Eindämmung von Eimeria-Resistenzen in der Broiler-Produktion mit phytogenen Lösungen
Dr. Ajay Bhoyar, Global Technical Manager, EW Nutrition
In der modernen, intensiven Geflügelproduktion stellt die zunehmende Gefahr resistenter Eimeria-Stämme eine erhebliche Herausforderung für die Nachhaltigkeit der Broilerhaltung dar. Eimeria-Arten, die Resistenzen gegen herkömmliche Maßnahmen entwickeln können, sind zu einem weltweiten Problem geworden. Die Resistenz gegenüber herkömmlichen Medikamenten und die Sorge um Rückstände haben den Bedarf an natürlichen, sicheren und wirksamen Alternativen verstärkt.
Mehrere pflanzliche Verbindungen wie Saponine, Tannine, ätherische Öle, Flavonoide, Alkaloide und Lektine wurden intensiv auf ihre kokzidienhemmenden Eigenschaften untersucht. Unter ihnen haben sich insbesondere Saponine und Tannine bestimmter Pflanzen als wirksame Mittel im Kampf gegen diese widerstandsfähigen Protozoen erwiesen. Im Folgenden werden innovative Strategien vorgestellt, die das Potenzial dieser Stoffe – insbesondere von Saponinen und Tanninen – nutzen, um Verluste zu vermeiden, indem das Risiko resistenter Eimeria-Stämme in der Geflügelproduktion gemindert wird.
Was hat es mit resistenten Eimeria in der Broilerhaltung auf sich?
Die wissenschaftliche Abteilung der Weltgesundheitsorganisation (World Health Organization, 1965) definierte Resistenzen allgemein als die Fähigkeit eines Parasitenstamms, trotz Verabreichung und Aufnahme eines Medikaments in der empfohlenen Dosis zu überleben und/oder sich zu vermehren.
Die hohe Reproduktionsrate von Eimeria-Arten ermöglicht es ihnen, sich schnell weiterzuentwickeln und Resistenzen gegenüber den eingesetzten Medikamenten zu entwickeln. Darüber hinaus können resistente Eimeria-Stämme dank ihrer widerstandsfähigen Oozysten in der Umwelt überdauern, was zu einer erneuten Infektion von Tieren und einer weiteren Verbreitung führt.
Resistente Eimeria-Stämme stellen viele Herausforderungen in der modernen Geflügelhaltung dar und beeinträchtigen die Produktivität und wirtschaftliche Nachhaltigkeit erheblich. Eine der größten Schwierigkeiten ist die sinkende Wirksamkeit herkömmlicher Kokzidiostatika.
Unterschiedliche Arten von Eimeria-Resistenz
Es gibt verschiedene Ursachen dafür, warum Eimeria gegen bestimmte Wirkstoffe resistent sind.
Erworbene Resistenz entsteht durch eine vererbbare Abnahme der Empfindlichkeit bestimmter Stämme und Arten gegenüber Medikamenten im Laufe der Zeit. Diese kann partiell oder vollständig sein, abhängig vom Ausmaß des Empfindlichkeitsverlustes. Es besteht ein direkter Zusammenhang zwischen Wirkstoffkonzentration und Resistenzgrad. Ein Stamm, der durch eine bestimmte Dosis kontrolliert wird, kann bei einer niedrigeren Konzentration Widerstand zeigen.
Kreuzresistenz bedeutet, dass Resistenzen gegenüber verschiedenen Wirkstoffen mit ähnlichem Wirkmechanismus geteilt werden können (Abbas et al., 2011). Dies ist jedoch nicht immer der Fall (Chapman, 1997).
Mehrfachresistenz ist eine Resistenz gegenüber mehreren Wirkstoffen, auch wenn diese unterschiedliche Wirkmechanismen haben (Chapman, 1993).
Natürliche Stoffe können die Wirksamkeit Kokzidien-hemmender Maßnahmen wiederherstellen
Es wurde festgestellt, dass sich die Empfindlichkeit eines Parasiten gegenüber einem Wirkstoff wieder einstellen kann, wenn dieser für eine Zeit nicht mehr eingesetzt oder mit einem anderen Wirkstoff kombiniert wird (Chapman, 1997).
Pflanzenstoffe und naturidentische Verbindungen sind bekannt für ihre antimikrobielle und antiparasitäre Wirkung und stellen daher ein wertvolles Mittel gegen Eimeria dar (Cobaxin-Cardenas, 2018). Ihre Wirkmechanismen umfassen den Abbau der Zellwand, Schädigung des Zytoplasmas, Ionenverlust mit Verringerung der Protonenmotorkraft und die Induktion von oxidativem Stress, was die Invasion hemmt und die Entwicklung der Eimeria-Arten stört (Abbas et al., 2012;Nazzaro et al., 2013). Natürliche Kokzidien hemmende Mittel bieten einen neuen Ansatz zur Bekämpfung der Kokzidiose, insbesondere angesichts zunehmender Resistenzen in der kommerziellen Geflügelproduktion (Allen and Fetterer, 2002).
Saponine und Tannine: Die Abwehrkräfte der Natur gegen Eimeria
Phytogene Lösungen, speziell auf Basis von Saponinen und Tanninen, haben sich als vielversprechende Alternativen zur Minderung des Eimeria-Problems in der Geflügelproduktion herausgestellt. Mit ihrer Hilfe können Tierhalter die Widerstandskraft ihrer Bestände stärken, damit die Tiergesundheit fördern und die Nachhaltigkeit der Branche unterstützen. Saponine sind Glykoside, die in vielen Pflanzen vorkommen und aufgrund ihrer Fähigkeit, in Wasser zu schäumen, charakteristische seifige Eigenschaften haben. Wenn es um Kokzidiose geht, können Saponine die Zellmembranen der Eimerien angreifen. Nach der Aufnahme mit dem Futter können Saponine die schützende Außenschicht der Eimeria schädigen, die Parasiten schwächen und sie für die Immunabwehr des Wirts angreifbar machen. Dieser Angriff mindert die Fähigkeit der Eimerien, sich an die Darmschleimhaut anzuheften und zu vermehren. Tannine sind polyphenolische Verbindungen mit adstringierender Wirkung, die in Blättern, Rinde oder Früchten vorkommen. Entscheidend für den Erfolg von tanninhaltigen Futterzusätzen ist die Auswahl der passenden Tanninsorte in der richtigen Menge und zum passenden Zeitpunkt.
Gegen Eimerien wirken Tannine auf unterschiedliche Weise: In den Parasiten binden sie sich an Proteine stören enzymatische Aktivitäten und Stoffwechselprozesse. Diese Beeinträchtigung schwächt die Eimerien und verringert ihre Fähigkeit, die Darmschleimhaut zu schädigen. Darüber hinaus wirken Tannine entzündungshemmend und reduzieren die durch Eimeria verursachten Entzündungen. Als Antioxidantien schützen sie zudem die Darmzellen vor oxidativem Stress.
