprzemienne stosowanie chemikaliów (związków syntetycznych) i jonoforów (antybiotyków polieterowych) o różnych sposobach działania.
Jonofory mają specyficzny sposób działania i zabijają oocysty, zanim zdążą zainfekować ptaki. Będąc bardzo małymi, cząsteczki jonoforu mogą być pobierane i dyfundowane do zewnętrznej błony sporozoitu. Tam zmniejsza gradient stężeń prowadząc do gromadzenia się wody w sporozoicie powodując jego pękanie.
Dieta
Minimalizacja polisacharydów nieskrobiowych (NSP) w zbożach
Aby zapobiec „żerowaniu” Clostridium perfringens, należy unikać lub przynajmniej minimalizować wysoką zawartość rozpuszczalnych w wodzie, ale niestrawnych NSP, takich jak pszenica, produkty uboczne pszenicy i jęczmień. Dodatkowo w składzie paszy powinny być zawarte ksylanazy, aby zmniejszyć szkodliwe skutki NSP i poprawić wykorzystanie energii paszy. Zamiast tych zbóż do diety można włączyć kukurydzę. Ze względu na wysoką zawartość energii i wysoką dostępność składników pokarmowych uznawana jest za doskonały składnik diet brojlerów.
Formułowanie diet / diet niskobiałkowych z wysoce przyswajalnymi aminokwasami
Karmienie dietą niskobiałkową uzupełnioną krystalicznymi aminokwasami może być korzystne dla zmniejszenia ryzyka martwiczego zapalenia jelit (Dahiya i in., 2007). Aby poprawić strawność białka, a tym samym zmniejszyć proliferację C. perfringens, do paszy można dodać proteazy.
Unikanie / minimalizowanie tłuszczów zwierzęcych w diecie
Tłuszcze zwierzęce mają tendencję do zwiększania liczby Clostridium perfringens ; w związku z tym należy je zastąpić źródłami tłuszczu roślinnego.
Forma paszy
Pod względem formy paszy Engberg i wsp. (2002) stwierdzili, że ptaki karmione granulatem wykazywały zmniejszoną liczbę Clostridium perfringens w jelicie ślepym i odbytnicy niż ptaki karmione papką. Branton i współpracownicy (1987) podali niższą śmiertelność przy podawaniu paszy mielonej na walcach (grubo zmielonej) niż w przypadku paszy mielonej w młynie.
Dodatki
Dodatki można stosować albo w celu zapobiegania namnażaniu się Clostridium perfringens, albo w celu zmiany warunków środowiskowych w taki sposób, aby zapobiec proliferacji C. perfringens.
Probiotyki
Te żywe suplementy mikrobiologiczne mogą pomóc w ustanowieniu, utrzymaniu lub przywróceniu mikroflory jelitowej.
Sposób działania:
konkurują z bakteriami chorobotwórczymi o substraty i miejsca przyczepu
wytwarzają substancje przeciwdrobnoustrojowe hamujące rozwój bakterii chorobotwórczych (Gillor et al., 2008)
wiąże i neutralizuje enterotoksyny (Mathipa i Thantsha, 2017)
promować funkcje odpornościowe gospodarza (Yang i in., 2012)
Prebiotyki
Te składniki pasz służą jako substraty do promowania korzystnych bakterii w jelicie.
Sposób działania:
D-mannoza lub fruktoza, skrobie nieulegające trawieniu przez ptaki, wybiórczo stymulują wzrost i aktywność „dobrej” flory jelitowej
Fruktooligosacharydy zmniejszają ilość perfringens i E. coli w jelitach i zwiększają różnorodność Lactobacillus Spp. (Kim i in., 2011)
Donoszono, że galaktooligosacharydy, w połączeniu z probiotykiem na bazie lactis, wybiórczo promują proliferację Bifidobacterium Spp. (Jung i in., 2008).
Kwasy organiczne
Kwasy organiczne są często stosowane w dietach zwierząt w celu poprawy zdrowia jelit.
Sposób działania:
obniżone pH sprzyja korzystnym bakteriom
kwas kaprylowy tłumi perfringens, ale także Salmonella Spp. poprzez hamowanie ich wykorzystania glukozy (Skrivanova et al., 2006)
kwas laurynowy, cytrynowy, oleinowy i linolowy oraz średniołańcuchowe kwasy tłuszczowe (C8-C14) hamują wzrost perfringens
Fitomolekuły
Fitomolekuły, znane również jako drugorzędne związki roślinne, są od wieków stosowane przeciwko patogenom. Ogólnie wiadomo, że dwie podgrupy tych substancji są skuteczne przeciwko Clostridium perfringens :
Garbniki
Wiele badań wykazało skuteczność tanin przeciwko różnym patogenom, takim jak robaki pasożytnicze, Eimeria, wirusy i bakterie
Wyciągi z kasztanowca i drzewa quebracho są skuteczne nie tylko przeciwko perfringens, ale także na jego toksyny (Elizando i in., 2010)
Aktywność tanin przeciwko Eimeria (Cejas i in., 2011) i Salmonella Sp., Dwa czynniki predysponujące do NE.
Olejki eteryczne
Ich hydrofobowość umożliwia im interakcję z lipidami błony perfringens.
Mogą wnikać w błonę bakteryjną i zakłócać jej integralność.
Zwiększa to przepuszczalność błony komórkowej dla jonów i innych małych cząsteczek, takich jak ATP, co prowadzi do zmniejszenia gradientu elektrochemicznego powyżej błony komórkowej i utraty równoważników energii komórki.
Oprócz bezpośredniego wpływu na Clostridium Spp., Wiele fitomolekuł poprawia zdrowie jelit i pomaga zapobiegać namnażaniu się Clostridium Spp. a zatem martwicze zapalenie jelit.
Wiążąc mikotoksyny, można zmniejszyć uszkodzenie nabłonka jelita lub nawet mu zapobiec, tak że nie powstają warunki do proliferacji Clostridium.
Wiązanie toksyn wytwarzanych przez Clostridium perfringens może zmniejszyć występowanie lub nasilenie zmian:
Alfa-toksyna, fosfolipaza C, hydrolizuje fosfolipidy błonowe i uszkadza erytrocyty, leukocyty, miocyty i komórki śródbłonka oraz powoduje ich lipolizę (Songer, 1996). Prowadzi to do martwicy i uszkodzenia tkanki.