Werden Saponine und Tannine dem Broilerfutter zugegeben, schaffen sie ein ungünstiges Umfeld für Eimeria und hemmen deren Wachstum und Vermehrung im Wirt. Zusätzlich stärken sie die natürlichen Abwehrkräfte der Tiere und erhöhen deren Widerstandskraft gegenüber Infektionen. So können die Auswirkungen resistenter Eimeria-Stämme wirksam kontrolliert und eingedämmt werden, was die Gesundheit der Geflügelbestände und eine nachhaltige Geflügelproduktion fördert.
Was ist Pretect D?
Pretect D ist eine einzigartige, proprietäre Mischung aus Phytomolekülen, darunter Saponine und Tannine, die dabei hilft, Kokzidioseprobleme in der Geflügelhaltung in den Griff zu bekommen. Es kann allein aber auch in Kombination mit Kokzidiose-Impfstoffen, Ionophoren oder chemischen Mitteln im Rahmen eines Shuttle- oder Rotationsprogramms eingesetzt werden.
Figure DE
Abb.1. Die wichtigsten Bestandteile von Pretect D
Wie wirkt Pretect D
Pretect D wirkt auf mehreren Ebenen, um die Darmgesundheit in problematischen Zeiten zu unterstützen. Dank seiner antiparasitären, entzündungshemmenden, immunmodulierenden und antioxidativen Eigenschaften:
• reduziert es die Oozysten-Ausscheidung und dämmt die Verbreitung der Krankheit ein
• fördert es die Wiederherstellung der Schleimhautbarriere und verbessert die Darmstruktur
• schützt es das Darmepithel vor Schäden durch Entzündungen und oxidativem Stress.
Die positiven Effekte von Pretect D
Die Einbindung von Pretect D ins Kokzidiose-Programm verbessert die allgemeine Darmgesundheit und die Produktionsleistung der Broiler.
In einer Studie mit Cobb 500 Masthähnchen, die einer Eimeria-Mischinfektion ausgesetzt waren, zeigte sich, dass die Gruppe mit Pretect D (500 g/Tonne Futter über 35 Tage) weniger Kokzidien-Läsionen hatte als die infizierte Kontrollgruppe (IK).
Abb. 2: Pretect D vermindert die Läsionen bei Broilern
In einer weiteren Feldstudie wurde ein herkömmliches Kokzidiose-Programm (Starter und Mastfutter 1 mit Narasin + Nicarbazin, Mastfutter 2 mit Salinomycin und im End- und Ausmastfutter keine Kokzidiostatika) mit einem Programm verglichen, das Kokzidiostatika mit Pretect D kombinierte (Starter und Mastfutter 1 mit Narasin + Nicarbazin, Mastfutter 2 und Endmastfutter mit Pretect D). Die Zugabe von Pretect D senkte die Oozystenzahl und den Läsionsindex signifikant im Vergleich zur Kontrollgruppe (Abb. 3 a+b)
Abb 3a und 3b. Pretect D vermindert durch Kokzidiose verursachte Läsionen und die Anzahl an Oozysten pro Gramm Kot (OPG)
Daraus ergab sich eine insgesamt bessere Produktionsleistung der Broiler mit Pretect D (Abb. 4a-c):
Abb. 4 a-c. Verbesserte gesamte Produktionsleistung mit Pretect D
Pretect D: Anwendungsstrategien
Die Einbeziehung einer wirksamen phytogenen Kombination ins Kokzidiose-Kontroll-Programm kann helfen, das Resistenzproblem zu mindern. Eine solche natürliche Lösung kann eigenständig – vorzugsweise in weniger herausfordernden Monaten – oder in Kombination mit chemischen Mitteln (Shuttle-/Rotationsprogramme) oder einem Kokzidiose-Impfstoff (Bio-Shuttle) eingesetzt werden, um den häufigen Einsatz von Medikamenten zu reduzieren.
Shuttle-Programme werden häufig bei der Kokzidiosebekämpfung eingesetzt und liefern zufriedenstellende Ergebnisse. Dabei werden Produkte aus verschiedenen Wirkstoffklassen innerhalb eines Bestandes verwendet. Zum Beispiel wird eine Wirkstoffklasse im Starterfutter, eine andere im Mastfutter und wieder die erste im Endmastfutter eingesetzt, gefolgt von einer Phase ohne Medikamente.
In Rotationsprogrammen werden Antikokzidia zwischen den Beständen gewechselt, nicht innerhalb eines Bestandes.
Abb. 5: Strategien zur Einbeziehung von Pretect D in Programme zur Kokzidiosebekämpfung
Fazit
Kokzidiose ist eine der wirtschaftlich bedeutendsten Geflügelkrankheiten, und die Entwicklung von Resistenzen bedroht die Rentabilität der Broilerhaltung. Daher ist die regelmäßige Überwachung der Eimeria-Arten und ihrer Resistenzen entscheidend, um geeignete Kokzidiostatika auszuwählen. Der Einsatz einer wirksamen phytogenen Lösung als strategische und taktische Maßnahme ist vielversprechend – und integrierte Programme werden auch in Zukunft eine wichtige Rolle im Resistenzmanagement spielen.
References:
Abbas, R.Z., D.D. Colwell, and J. Gilleard. “Botanicals: An Alternative Approach for the Control of Avian Coccidiosis.” World’s Poultry Science Journal 68, no. 2 (June 1, 2012): 203–15. https://doi.org/10.1017/s0043933912000268.
Abbas, R.Z., Z. Iqbal, D. Blake, M.N. Khan, and M.K. Saleemi. “Anticoccidial Drug Resistance in Fowl Coccidia: The State of Play Revisited.” World’s Poultry Science Journal 67, no. 2 (June 1, 2011): 337–50. https://doi.org/10.1017/s004393391100033x.
Allen, P. C., and R. H. Fetterer. “Recent Advances in Biology and Immunobiology of Eimeria Species and in Diagnosis and Control of Infection with These Coccidian Parasites of Poultry.” Clinical Microbiology Reviews 15, no. 1 (January 2002): 58–65. https://doi.org/10.1128/cmr.15.1.58-65.2002.
Chapman, H. D. “Biochemical, Genetic and Applied Aspects of Drug Resistance in Eimeria Parasites of the Fowl.” Avian Pathology 26, no. 2 (June 1997): 221–44. https://doi.org/10.1080/03079459708419208.
Chapman, H.D. “Resistance to Anticoccidial Drugs in Fowl.” Parasitology Today 9, no. 5 (May 1993): 159–62. https://doi.org/10.1016/0169-4758(93)90137-5.
Cobaxin-Cárdenas, Mayra E. “Natural Compounds as an Alternative to Control Farm Diseases: Avian Coccidiosis.” Farm Animals Diseases, Recent Omic Trends and New Strategies of Treatment, March 21, 2018. https://doi.org/10.5772/intechopen.72638.
Nazzaro, Filomena, Florinda Fratianni, Laura De Martino, Raffaele Coppola, and Vincenzo De Feo. “Effect of Essential Oils on Pathogenic Bacteria.” Pharmaceuticals 6, no. 12 (November 25, 2013): 1451–74. https://doi.org/10.3390/ph6121451.
Pop, Loredana Maria, Erzsébet Varga, Mircea Coroian, Maria E. Nedișan, Viorica Mircean, Mirabela Oana Dumitrache, Lénárd Farczádi, et al. “Efficacy of a Commercial Herbal Formula in Chicken Experimental Coccidiosis.” Parasites & Vectors 12, no. 1 (July 12, 2019). https://doi.org/10.1186/s13071-019-3595-4.
World Health Organization Technical Report Series No. 296, (1965) pp:. 29.
Optimierung der Qualität von Eintagsküken aus Perspektive des Elterntierbetriebs
Konferenzbericht
Bei der Poultry Academy von EW Nutrition im Herbst 2023 erklärte Judy Robberts, Technical Service Manager, Aviagen, dass es für den Erfolg einer Elterntierherde essentiell ist, qualitativ hochwertige Bruteier mit hoher Schlupffähigkeit zu produzieren und erstklassige Küken zu liefern. Dabei stellen sich zwei entscheidende Fragen: Welchen Einfluss hat der Zuchtbetrieb auf die Kükenqualität? Und welche Bereiche werden von Züchtern am häufigsten übersehen?
Legenester müssen sauber gehalten werden
Die Hygiene der Legenester ist entscheidend für hochwertige Bruteier. Kurz nach der Eiablage ist die Eierschale noch feucht, und die Cuticula bietet noch keinen wirksamen Schutz. Zudem kühlt das Ei in dieser Zeit von der Körpertemperatur der Henne (41 °C) auf Stalltemperatur ab. Beim Abkühlen zieht sich der Inhalt zusammen, es entsteht ein Unterdruck, der Luft einsaugt und die Luftkammer bildet. Mit dieser Luft können auch Bakterien leicht ins Ei eindringen. Deshalb dürfen nur Bruteier verwendet werden, die in sauberen Nestern gelegt wurden.
Eine saubere Umgebung der Nistplätze und eine regelmäßige Reinigung der Nestmatte oder häufiger Austausch des Einstreumaterials verringert das Risiko einer bakteriellen Kontamination. Saubere Nester und sauberes Nistmaterial sind unerlässlich, um Verunreinigungen zu vermeiden.
Regelmäßiges Absammeln und Abtransportieren
Nest-Eier sollten mindestens viermal täglich gesammelt werden – bei heißem Wetter noch öfter, da die Eier im Stall nicht ausreichend abkühlen können, um die Embryonalentwicklung zu unterbrechen. Die genauen Zeiten sollten so gewählt werden, dass höchstens 30 % der Eier pro Sammelrunde anfallen (mehr erhöht die Bruchrate). Zu beachten ist dabei:
Die meisten Eier werden morgens gelegt – die Sammelintervalle sollten entsprechend geplant sein.
Eier, die länger als empfohlen in Nestern oder auf Bändern liegen, haben ein höheres Risiko, zu verschmutzen oder beschädigt zu werden.
Die Übergänge zwischen den Förderbändern müssen glatt sein, damit sich die Eier nicht stauen und gegeneinanderstoßen.
Eier dürfen nie über Nacht in Nestern oder auf Bändern bleiben.
Eier in herkömmlichen Nestern sind anfällig für Zehenpicken oder Verschmutzungen durch andere Hennen.
Bodeneier (außerhalb der Legenester gelegte Eier) sollten häufiger gesammelt werden als Nest-Eier.
Eier sollten nicht in Papp-Eierkartons gesammelt werden, da das Faser-Material die Wärme des Eies speichert und das Abkühlen verzögert. Außerdem sind diese Eierkartons porös, können Keime/Bakterien/Pilze beherbergen und Ungeziefer anziehen.
Idealerweise sollten Bruteier mindestens 50 g wiegen und von Herden stammen, die mindestens 22 Wochen alt sind. Kleinere Eier von jungen Herden können zwar verwendet werden, aber Kükengewicht und Vitalität in den ersten Tagen sind nicht optimal. Ein Küken entspricht etwa 68 % des Eigewichts. Ein kleines Ei bringt also auch ein kleines Küken hervor.
Sauberkeit der Eier
Nach dem Sammeln von Bodeneiern und vor jeder Sammlung von Nesteiern müssen die Hände gewaschen werden. Bodeneier sollten niemals ins Nest zurückgelegt werden – auch wenn sie sauber aussehen. Das Waschen von Boden- oder Schmutzeiern entfernt die schützende Schicht der Schale. Ein gewaschenes Ei bleibt ein schmutziges Ei – nur ein Ei, das nie schmutzig war, ist wirklich sauber.
Eier sollten chemisch desinfiziert werden, da Kratzen, Reiben oder Waschen die Cuticula beschädigen und damit die physikalische und antimikrobielle Barriere entfernen. Da die Schalenpermeabilität nach 24 Stunden zunimmt, sollten Eier so schnell wie möglich desinfiziert werden. Die beliebteste Methode ist das Nebeln, da damit das Erreichen aller Eier gewährleistet wird und die Eier nicht nass werden.
Bodeneier sind keine Bruteier
Die Brüterei kann Fehler des Zuchtbetriebs nicht beheben. Daher sollten Bodeneier – also Eier, die außerhalb der Legenester gelegt wurden – AUF KEINEN FALL bebrütet werden. Sie haben eine höhere bakterielle Belastung als Nest-Eier und damit eine geringere Schlupffähigkeit. Zudem können sie als „Stinkbomben“ (im Englischen „Banger“ oder „Exploder“) andere Eier im Inkubator kontaminieren.
Die Aussortierung von Bodeneiern muss auf dem Betrieb erfolgen, damit keine verschmutzten Eier in die Brüterei gelangen. Wenn unbedingt nötig, sollten Bodeneier nur dann bebrütet werden, wenn die Nachteile bekannt sind und akzeptiert werden. Werden sie verwendet, müssen sie klar markiert und getrennt von Nest-Eiern gelagert werden, damit die Brüterei das Kontaminationsrisiko managen kann.