Stale rosnąca tendencja do zmniejszania stosowania antybiotyków i jonoforów przyczynia się do zwiększonej częstości występowania martwiczego zapalenia jelit w produkcji drobiu.
Subkliniczna postać martwiczego zapalenia jelit na ogół pozostaje niezauważona, co skutkuje niską wydajnością paszy i jest główną przyczyną strat finansowych producentów drobiu.
Utrzymanie optymalnego zdrowia jelit jest kluczem do zapobiegania występowaniu martwiczego zapalenia jelit. W dobie wolnej od antybiotyków produkcji drobiu należy rozważyć alternatywne metody zwalczania tej chorobotwórczej bakterii, a także czynników predysponujących ją do zwalczania tej wyniszczającej choroby.
References
Annett, C.B., J. R. Viste, M. Chirino-Trejo, H. L. Classen, D. M. Middleton, and E. Simko. “Necrotic enteritis: effect of barley, wheat and corn diets on proliferation of Clostridium perfringens type A.” Avian Pathology 31 (2002): 599– 602. https://doi.org/10.1080/0307945021000024544
Antonissen G, F. Van Immerseel, F. Pasmans, R. Ducatelle, F. Haesebrouck, L. Timbermont, M. Verlinden, G.P.J. Janssens, V. Eeckhaut, M. Eeckhout, S. De Saeger, S. Hessenberger, A. Martel, and S. Croubels. “The mycotoxin deoxynivalenol predisposes for the development of Clostridium perfringens-Induced necrotic enteritis in broiler chickens. PLoS ONE 9 no. 9 (2014): e108775. https://doi.org/10.1371/journal.pone.0108775
Antonissen, G., V. Eeckhaut, K. Van Driessche, L. Onrust , F. Haesebrouck, R. Ducatelle, R.J. Moore, and F. Van Immerseel. “Microbial Shifts Associated With Necrotic enteritis.” Avian Pathol. 45 no. 3 (2016): 308-312. https://doi.org/10.1080/03079457.2016.1152625
Branton, S.L., F.N. Reece, and W.M. Hagler. “Influence of a wheat diet on mortality of broiler chickens associated with necrotic enteritis.” Poultry Sci. 66 (1987): 1326-1330. https://doi.org/10.3382/ps.0661326
Cejas, E., S. Pinto, F. Prosdócimo, M. Batalle, H. Barrios, G. Tellez, and M. De Franceschi. “Evaluation of quebracho red wood (Schinopsis lorentzii) polyphenols vegetable extract for the reduction of coccidiosis in broiler chicks.” International Journal of Poultry Science 10 no. 5 (2011): 344–349. https://doi.org/10.3923/ijps.2011.344.349
Collier, C.T., C.L. Hofacre, A.M. Payne, D.B. Anderson, P. Kaiser, R.I. Mackie, and H.R. Gaskins. “Coccidia-induced mucogenesis promotes the onset of necrotic enteritis by supporting Clostridium perfringens growth.” Veterinary Immunology and Immunopathology 122 (2008):104–115.
Dahiya, J.P., D. Hoehler, A.G. Van Kessel, and M.D. Drew. “Effect of different dietary methionine sources on intestinal microbial populations in broiler chickens.” Poultry Science 86 (2007):2358–2366
Dahiya, J.P., D. Hoehler, D.C. Wilkie, A.G. van Kessel, and M.D. Drew. “Dietary glycine concentration affects intestinal Clostridium perfringens and Lactobacilli populations in broiler chickens.” Poultry Science 84 no.12 (2005):1875-85. https://doi.org/10.1093/ps/84.12.1875
Diaz Carrasco, J.M., L.M. Redondo, E.A. Redondo, J.E. Dominguez, A.P. Chacana, and M.E. Fernandez Miyakawa. “Use of plant extracts as an effective manner to control Clostridium perfringens induced necrotic enteritis in poultry.” BioMed Research International (2016): Article ID 3278359. https://dx.doi.org/10.1155/2016/3278359
Ducatelle, R. and F. van Immerseel. “Necrotic enteritis: emerging problem in broilers.” WATTAgNet.com – Poultry Health and Disease (April 9, 2010).
Elizondo, A.M., E.C. Mercado, B.C. Rabinovitz, and M.E. Fernandez-Miyakawa. “Effect of tannins on the in vitro growth of Clostridium perfringens.” Veterinary Microbiology 145 no. 3-4 (2010): 308–314. https://doi.org/10.1016/j.vetmic.2010.04.003
Engberg, R.M., M.S. Hedemann, and B.B. Jensen. “The influence of grinding and pelleting of feed on the microbial composition and activity in the digestive tract of broiler chickens.” · British Poultry Science 43 no. 4 (2002):569-579. https://doi.org/10.1080/0007166022000004480
Fischetti, V.A. “Bacteriophage endolysins: A novel anti-infective to control Gram-positive pathogens.” J Med Microbiol. 300 no. 6 (2010): 357–362. https://doi.org/10.1016/j.ijmm.2010.04.002
Gillor, O., A. Etzion and M.A. Riley. “The dual role of bacteriocins as anti- and probiotics.” Appl Microbiol Biotechnol. 81 no. 4 (2008): 591–606. https://doi.org/10.1007/s00253-008-1726-5
Hassan, J. O., and R. Curtiss III. “Virulent Salmonella typhimurium induced lymphocyte depletion and immunosuppression in chickens.” Infect. Immun. 62 (1994):2027–2036 https://doi.org/10.1128/IAI.62.5.2027-2036.1994
Hofacre, C.L., J.A. Smith, and G.F. Mathis. “Invited Review. An optimist’s view on limiting necrotic enteritis and maintaining broiler gut health and performance in today’s marketing, food safety, and regulatory climate.” Poultry Science 97 (2018):1929–1933. https://dx.doi.org/10.3382/ps/pey082
Jung, S.J., R. Houde, B. Baurhoo, X. Zhao, and B. H. Lee. “Effects of galacto-oligosaccharides and a bifidobacteria lactis-based probiotic strain on the growth performance and fecal microflora of broiler chickens.” Poultry Science 87 (2008):1694–1699. https://doi.org/10.3382/ps.2007-00489