Bodeneier bergen ein deutlich höheres Risiko mikrobieller Kontamination, was Schlupfrate und Kükenqualität mindert
Eihygiene – bakterielle Belastung
Ei-Zustand
Gesamtkeime (cfu/cm2)
Frisch gelegt
300
Abgekühltes, sauberes Ei
3,000
“Sauberes” Bodenei
30,000
Schmutziges Ei
300,000
Die Anzahl der Bodeneier sollte überwacht und das Management angepasst werden, um diese zu minimieren. Bodeneier sind ein Problem, das durch gutes Herdenmanagement und eine geeignete Stall-Ausrüstung reduziert werden muss. Liegt der Anteil der Bodeneier über die Lebensdauer der Herde bei über 2–3 %, besteht Handlungsbedarf. Zu Beginn der Legeperiode ist der Anteil höher, sollte aber zum Höhepunkt der Legeleistung auf 1–2 % sinken.
Knickeier
Knickeier sind anfälliger für Infektionen und führen zu niedrigeren Schlupfraten und einer schlechteren Kükenqualität.
Einfluss von verschiedenen Eischalenverletzungen auf Schlupfrate und Kükenqualität
Eischale
Eigewicht beim Umsetzen (g)
Gewichtsverlust (%)
Befruchtung (%)
Schlupfrate (%)
Kükengewicht (g)
Küken-Uniformität (%)
Normal
62.0a
11.4c
97.8a
83.9a
48.9a
82.6
Sternriss
55.6b
20.7b
89.4b
49.4b
48.2a
70.3
Haarriss
53.1c
24.0a
83.3c
30.0c
45.6b
70.2
(Quelle: Khabisi et al., 2011 – Werte mit unterschiedlichen Buchstaben in einer Spalte unterscheiden sich signifikant, p ≤ 0,05)
Rissige Eier dürfen nicht bebrütet werden. Die Anzahl solcher Eier muss dokumentiert werden. Ist der Anteil zu hoch, müssen die Ursachen ermittelt und behoben werden.
Eilagerung auf dem Betrieb
Die Lagerung beginnt mit dem Legen der Eier, nicht erst mit der Ankunft in der Brüterei.
Die Eier müssen so schnell wie möglich unter 24 °C (physiologischer Nullpunkt) abgekühlt werden, um das Zellwachstum des Embryos bis zur Eiverarbeitung in der Brüterei zu stoppen. Dies minimiert Embryo-Verluste, maximiert die Schlupfrate und sichert die Kükenqualität. Die Eier sollten innerhalb von 4 Stunden nach dem Sammeln eingelagert werden.
Auf den Elterntierbetrieben werden die Eier bis zum Transport zur Brüterei gelagert. Die Lagerdauer hängt von der Lagerkapazität, dem Angebot an Bruteiern, der Kapazität der Brüterei und der Nachfrage nach Eintagsküken ab. Wenn der Betrieb einen klimatisierten Eilagerraum hat, reicht es, wenn die Brüterei die Eier zweimal pro Woche abholt. Gibt es keinen speziellen Lagerraum, müssen die Eier täglich transportiert werden. Unkontrollierte Temperaturschwankungen führen zu wiederholtem Start-Stopp-Wachstum der Keimscheibe und beeinträchtigen die Schlupffähigkeit.
Die Temperatur im Eilagerraum sollte höher sein als im Transportfahrzeug, und die Transporttemperatur sollte höher sein als die Lagertemperatur in der Brüterei. Dieser stufenweise Temperaturabfall verhindert Kondensation („Schwitzen“) auf den Eiern. Kondensation beeinträchtigt die natürliche Abwehr und bietet Bakterien ideale Bedingungen, um die Schale zu durchdringen. Schwitzen tritt besonders in heißen und feuchten Klimazonen häufig auf.
Eilagerräume sind wichtig, werden aber oft vernachlässigt. Beachtenswerte Punkte sind:
Konstante Temperatur rund um die Uhr (Isolierung minimiert Schwankungen)
Temperatur-Alarmsystem: maximal 21 °C, minimal 16–18 °C
Temperatur- und Feuchtigkeitssensoren richtig platzieren – nicht direkt an Wärme- oder Feuchtigkeitsquellen, nicht an Wänden, damit keine falschen Werte entstehen
Genauigkeit der Sensoren sicherstellen (Datenlogger empfohlen)
Ventilatoren für gleichmäßige Luftverteilung
Eier nicht direkt an Wände oder auf den Boden stellen, um Luftzirkulation und gleichmäßige Bedingungen zu gewährleisten
Direkten Luftstrom von Ventilatoren, Kühlern oder Befeuchtern auf die Eier vermeiden, um Feuchtigkeitsverlust und Temperaturunterschiede zu verhindern
Fazit
Der Elterntierbetrieb ist der Ausgangspunkt für die Kükenqualität. Sorgfalt und Hygiene im gesamten Prozess sind entscheidend. Durch Überwachung und Kontrolle können Schwachstellen erkannt und behoben werden, um weiterhin hochwertige Bruteier zu produzieren.
Optimierung der Qualität von Eintagsküken aus der Brütereiperspektive
Konferenzbericht
Bei der Poultry Academy von EW Nutrition im Herbst 2023, sprach Judy Robberts, Technical Service Manager, Aviagen über den Einfluss der Brüterein auf die Kükenqualität. Transport und Lagerung von Bruteiern, prophylaktische Maßnahmen sowie der Transport der Eintagsküken spielen eine entscheidende Rolle. Werden diese Bereiche schlecht gemanagt, können sie die Investitionen und Verbesserungen auf dem Elterntierbetrieb oder sogar in der Brüterei selbst zunichtemachen..
Transport der Bruteier vom Elterntierbetrieb zur Brüterei
Der Transport der Bruteier vom Elterntierbetrieb zur Brüterei ist von entscheidender Bedeutung: Der LKW muss vor der Nutzung gereinigt und desinfiziert werden, um eine Verbreitung von Krankheitserregern zu verhindern, und es sollten ausschließlich Fahrzeuge verwendet werden, die nur für den Transport von Bruteiern vorgesehen sind. Eier sollten immer mit dem spitzen Ende nach unten transportiert werden, damit sich die Luftkammer nicht löst.
Die Temperatur im Lagerraum für Bruteier auf dem Betrieb sollte höher sein als die Temperatur im Transportfahrzeug, um Kondensation (auch als „Schwitzen“ bezeichnet) auf den Eiern zu vermeiden. Kondensation auf der Eierschale beeinträchtigt die natürlichen Abwehrmechanismen und bietet Bakterien ideale Bedingungen, um zu wachsen, die Schale zu durchdringen und das Ei zu kontaminieren. Kondensation tritt häufiger in heißen und feuchten Klimazonen auf, wie sie in vielen Teilen Asiens üblich sind. Selbst wenn die Temperaturen in Lagerraum und Transportfahrzeug gleich sind, kann es beim Be- und Entladen, besonders an warmen, feuchten Tagen, zum Schwitzen kommen. In solchen Fällen kann eine höhere Lagertemperatur von 23 °C anstelle der allgemein empfohlenen 18–20 °C in Betracht gezogen werden.