Kaldhusdal and Skjerve. “Association between cereal contents in the diet and incidence of necrotic enteritis in broiler chickens in Norway.” Preventive Veterinary Medicine 28 (1996):1-16. https://doi.org/10.1016/0167-5877(96)01021-5
Keyburn, A. L., S. A. Sheedy, M. E. Ford, M. M. Williamson, M. M. Awad, J. I. Rood, and R. J. Moore. “Alpha-toxin of Clostridium perfringens is not an essential virulence factor in necrotic enteritis in chickens.” Infect. Immun. 74 (2006): 6496–6500. https://doi.org/10.1128/IAI.00806-06
Keyburn, A.L., J.D. Boyce, P. Vaz, T.L. Bannam, M.E. Ford, D. Parker, A. Di Rubbo, J.I. Rood, and R.J. Moore. “NetB, a new toxin that is associated with avian necrotic enteritis caused by Clostridium perfringens.” PLoS Pathog 4 no. 2, e26 (2008): 0001-0011. https://doi.org/10.1371/journal.ppat.0040026
Kim, G.-B., Y. M. Seo , C. H. Kim , and I. K. Paik. “Effect of dietary prebiotic supplementation on the performance, intestinal microflora, and immune response of broilers.” Poultry Science 90 (2011):75–82. https://doi.org/10.3382/ps.2010-00732
Knap, I., B. Lund, A. B. Kehlet, C. Hofacre, and G. Mathis. “Bacillus licheniformis prevents necrotic enteritis in broiler chickens.” Avian Diseases 54 no. 2 (2010):931-935. https://doi.org/10.1637/9106-101509-ResNote.1
Knarreborg, A., M.A. Simon, R.M. Engberg, B.B. Jensen, and G.W. Tannock. “Effects of Dietary Fat Source and Subtherapeutic Levels of Antibioticon the Bacterial Community in the Ileum of Broiler Chickensat Various Ages.” Applied and Environmental Microbiology 68 no. 12 (2002): 5918-5924. https://doi.org/0.1128/AEM.68.12.5918–5924.2002
Kocher, A. and M. Choct. “Improving broiler chicken performance. The efficacy of organic acids, prebiotics and enzymes in controlling necrotic enteritis.” Australian Government-Rural Industries Research and Development Corporation. Publ. no. 08/149 (2008).
Kondo, F. “In vitro lecithinase activity and sensitivity to 22 antimicrobial agents of Clostridium perfringens isolated from necrotic enteritis of broiler chickens.” Research in veterinary Science 45 (1988): 337-340. https://doi.org/10.1016/S0034-5288(18)30961-5
Kubena, L.F., J.A. Byrd, C.R. Young, and D.E. Corrier. “Effects of tannic acid on cecal volatile fatty acids and susceptibility to Salmonella typhimurium colonization in broiler chicks.” Poultry Science 80, no. 9 (2001): 1293–1298. https://doi.org/10.1093/ps/80.9.1293
Los, F.C.O., T.M. Randis, R.V. Aroian, and A.J. Ratner. “Role of pore-forming toxins in bacterial infectious diseases.” Microbiology and Molecular Biology Reviews 77 (2013): 173-207 https://doi.org/10.1128/MMBR.00052-12
M’Sadeq S.A., Shubiao Wu, Robert A. Swick, Mingan Choct. “Towards the control of necrotic enteritis in broiler chickens with in-feed antibiotics phasing-out worldwide.” Animal Nutrition 1 (2015): 1-11. https://dx.doi.org/10.1016/j.aninu.2015.02.004
Mathipa, M.G. and M.S. Thantsha. “Probiotic engineering: towards development of robust probiotic strains with enhanced functional properties and for targeted control of enteric pathogens.” Gut Pathog. 9 no. 28 (2017). https://doi.org/10.1186/s13099-017-0178-9
McDevitt, R.M., J.D. Brooker, T. Acamovic, and N.H.C. Sparks. “Necrotic enteritis, a continuing challenge for the poultry industry.” World’s Poultry Science Journal 62; World’s Poultry Science Association (June 2006). https://doi.org/10.1079/WPS200593
Miller, R.W., J. Skinner, A. Sulakvelidze, G.F. Mathis, and C.L. Hofacre. “Bacteriophage therapy for control of necrotic enteritis of broiler chickens experimentally infected with Clostridium perfringens.” Avian Diseases 54 no. 1 (2010): 33-40. https://doi.org/10.1637/8953-060509-Reg.1
Mitsch, P., K. Zitterl-Eglseer, B. Köhler, C. Gabler, R. Losa, and I. Zimpernik. “The Effect of Two Different Blends of Essential Oil Components on the Proliferation of Clostridium perfringens in the Intestines of Broiler Chickens.” Poultry Science 83 (2004):669–675. https://doi.org/10.1093/ps/83.4.669
Olkowski, A.A., C. Wojnarowicz, M. Chirino-Trejo, B. Laarveld, and G. Sawicki. “Sub-clinical necrotic enteritis in broiler chickens: Novel etiological consideration based on ultra-structural and molecular changes in the intestinal tissue.” Veterinary Science 85 (2008): 543–553. https://doi.org/10.1016/j.rvsc.2008.02.007
Pan, D. and Z. Yu. “Intestinal microbiome of poultry and its interaction with host and diet.” Gut Microbes 5 no. 1 (2014): 108–119. https://dx.doi.org/10.4161/gmic.26945
Rougière, N. and B. Carré. “Comparison of gastrointestinal transit times between chickens from D + and D- genetic lines selected for divergent digestion efficiency.” Animal 4 no. 11 (2010): 1861-1872. https://doi.org/10.1017/S1751731110001266
Santos, F.B.O., B.W. Sheldon, A.A. Santos Jr., and P.R. Ferket. ”Influence of housing system, grain type, and particle size on Salmonella colonization and shedding of broilers fed triticale or corn-soybean meal diets.” Poultry Science 87 (2008): 405-420. https://dx.doi.org/10.3382/ps.2006-00417
Schiavone, A. , K. Guo, S. Tassone, L .Gasco, E. Hernandez, R. Denti, and I. Zoccarato. “Effects of a Natural Extract of Chestnut Wood on Digestibility, Performance Traits, and Nitrogen Balance of Broiler Chicks.” Poult Sci. 87 no. 3 (2008): 521-527. https://doi.org/10.3382/ps.2007-00113