Plötzliche Temperaturschwankungen sollten vermieden werden. Dafür eignen sich Temperatur-Datenlogger, die während des Transports eingesetzt werden, um Schwankungen zu dokumentieren. Im Brütereibetrieb hilft die Messung der Eier-Innentemperatur an verschiedenen Orten bei jeder Lieferung, um die Transportbedingungen zu überprüfen. Die relative Luftfeuchtigkeit im LKW sollte bei 65–70 % liegen.
Lagerung der Eier in der Brüterei
Die Lagerung beginnt mit der Eiablage, nicht erst mit der Anlieferung in der Brüterei. Lagerräume werden oft übersehen, sind aber entscheidend. Wichtige Punkte sind:
Konstante Temperatur rund um die Uhr (Isolierung und Ventilatoren minimieren Schwankungen)
Vermeidung von Kondensation
Eier nicht direkt an Wände oder auf den Boden stellen, um die Luftzirkulation zu maximieren und gleichmäßige Bedingungen zu schaffen,
Alarmsysteme: maximal 21 °C, minimal 16–18 °C,
Sensoren sollten genau sein (Datenlogger werden empfohlen),
Sensorplatzierung: Platzierung nicht direkt in der Nähe von Wärme- oder Feuchtigkeitsquellen, um Fehlmessungen zu vermeiden; ebenso nicht an Wände stellen, damit die Luft zirkulieren kann.
.
Temperatur und Lagerungszeit
„Die Lagertemperatur sollte abhängig von der Lagerdauer gewählt werden“, rät Judy Robberts. Eier, die innerhalb von vier Tagen nach dem Legen eingelegt werden, müssen nicht unter 20 °C gelagert werden; hier gelten 21–22 °C als optimal. Diese relativ hohe Temperatur fördert die Verdünnung des Eiklars, was den Gasaustausch während der frühen Brut verbessert. Gleichzeitig ist sie niedrig genug, um die Vitalität des Embryos zu erhalten. Die besten Schlupfergebnisse werden mit 3-7 Tage alten Eiern erzielt. Für eine Lagerung länger als sieben Tage sollten kühlere Temperaturen angestrebt werden, um eine Minderung der Schlupfrate durch Absterben der Embryozellen und Qualitätsverlust im Ei zu verringern. Liegt die Lagerzeit unter sieben Tagen, wird eine Temperatur von 16–18 °C empfohlen, bei längerer Lagerung meist 10–12 °C. Eier von jungen Elterntierherden eignen sich besser für längere Lagerzeiten, da die Qualität des Eiklars bei jüngeren Tieren höher ist.
Temperaturunterschiede führen dazu, dass die Eier zu unterschiedlichen Zeiten Bruttemperatur erreichen und folglich zu unterschiedlichen Zeiten schlüpfen, was das Schlupffenster vergrößert.
Relative Luftfeuchtigkeit
Um ein Austrocknen der Eier zu verhindern und die Qualität im Inneren zu erhalten, sollte die relative Luftfeuchtigkeit im Eilagerraum bei 70–80 % liegen. Der Nebel sollte fein sein, damit die Eier nicht nass werden. Da verschmutzte Geräte eine erhebliche Keimquelle darstellen, kann eine regelmäßige Wartung und Reinigung der Befeuchter das Risiko einer Kontamination der Eier mindern. Für die Platzierung der Wagen, Abstände und Luftzirkulation im Lagerraum der Brüterei gelten die gleichen Empfehlungen wie für den Lagerraum auf dem Betrieb. Ebenso gelten dieselben Empfehlungen für die Temperaturüberwachung und Platzierung der Thermometer.
Wartung nicht vergessen
Wartung ist oft reaktiv statt präventiv – meist wird etwas erst repariert, wenn es kaputt ist. Dies kann die Schlupfrate und die Kükenqualität beeinträchtigen. Einige Punkte, die für einen Wartungsplan relevant sind:
Eine für Wartung verantwortliche Person benennen, die direkt an den Brütereileiter berichtet
Eine Liste aller zu wartenden Geräte mit Wartungsintervallen erstellen
Alle durchgeführten Wartungsarbeiten dokumentieren
Wartung schließt die Kalibrierung von Geräten ein
Ersatzteile vorrätig halten
Auch das Gebäude und Zusatzgeräte ins Programm aufnehmen
Transport der Eintagsküken
Der Transport kann die Qualität der Eintagsküken nicht verbessern, aber er kann das Wohlergehen, Wachstum, die Entwicklung und Leistung der Küken erheblich beeinträchtigen.
Werden Küken außerhalb ihrer thermoneutralen Zone (32–35 °C) transportiert, verbrauchen sie die Nährstoffe aus dem Dottersack deutlich schneller, um ihre Körpertemperatur (40–41 °C) zu halten. Liegt die Kerntemperatur nach dem Schlupf über 41 °C, beginnen die Küken zu hecheln, was zu Wasserverlust und Austrocknungsgefahr führt. Liegt sie unter 39,5 °C, sinken Aktivität und Futteraufnahme. Sollte die optimale Temperatur in den Kükenboxen aufgrund technischer Einschränkungen des Transportmittels nicht erreicht werden können, muss die Anzahl der Küken pro Box angepasst werden. Optimale Transportbedingungen vom Brütereibetrieb bis zum Stall sind entscheidend für die spätere Leistung der Küken.
Fazit
Eine moderne Brüterei ist eine große Investition – daher ist es nur logisch, auf jedes Detail zu achten, um die Qualität der Bruteier zu erhalten und hochwertige Küken zu produzieren. Faktoren wie die Lagerbedingungen spielen eine entscheidende Rolle für eine maximale Schlupfrate. Durch Überwachung und Überprüfung können Schwachstellen erkannt und behoben werden, um weiterhin hochwertige Bruteier zu liefern. Der Transport der Eintagsküken muss sicherstellen, dass die Tiere in demselben Zustand im Stall ankommen, in dem sie die Brüterei verlassen haben.
Muskel- und Fettwachstum: Auswirkungen auf die Futterverwertung bei Masthühnern
Konferenzbericht
Auf der jüngsten Veranstaltung der EW Nutrition Poultry Academy in Jakarta, Indonesien, betonte Dr. Steve Leeson, emeritierter Professor an der University of Guelph, Kanada, dass die Futteraufnahme der wichtigste Faktor für die Wachstumsrate von Masthühnern ist, während die Futterverwertung stärker vom Muskelaufbau als vom Fettwachstum beeinflusst wird.
Die Zucht auf schnelles Wachstum hat dazu geführt, dass der Appetit indirekt zu einem wichtigen Kriterium bei der Selektion moderner Masthuhn-Linien geworden ist. Diese Linien sind sehr magerfleischig. Dr. Leeson erklärte: „Die Förderung des Muskelwachstums im Vergleich zum Fettwachstum verbessert stets die Futterverwertung. Der Grund dafür ist ganz einfach – 1 kg Fett enthält 9.000 kcal, während 1 kg Muskelfleisch, das zu 80 % aus Wasser besteht, nur 1.000 kcal liefert. Daher reagieren Masthühner heute sehr gut auf Aminosäuren. Das bedeutet, dass Muskelaufbau fast zehnmal effizienter ist als Fetteinlagerung.“ Dr. Leeson merkte auch an, dass die Fütterung des Broilerkükens heute wichtiger ist denn je. „Das Körpergewicht am Tag 7 ist heute ein Standardmaß zur Messung der Produktivität. Jedes Gramm Körpergewicht im Alter von 7 Tagen entspricht 10 Gramm im Alter von 35 Tagen.“
Faktoren, die die Futteraufnahme beeinflussen
Um das genetische Potenzial zu maximieren und die Fresslust moderner Masthühner auszunutzen, ist es entscheidend, Faktoren, die die Futteraufnahme hemmen, auf ein Mindestmaß zu beschränken. Dazu gehören:
Futterpartikelgröße — Die Futteraufnahme wird durch die Fütterung möglichst großer Futterpartikel maximiert
Besatzdichte — Es ist unwahrscheinlich, dass die Fütterung nach dem 28. Lebenstag bei einer Besatzdichte von mehr als 35 kg/m2 wirklich noch ad libitum erfolgen kann
Umgebungstemperatur — die maximale Futteraufnahme findet nach der Aufzuchtphase bei einer Temperatur von 15°C statt, ein optimales Futter/Zuwachs-Verhältnis erzielt man bei etwa 26°C
Beleuchtung — je länger die Lichtphasen sind, desto höher ist die Futteraufnahme. Es sind jedoch 4 Stunden Dunkelheit erforderlich, um die Gesundheit der Tiere und ihre Immunabwehr nicht zu beeinträchtigen.
Energieniveau des Futters — Masthühner fressen noch immer nach Energiebedarf.
Besatzdichte und eingeschränkte Fütterung
Bis zum Alter von 21 Tagen erreichen Masthähnchen ihre erwarteten Gewichte, ab 28 Tagen jedoch liegen sie unter ihrem genetischen Potential. Oft wird dieser Wachstumsabfall irrtümlicherweise der Umstellung auf Endmastfutter, einer subklinischen Erkrankung oder anderen Problemen zugeschrieben. In vielen Fällen ist er jedoch einfach nur auf eine, durch den begrenzten Zugang zu Futterautomaten verursachte, geringere Futteraufnahme zurückzuführen. Warum? Bei höheren Besatzdichten, die die Profitabilität pro Stall und nicht pro Tier maximieren, konkurrieren die nun größeren Tiere um den Platz am Futterautomaten, was möglicherweise eine Futteraufnahme ad libitum verhindert. Masthähnchen müssen pro Stunde etwa 8 Minuten lang fressen, dies erfolgt aber normalerweise nicht auf einmal.
Das genetische Potenzial für die Futteraufnahme (Gramm/Tag) hängt vom Alter der Tiere ab. Bei einem Masthähnchen von 21 bis 42 Tagen entspricht die Futteraufnahme beispielsweise dem Alter (Tage) x 6, sodass ein 28 Tage altes Masthähnchen 168 Gramm Futter pro Tag aufnehmen kann.
Umgebungstemperatur
Moderne Masthühner reagieren zunehmend empfindlich auf Hitzestress. Wir können den Gesamtenergiebedarf beeinflussen, indem wir den Energiebedarf zur Aufrechterhaltung der Körpertemperatur reduzieren. Das Halten von Masthühnern bei <15 °C ist bei hohen Besatzdichten eine Herausforderung. Wenn Masthühner nahe am thermischen Neutralpunkt von etwa 24 °C gehalten werden, wird der Energiebedarf unabhängig von der Besatzdichte minimiert.
Wie kann die Futteraufnahme gefördert werden?
Verbesserung der Pelletqualität
Bis zum Alter von 21 Tagen erreichen Masthähnchen ihre erwarteten Gewichte, ab 28 Tagen jedoch liegen sie unter ihrem genetischen Potential. Oft wird dieser Wachstumsabfall irrtümlicherweise der Umstellung auf Endmastfutter, einer subklinischen Erkrankung oder anderen Problemen zugeschrieben. In vielen Fällen ist er jedoch einfach nur auf eine, durch den begrenzten Zugang zu Futterautomaten verursachte, geringere Futteraufnahme zurückzuführen. Warum? Bei höheren Besatzdichten, die die Profitabilität pro Stall und nicht pro Tier maximieren, konkurrieren die nun größeren Tiere um den Platz am Futterautomaten, was möglicherweise eine Futteraufnahme ad libitum verhindert. Masthähnchen müssen pro Stunde etwa 8 Minuten lang fressen, dies erfolgt aber normalerweise nicht auf einmal.
Das genetische Potenzial für die Futteraufnahme (Gramm/Tag) hängt vom Alter der Tiere ab. Bei einem Masthähnchen von 21 bis 42 Tagen entspricht die Futteraufnahme beispielsweise dem Alter (Tage) x 6, sodass ein 28 Tage altes Masthähnchen 168 Gramm Futter pro Tag aufnehmen kann.
Umgebungstemperatur
Moderne Masthühner reagieren zunehmend empfindlich auf Hitzestress. Wir können den Gesamtenergiebedarf beeinflussen, indem wir den Energiebedarf zur Aufrechterhaltung der Körpertemperatur reduzieren. Das Halten von Masthühnern bei <15 °C ist bei hohen Besatzdichten eine Herausforderung. Wenn Masthühner nahe am thermischen Neutralpunkt von etwa 24 °C gehalten werden, wird der Energiebedarf unabhängig von der Besatzdichte minimiert.
Wie kann die Futteraufnahme gefördert werden?
Verbesserung der Pelletqualität
Eine höhere Pelletqualität bedeutet, dass die Tiere weniger Zeit zur Futteraufnahme benötigen und dadurch weniger Energie für die Erhaltung verbrauchen. Eine Verbesserung der Pelletqualität ermöglicht eine Reduzierung der scheinbar umsetzbaren Energie (AME) im Futter. Wenn beispielsweise die Pelletqualität von 60 % auf 80 % steigt, entspricht dies einer Erhöhung der Futterenergie um 60 kcal, ohne die Zusammensetzung der Nahrung zu ändern. Alternativ kann die AME um 60 kcal reduziert werden, was die Futterkosten senkt, ohne die Leistung zu beeinträchtigen.