Shivaramaiah, S., R. E. Wolfenden, J. R. Barta, M. J. Morgan, A. D. Wolfenden, B. M. Hargis, and G. Téllez. „The role of an early Salmonella typhimurium infection as a predisposing factor for necrotic enteritis in a laboratory challenge model.” Avian Diseases 55 (2011): 319-323. https://doi.org/10.1637/9604-112910-ResNote.1
Singh, Y., V. Ravindran, T.J. Wester, A.L. Molan, and G. Ravindran. “ Influence of feeding coarse corn on performance, nutrient utilization, digestive tract measurements, carcass characteristics, and cecal microflora counts of broilers.” Poultry Science 93 (2014): 607–616. https://dx.doi.org/10.3382/ps.2013-03542
Skrivanova, E., M. Marounek, V. Benda, and P. Brezina. “Susceptibility of Escherichia coli, Salmonella sp. and Clostridium perfringens to organic acids and monolaurin.” Veterinarni Medicina 51 no. 3 (2006): 81–88. https://doi.org/10.17221/5524-VETMED
Stanley D., Wu S.-B., Rodgers N., Swick R.A., and Moore R.J. “Differential Responses of Cecal Microbiota to Fishmeal, Eimeria and Clostridium perfringens in a Necrotic Enteritis Challenge Model in Chickens.” PLoS ONE 9 no. 8 (2014): e104739. https://doi.org/10.1371/journal.pone.0104739
Tan, L., D. Rong, Y. Yang, and B. Zhang. “Effect of Oxidized Soybean Oils on Oxidative Status and Intestinal Barrier Function in Broiler Chickens.” Brazilian Journal of Poultry Science 18 no. 2 (2018): 333-342. http://dx.doi.org/10.1590/1806-9061-2017-0610
Tan, L., D. Rong, Y. Yang, and B. Zhang. “The Effect of Oxidized Fish Oils on Growth Performance, Oxidative Status, and Intestinal Barrier Function in Broiler Chickens.” J. Appl. Poult. Res. 28 (2019): 31-41. http://dx.doi.org/10.3382/japr/pfy013
Timbermont L., A. Lanckriet, J. Dewulf, N. Nollet, K. Schwarzer, F. Haesebrouck, R. Ducatelle, and F. Van Immerseel. “Control of Clostridium perfringens-induced necrotic enteritis in broilers by target-released butyric acid, fatty acids and essential oils.” Avian Pathol. 39 no. 2 (2010): 117-21. https://doi.org/10.1080/03079451003610586
Tsiouris, V. “Poultry management: a useful tool for the control of necrotic enteritis in poultry.” Avian Pathol. 45 no. 3 (2016):323-325. https://doi.org/10.1080/03079457.2016.1154502
Van der Most, P.J., B. de Jong, H.K. Parmentier and S. Verhulst. “Trade-off between growth and immune function: a meta-analysis of selection experiments.” Functional Ecology 25 (2011): 74-80. https://doi.org/0.1111/j.1365-2435.2010.01800.x
Van der Sluis, W. “Clostridial enteritis is an often underestimated problem.” Worlds Poult. Sci. J. 16 (2000):42–43.
Van Immerseel, F., J. De Buck, F. Pasmans, G. Huyghebaert, F. Haesebrouck, and R. Ducatelle. “Clostridium perfringens in poultry: an emerging threat of animal and public health.” Avian Pathology 33 (2004): 537-549. https://doi.org/10.1080/03079450400013162
Van Immerseel, F., J.I. Rood, R.J. Moore, and R.W. Titball. “Rethinking our understanding of the pathogenesis of necrotic enteritis in chickens.” Trends in Microbiology 17 no. 1 (2008):32-36. https://doi.org/10.1016/j.tim.2008.09.005
Wade, B., A.L. Keyburn, T. Seemann, J.I. Rood, and R.J. Moore. “Binding of Clostridium perfringens to collagen correlates with the ability to cause necrotic enteritis in chickens.” Veterinary Microbiology 180 no. 3–4 (2015): 299-303. https://doi.org/10.1016/j.vetmic.2015.09.019
Williams, R.B. “Intercurrent coccidiosis and necrotic enteritis of chickens: rational, integrated disease management by maintenance of gut integrity.” Avian Pathology 34 no. 3 (2005):159-180. https://doi.org/10.1080/03079450500112195
Yang , C.M., G.T. Cao, P.R. Ferket, T.T. Liu, L. Zhou, L. Zhang, Y.P. Xiao, and A. G. Chen. “ Effects of probiotic, Clostridium butyricum, on growth performance, immune function, and cecal microflora in broiler chickens.” Poultry Science 91 (2012): 2121–2129. https://dx.doi.org/10.3382/ps.2011-02131
Egg immunoglobulins as a plasma replacer
Animal plasma has been widely used in piglet feeding, not only as a protein source, but also as a tool to reduce gastrointestinal disorders after weaning. Drs FELLIPE BARBOSA and INGE HEINZL* consider a safe alternative in order to keep animals healthy and to avoid loss of performance.
The recent developments surrounding the health risks associated with using animal plasma as a piglet feed ingredient is growing serious concerns in China. After the reported cases of African swine fever (ASF) commencing in August 2018, the Chinese government decided to ban the use of pig blood (and its by-products) in animal feed for some time.
The reason for the temporary ban of pig blood ingredients: African swine fever. ASF is a viral disease of pigs and wild boars. The virus causes a lethal hemorrhagic disease in pigs. In some cases, the death of infected animals can occur during one week after the infection. There are no vaccines against the ASF Virus. When it hits the herd it is virtually impossible to stop its spread contaminating all animals.
Spreading of the virus occurs as follows:
• contact with contagious pigs from infected areas,
• contact with contaminated materials, being fed with kitchen waste and
• non-trusted animal origin feed ingredients.
There is a risk of pig blood carrying different types of viruses like ASF virus. Therefore, from time to time the use of ingredients based on blood is questioned by pig producers. To minimise this risk, the use of ingredients derived from pig slaughterhouses (including animal plasma) in pig feed is no longer allowed in China. This measure will cause not only a protein deficit in piglet feeds but also reduced protection of weaned piglets when intestinal disorders are concerned.
Immunoglobulins from animal plasma and its benefits on reducing post-weaning diarrhea (PWD)
The use of animal plasma has a positive effect on post-weaning performance of piglets. It is generally known that as a palatable ingredient, animal plasma stimulates feed intake. This results in better growth and a higher post-weaning performance in piglets. However, a closer inspection on the mode of action of spray dried plasma reveals its properties as an immune-ingredient and shows its supporting effect on the overall health status of the animals. Scientific publications showed that the positive influence on growth when feeding plasma to piglets is mainly due to its “immunoglobulin fraction”. This assigns to plasma a specific role in nutrition of weaned pigs to prevent PWD and to reduce the need for antibiotics.
Egg immunoglobulins: a natural way of protecting weaned piglets Globigen® Jump Start (EW Nutrition GmbH) is a functional and standardized product based on whole egg powder. It contains natural immunoglobulins (IgY – “immunoglobulins from yolk”) mixed with a carrier. IgY are cells of the immune system from birds similar to the IgG in mammals. They have the main function of identifying and neutralizing harmful substances in the body. IgYs are obtained through a non-invasive process and are natural ingredients from eggs. There is no connection with blood and slaughter by-products and therefore no risk of carrying animal diseases.
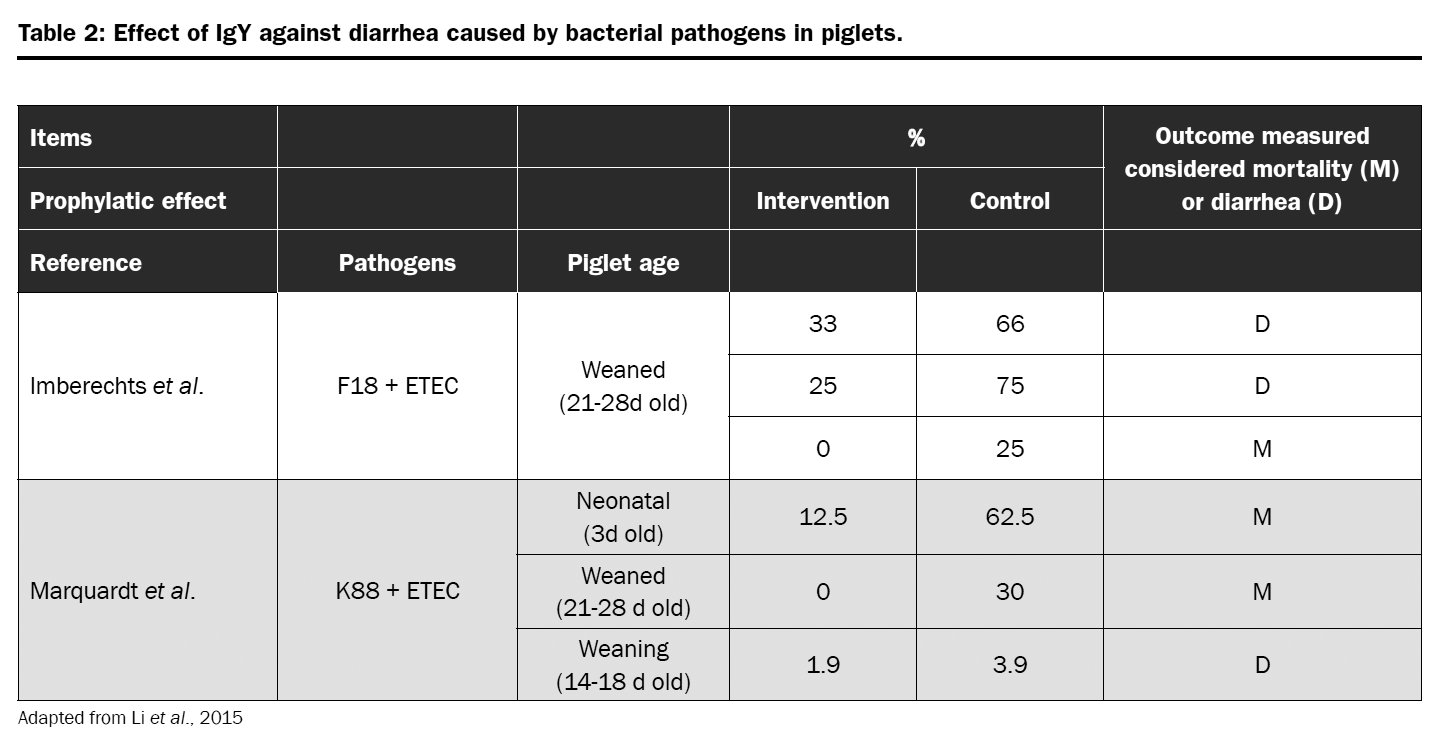
Globigen® Jump Start is used to support piglets during critical stages of life, as long as their natural immunity is not completely developed. Scientific data confirmed that the IgY present in egg powder are capable of supporting intestinal health and growth performance of newly weaned piglets. More recently, also the possibility of using immunoglobulins as alternatives to zinc oxide (ZnO) and in-feed antibiotics (Hedegaard et al., 2017; Li at al. 2015) were evaluated with promising results.
Better results than plasma IgG: understanding the antigens causing post-weaning diarrhea
Animal plasma is a by-product of the meat industry. The animals slaughtered were possibly exposed to various diseases over their whole life. It cannot be considered as a standardized product in terms of immunoglobulins (either quantity nor quality). The Ig contained could be useful but also totally useless, depending on the pathogens the animals have been confronted with. As a source of immunoglobulins Globigen Jump Start is a costefficient and effective alternative to replace plasma in piglets’ diets. Its IgY content will have the same protection effect in the gut as IgG, but the nutritionist will have the possibility of choosing different protein sources in the market, either because of price or availability of raw materials. Our recommendation is that 40kg of plasma can be replaced by 2kg of Globigen Jump Start supplied with different high digestible protein sources.