Übergang von Krümelfutter zu Pellets
Der Übergang von Krümelfutter zu Pellets erfolgt in der kommerziellen Geflügelhaltung oft zu spät, was die Wachstumsrate einschränkt. Die Zurückhaltung, früher auf Pellets umzustellen, beruht auf der wahrgenommenen Futterverweigerung, weshalb das Umsteigen von Krümelfutter zu Pellets häufig erst im Alter von 21 bis 24 Tagen erfolgt. Masthühner fressen jedoch bereits sehr früh große Futterpartikel. Die vorübergehende Futterverweigerung dauert Minuten oder Stunden, nicht Tage. Futterreste werden überschätzt und betragen maximal nur 1–2 Gramm pro Tier. Die Futterverweigerung kann minimiert werden, indem der letzten Portion Krümelfutter 5 % Pellets zugesetzt werden und die erste Portion des Mastfutters zu 50 % aus Krümelfutter und zu 50 % aus Pellets besteht.
Größe der Pellets
Die Anpassung der Pelletgröße an das Vogelalter nimmt mit zunehmender Besatzdichte an Bedeutung zu. Dr. Leeson empfiehlt für die Tiere folgende ideale Pelletgrößen:
Prestarter (0-10 Tage) 2 mm
Starter (11-21 Tage) 3,5 mm kurz
Mastfutter 1 (22-32 Tage) 4 mm und
Endmastfutter (32+ Tage) 5 mm.
Wenn die Tiere Futter mit unterschiedlichen Partikelgrößen erhalten, bevorzugen sie ausnahmslos die größten Partikel. Von größeren Pellets müssen die Tiere weniger konsumieren und so weniger Zeit am Futterautomaten verbringen.
Fazit
Dr. Leeson kam zu dem Schluss, dass die Futteraufnahme die Wachstumsrate bestimmt. Daher sollten die Vögel so früh wie möglich auf Pellets umgestellt werden, und die Pelletgröße an die Bedürfnisse der Tiere angepasst werden. Jede Managementmaßnahme, die die Futteraufnahme einschränkt, wie eine hohe Besatzdichte oder Hitzestress, erfordert eine entsprechende Verzögerung bei der Futterumstellung.
Stoffwechselstörungen und Muskeldefekte
Konferenzbericht
Im Rahmen der EW Nutrition Poultry Academy im indonesischen Jakarta definierte Dr. Steve Leeson, emeritierter Professor der University of Guelph in Kanada, Stoffwechselstörungen wie folgt: Sie sind nicht ansteckend, treten bei entsprechender Ernährung unter „normalen“ Bedingungen auf und sind meist tierartspezifisch. Ihre Häufigkeit korreliert negativ mit der Produktivität. Obwohl die Genetik häufig eine große Rolle spielt, ist die genetische Selektion zur Bekämpfung des Problems oft der letzte Ausweg, da in der Regel eine negative Korrelation mit der Produktivität besteht.
Aszites
Aszites oder „Bauchwassersucht“ wurde erstmals in den 1970er Jahren beschrieben und ist heute wahrscheinlich die Stoffwechselstörung Nummer eins. Es handelt sich dabei um eine Flüssigkeitsansammlung im Bauchraum. Sie wird verursacht durch eine Reihe von Ereignissen, die bedingt durch den hohen Sauerstoffbedarf der Gewebe stattfinden. Ursprünglich trat die Erkrankung vor allem bei schnell wachsenden männlichen Masthühnern auf, die in großen Höhen gehalten wurden und einem gewissen Kältestress ausgesetzt waren. Heutzutage kann das Problem jedoch in jeder Höhenlage auftreten. In Extremsituationen kann die Mortalität bis zu 8% betragen, meistens sind es jedoch 1–3%. Da höhere Wachstumsraten hauptsächlich dafür verantwortlich sind, tritt das Problem heutzutage wieder häufiger auf.
Um Aszites möglichst zu vermeiden, können folgende Maßnahmen ergriffen werden:
Wachstumsrate begrenzen
Futterbeschaffenheit (Breifutter besser als Pellets)
Temperatur bei Tieren jeglicher Altersklasse niemals unter 15°C
Ausreichende Lüftung – Frischluft vs. Temperatur
Minimierung von Umweltschadstoffen wie Staub
Lichtprogramme (4-6 Stunden Dunkelheit)
Plötzlicher Tod (Sudden Death Syndrome, SDS)
SDS betrifft fast immer männliche Tiere, die ihr Schlachtgewicht erreichen. Häufig sind 1–5 % des Bestandes davon betroffen und im Alter von 21–35 Tagen ist SDS normalerweise die Haupttodesursache. Betroffene Tiere machen einen gesunden Eindruck, sind wohlgenährt und haben stets Futter im Verdauungstrakt. Der Tod tritt innerhalb von 1–2 Minuten ein und die Tiere werden meistens tot auf dem Rücken liegend aufgefunden. Es gibt nur wenige Veränderungen in der makroskopischen Pathologie. Das Herz kann Blutgerinnsel enthalten, die aber wahrscheinlich erst postmortal entstanden sind; die Herzkammern sind normalerweise leer. Die Diagnose erfolgt normalerweise durch Ausschluss anderer Krankheiten. Die Lunge ist häufig ödematös. Dies kann auch als Folge der längeren Rückenlage der Tiere auftreten, wenn die Flüssigkeit durch die Schwerkraft in die Lungenregion abfließt. Spezifische Gewebe- oder Blutbildveränderungen, die eine Diagnose ermöglichen, sind nicht erkennbar. SDS wird durch schnelles Wachstum hervorgerufen, in diesem Fall ist aber eine Eindämmung des Problems durch Nährstoffbeschränkung in unterschiedlichem Ausmaß möglich.
Spiking-Mortalitätssyndrom (SMS)
SMS ist durch eine schwere, unerklärliche Hypoglykämie gekennzeichnet und tritt immer im Alter zwischen 18 und 21 Tagen auf. Da es nur wenige Obduktionsberichte gibt, kommt es häufig zu Fehldiagnosen. Die Sterblichkeit kann zwische 2 und 3% betragen. Männchen sind anfälliger als Weibchen, wahrscheinlich weil sie schneller wachsen. Tiere, die ausschließlich mit pflanzlicher Nahrung gefüttert werden, können anfälliger für SMS sein. Die Ergänzung einer rein pflanzlichen Fütterung mit Milchpulver (das reich an Serin ist), Kasein oder Serin wird empfohlen, da es zu einem Anstieg des Blutzuckerspiegels führt.