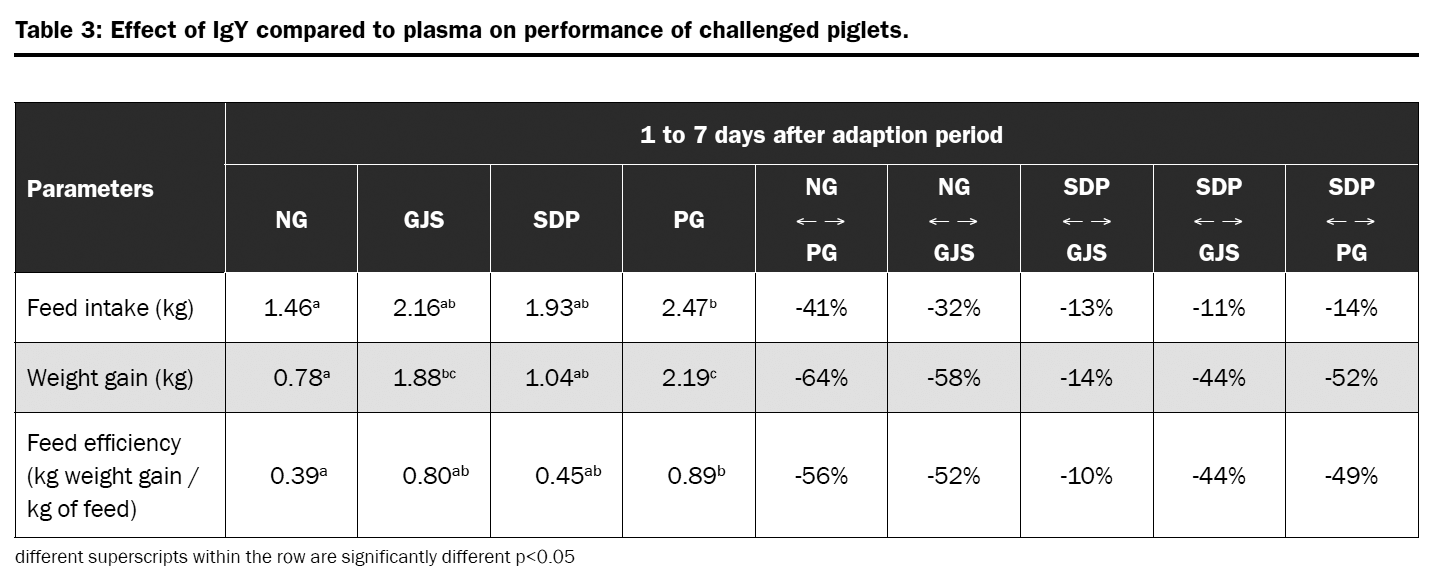
A piglet trial was conducted with the objective of evaluate the efficacy of egg immunoglobulins on performance parameters of weaned piglets and to evaluate it as a substitute for animal plasma. Piglets were challenged with F4 and F18 entertoxic E. coli (ETEC) strains and feed either 2kg of Globigen Jump Start (GJS) or 40kg of spray dried plasma (SDP) in the weaner diet. The comparison was also done to a negative group (NG – microbiological challenge and no protection in the diet); and a positive group (PG – no microbiological challenge and antibiotics + ZnO in the diet).
Piglets from NG had lower feed intake, weight gain, and feed efficiency than animals from PG. The same was observed for piglets from GJS and SDP group. However, the impact of bacterial challenge on weight gain was lower for GJS piglets than for SDP (-14% and -52% when compared to PG); whereas feed intake was similar for both groups (-13% and -14% when compared to PG). The results showed that piglets receiving GJS where more efficient on converting feed into growth even when challenged when compared to SDP animals.
Trial conclusion
In this trial, the product based on egg immunoglobulins showed better influence on the performance of piglets than blood plasma. This may be due to the fact that the quality of the plasma depends on the animals slaughtered and on their contact with diseases, determining how much and which antibodies are available in this feed.
Additionally, blood plasma includes the danger of infectious diseases.
Safe and standard: free of swine related diseases and ruminant material
EW Nutrition clearly understands the importance of maintaining standardisation. It is a key factor for the customers to have a product that they can depend on every day.
Therefore, trough specific steps during the production of Globigen products, EW Nutrition ensures product quality. During production, all eggs are pasteurised and dried to a whole egg powder. In between steps include microbiological analysis, Salmonella, and avian disease controls to ensure the final product is free of the mentioned threats. Furthermore, as Globigen products are originated from laying hen farms there is no risk of contamination with any swine disease, like the devastating ASF. Finally, Globigen products do not contain any raw materials produced from, or substances derived from ruminants nor do the products come in contact with risk materials during the whole process (not be at risk for carrying transmissible spongiform encephalopathy or bovine spongiform encephalopathy – BSE).
ASIAN FEED MAGAZINE – February/March 2019
Understanding the dangers of mycotoxins for breeder hens
Breeding hens are a valuable asset for the poultry industry, as they produce the hatching eggs and day-old chicks. It is therefore important to manage contamination as well as possible. Mycotoxin management is part of that.
As the producers of hatching eggs and day-old chicks, breeding hens are the backbone of the poultry industry. Hence it is common practice to pay particular attention to this valuable asset’s feed, selecting raw materials of high nutritional quality and safety. However, in any feed formulated for animals in production and reproduction, studies show that it is almost inevitable to find a certain level of mycotoxin contamination.
Mycotoxins exert toxic effects mainly on the gastrointestinal tract, liver, and kidneys and can accumulate in some tissues but also in the eggs. Mycotoxin contamination in breeding birds rations does not always lead to visible symptoms, such as when trichothecenes cause oral lesions. However, it may influence productivity, egg quality, hatchery performance, as well as chick quality and immunity. Mycotoxin risk management is thus an essential part of managing breeder hens.
Type of mycotoxin and exposure time determine effect on egg production
Mycotoxicosis in hens can cause reduced egg production, most likely because it causes a decrease in protein synthesis. A lower synthesis of albumin results from a degeneration of the liver tissue due to aflatoxin, ochratoxin, T2 and DON exposure. The liver then may look pale, friable and occasionally shows superficial haemorrhages.
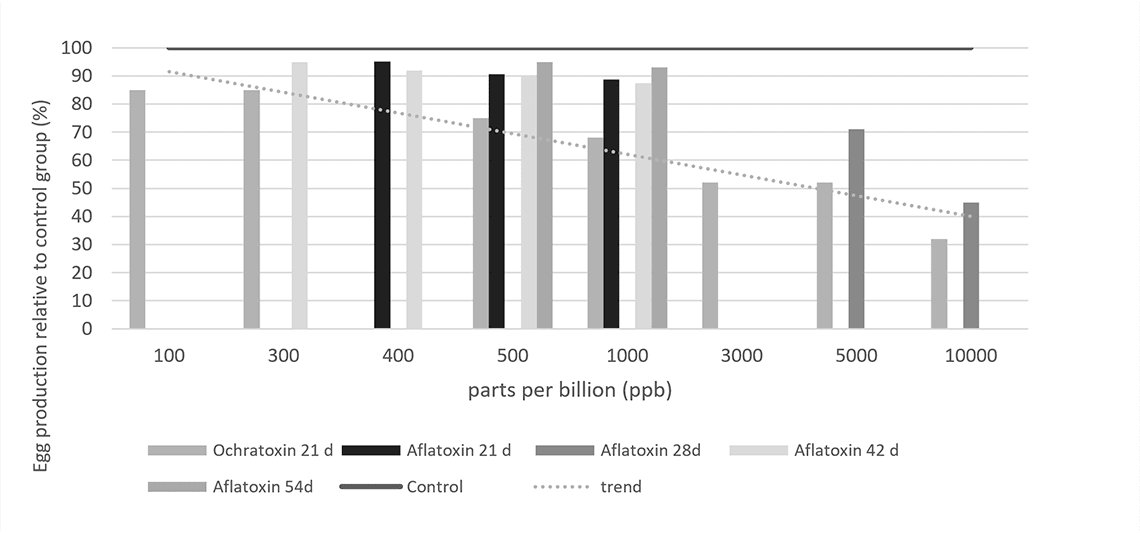
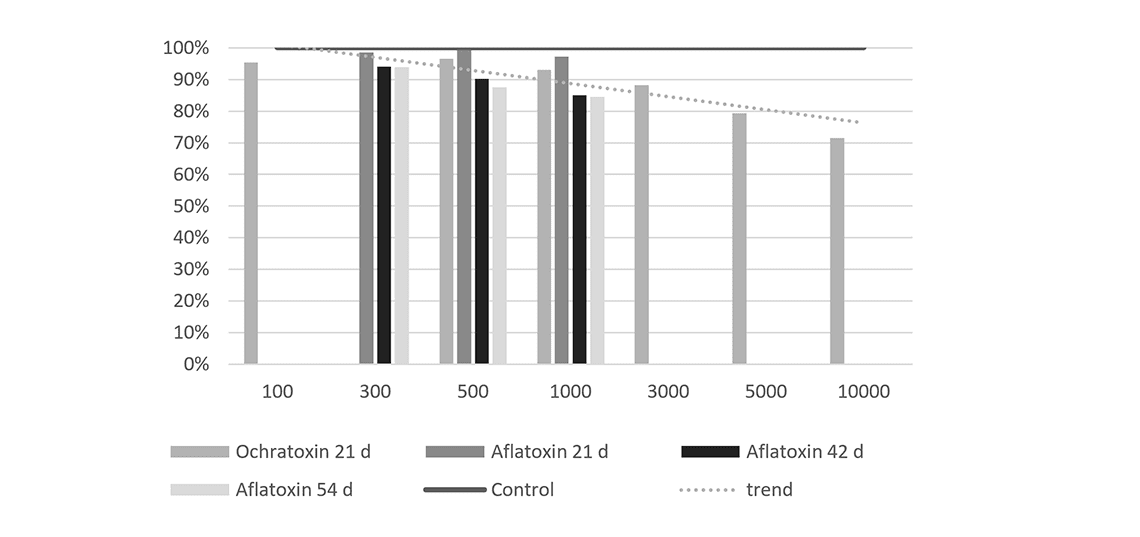
The contamination levels at which these effects can be observed are as low as 100ppb in feed, for example during a 21 day exposure to ochratoxin (Figure 1). With increasing levels of the toxin, production further decreases. A similar effect is observed when breeder hens are exposed to aflatoxins.
Figure 1 – Effect of mycotoxins on egg production, compared to non-contaminated control (=100 %).
Egg production, however, is not the only parameter that is affected when breeding hens are exposed to mycotoxins. Earlier on in the reproductive cycle they already impact on embryonic mortality and hatchability. These effects are potentially more severe and may even occur without any noticeable change in the number of eggs produced.
Mycotoxins’ insidious consequences for eggshell quality and embryonic mortality
The eggshell is important to protect the progeny: thin and fragile shells can increase embryonic mortality, lower embryonic weight gain and decrease hatchability. Egg shell quality is a function of the hen’s calcium and vitamin D3 metabolism. The bioavailability of calcium and of vitamin D3 depends on intestinal integrity and on the production of enzymes and transporters that aid in feed metabolism. These processes can be adversely affected by aflatoxins, DON, T2, and Fumonisins.
The gastrointestinal tract is not the only site of mycotoxin action, however. Mycotoxins such as aflatoxins and ochratoxins have nephrotoxic effects, affecting calcium metabolism and increasing its excretion via the urine, while lowering its levels in blood serum.
Moreover, mycotoxins damage the liver, which plays a central role in egg production, being responsible for vitamin D3 metabolism and the synthesis of the lipids that make up the yolk. Moreover, the synthesis of transporters for lipids, calcium and carotenoids ̶ important components of the egg ̶ also takes place in liver. When liver function is impaired, the internal and external quality of the egg declines, which, in the end, affects the production of day-old chicks.
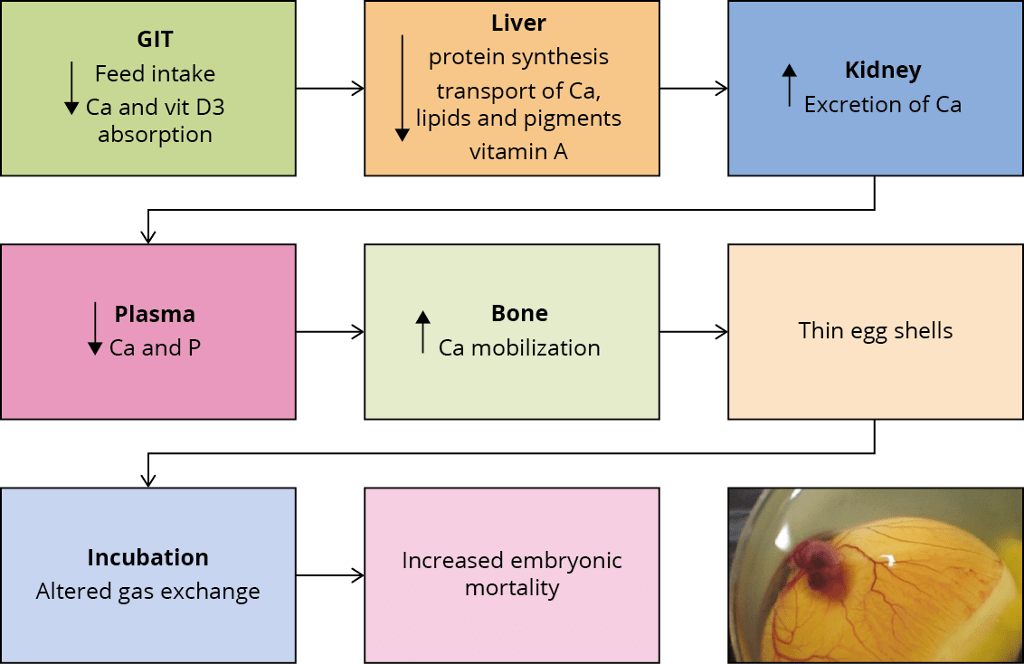
Figure 2 – Effects of mycotoxins on eggshell quality and embryonic mortality.
Figure 2 summarises the possible ways in which mycotoxins can negatively affect eggshell quality and, as a consequence, increase embryonic mortality. If a hen’s intestinal integrity is compromised, the utilisation of nutrients decreases. Liver and kidney damage leads to a diminished availability of calcium and other nutrients necessary for egg formation. The birds’ calcium (and phosphorus) levels in the plasma are then lower and may lead to a greater mobilisation of calcium from the bones. However, this response cannot be maintained and the eggs get a thinner shell.
The thickness of the eggshell influences the egg’s moisture loss and exchange with the environment during the incubation period. An eggshell of optimal quality does not allow the loss of nutrients and prevents bacterial contamination. Thinner eggshells are less able to fulfil these functions, leading to higher embryo mortality.
Figure 3 – Effects of mycotoxins on embryonic mortality
Figure 3 shows the effect of different mycotoxins on embryonic mortality. Incremental levels of ochratoxin or aflatoxin heighten embryonic mortality in a range from 1.5 to 7.5 times the embryonic mortality of the control group. In some cases, embryos are affected even when the hens received feed contaminated with mycotoxin levels that are within the guidelines suggested by the EFSA.
For example, an exposure to 4900ppb of DON for ten weeks increases the number of embryos with abnormalities. The causes are not entirely clear, as only traces of DON can be found in the egg. However, we do know that this mycotoxin can affect the protein synthesis at the level of the hen’s liver and therefore compromise the deposition of nutrients into the egg.
Mycotoxins’ effects on the progeny may cause long-term damage
Ochratoxin and aflatoxin can be transferred into the egg, where they exert toxicity on the embryos. This does not necessarily result in mortality. However, the chicks can suffer from a compromised immune function due to two reasons: lower transmission of antibodies from the hen and lower viability of the chickens’ immune cells, accompanied by a lower relative weight of the bursa of Fabricio and the thymus.
When both aflatoxin and ochratoxin are present in the feed, the effect on these parameters is synergistic. As a consequence of mycotoxin contamination, the animals’ immune response is impaired, which makes them more susceptible to infection. The final result could be an increased early chick mortality due to a higher incidence of bacterial and viral infections.
The transmission of other mycotoxins into the egg is minimal. While this means that a direct effect on the progeny is unlikely to occur, mycotoxin contamination still has a snowball effect: we have to consider the indirect effect of a lower deposition of nutrients on chick quality.
Prevention is key: mycotoxin risk management for breeder hens
The best approach to manage mycotoxin risk is to implement an integrated strategy that includes good crop and grain storing practices, regular raw material sampling and mycotoxin evaluation and analysis. Management tools (such as MasterRisk) can help to evaluate mycotoxin interactions and to choose the best strategy for dealing with specific mycotoxin challenges.
The results of mycotoxin analyses can be used to take decisions regarding the inclusion levels of raw materials and in choosing feed additives that counteract mycotoxins. Products based on plant extracts, yeast cell walls and clay minerals can help to stabilise a digestive system challenged by mycotoxins. They support the barrier function in the intestine, preventing the passage of mycotoxins into the bloodstream.
Phytomolecules are another piece of the puzzle: thanks to their antimicrobial, anti-inflammatory and antioxidant properties, they support liver function. This is particularly important for long-living animals prone to accumulating mycotoxins in their body tissues.
For a long time the “deleterious effects” of mycotoxins on breeder hens and “their repercussions on progeny health status and performance have not received from a scientific point of view as much attention”(Calini and Sirri, 2007) as they ought to have. However, now that the dangers of mycotoxins for breeder hens’ welfare, health and performance are better understood, it is clear that mycotoxin risk evaluation and management is central to successful poultry production.

 Rysunek 1: Stabilność fitomolekuł w warunkach przetwarzania, w stosunku do linii podstawowej zacieru (100%)
Rysunek 1: Stabilność fitomolekuł w warunkach przetwarzania, w stosunku do linii podstawowej zacieru (100%) Rysunek 2: Średni przyrost dobowy (g) – wyniki kilku badań przeprowadzonych z brojlerami
Rysunek 2: Średni przyrost dobowy (g) – wyniki kilku badań przeprowadzonych z brojlerami Rysunek 3: FCR – wyniki kilku badań przeprowadzonych z brojlerami
Rysunek 3: FCR – wyniki kilku badań przeprowadzonych z brojlerami