Gestörte Skelettintegrität
Diese Störung ist nicht auf das erhöhte Körpergewicht der Masthähnchen zurückzuführen, da diese in der Lage sind, Gewichte zu tragen, die ihr eigenes Körpergewicht bei weitem übersteigen. Vielmehr liegt es daran, dass sich der Schwerpunkt des Tieres durch das stärkere Wachstum der Brustmuskulatur nach vorne verschiebt und die Beine weiter auseinander bewegt werden, was einen Torsionsdruck auf den Oberschenkelknochenkopf ausübt. Neben den Problemen, die schon auf dem landwirtschaftlichen Betrieb auftreten, kommt es zusätzlich noch zu Komplikationen bei der maschinellen Verarbeitung im Schlachthof.
Ursachen für gestörte Skelettintegrität können eine unausgewogene Nährstoffversorgung, wie beispielsweise ein Überschuss an Chlorid, oder eine Infektion mit Bakterien, Viren und insbesondere Mykoplasmen sein.
Tibiadyschondroplasie (TD)
TD wird durch eine abnormale Knorpelentwicklung verursacht. Eine gestörte Gefäßversorgung verhindert die Mineralisierung. TD ist durch eine Vergrößerung des Sprunggelenks, verdrehte Mittelfußknochen und verrutschte Sehnen gekennzeichnet. Eine niedrige Elektrolytkonzentration (<200 MEq), hoher Chloridgehalt (>0,3 %) oder ein niedriges Ca:P- bzw. ein hohes P:Ca-Verhältnis können TD auslösen. Durch die Ergänzung der Ernährung mit Mangan und Cholin lässt sich das Problem weitestgehend beseitigen.
Perosis
Die Krankheit wird heute häufig als Chondrodystrophie bezeichnet. Die klassische Ursache ist Mangan- oder Cholinmangel, sie kann jedoch auch durch den Mangel an anderen B-Vitaminen auftreten. Wie bei TD können einige Getreidebegasungsmittel den Krankheitsverlauf verschlimmern.
Verdrehter Rücken oder Kinky Back-Syndrom
Bei dem auch als Spondylolisthesis bekannten Syndrom handelt es sich nicht wirklich um eine Stoffwechselstörung, da die häufigste Ursache eine Enterokokken-Infektion ist. Hühner mit Kinky Back-Syndrom sitzen häufig auf ihrem Schwanz, strecken ihre Füße nach außen oder lassen sie auf eine Seite ihres Körpers fallen. Wenn die Vögel aufgrund dieser Krankheit nicht mehr laufen können, sind sie nicht mehr in der Lage, selbständig Nahrung oder Wasser zu erreichen und es besteht die Gefahr, zu verhungern. Es gibt keine Behandlung für diese Krankheit.
Magenerosion und Drüsenmagen
Obwohl Läsionen im Muskelmagen sehr häufig sind, geht Dr. Leeson von einer Überbewertung ihrer Bedeutung aus. Muskelmagenerkrankungen kommen bei Legehennen und noch häufiger bei Masthühnern vor.
Der Zugang zu Grit und die Einbeziehung von mindestens 20 % Getreidepartikeln mit einer Größe über 1 mm in die Fütterung wirken sich positiv auf die Entwicklung und Funktion des Muskelmagens aus. Außerdem mindern sie Häufigkeit und Schwere von Muskelmagenverletzungen bei Geflügel. Auf Struktur und Funktion des Muskelmagens hat die Aufnahme nichtlöslicher Ballaststoffe nachweislich starke Auswirkungen. Die Zugabe von mindestens 3 % grobgemahlener Rohfaser zum Futter erhöhte das relative Gewicht des Muskelmagens und senkte den pH-Wert dessen Inhalts, was auf eine vorbeugende Wirkung von Rohfaser schließen lässt.
Mit einer Magenerosion wird häufig der Vormagen, ein sehr großes Organ, in Verbindung gebracht. Bei einer Erkrankung der Vormagendrüsen kommt es zu einer geringeren Sekretion von Salzsäure und Enzymen. Dadurch gelangt mehr unverdaute Nahrung in den Darm, wo sie als Nährboden für Krankheitserreger dienen und Verdauungsinfektionen auslösen kann.
Brustmuskeldefekte
Brustmuskeldefekte stellen weder für das Geflügel und für die Effizienz/Ökonomie des Wachstums noch für die Lebensmittelsicherheit ein Problem dar. Erst in der Primär- und Sekundärverarbeitung und für die Akzeptanz beim Verbraucher wird es zum Thema. Durch das schnelle Muskelwachstum und die vergrößerten Muskelzellen verkleinert sich der Abstand zwischen den Muskelfasern. Das führt zu einer eingeschränkten Blutversorgung der Muskeln, die damit nicht mehr den gewünschten Sauerstoffgehalt erreichen können.
White Striping
Weiße Streifen sind ein Qualitätsmanko bei Hähnchenbrustfleisch. Dabei handelt es sich um Fettablagerungen im Muskel, die während des Wachstums und der Entwicklung des Tieres entstehen. „Es ist wie die Marmorierung von rotem Fleisch“, scherzte Dr. Leeson „man sollte es als marmoriertes Huhn bewerben – wie Wagyu-Rindfleisch“. Da Hypoxie mit der Bildung weißer Streifen einhergeht, könnte man sich überlegen, ob eine Arginin-Ergänzung die Gefäßerweiterung fördern und so die Muskeln mit mehr Sauerstoff versorgen könnte.
Wooden Breast (WB) -“Holzbrust“
WB ist ein immer häufiger auftretender Qualitätsmangel. Makroskopisch ist es durch tastbare harte, blasse, kammartige Ausbuchtungen am kaudalen Ende gekennzeichnet, die in Verbindung mit klarer, zäher Flüssigkeit, kleinen Blutungen und weißen Streifen, einzeln oder zusammen, auftreten können. Hauptursache ist das schnelle Wachstum und der hohe Brustfleischertrag. Über die Fütterung oder das Management gibt es gegen diese Art von Muskelveränderung keine Lösung.
Wooden Breast kommt häufig bei männlichen Broilern mit einem Körpergewicht von über 2,5 kg vor und die Häufigkeit nimmt tendenziell mit der Größe des Brustfilets zu.
Während sich die Häufigkeit von Wooden Breast erhöht, ist White Striping tendenziell auf dem Rückzug. Aufgrund optischer Mängel und der harten, zähen Textur genießen WB-Filets nur eine geringe Akzeptanz beim Verbraucher. Sie werden in der Regel abgewertet und für die Hackfleischproduktion verwendet.
Durch die Reduzierung von oxidativem Stress und eine höhere Sauerstoffversorgung der Muskelzellen, die es ihnen ermöglicht sehr schnell und ohne Fleischeinbuße zu wachsen, kann die Häufigkeit von Wooden Breast reduziert werden.
Die EW Nutrition Poultry Academy fand Anfang September 2023 in Jakarta and Manila statt. Dr. Steve Leeson, anerkannter Experte für Geflügelernährung und -produktion und mit nahezu 50 Jahren Erfahrung in der Geflügelindustrie, war der geschätzte Hauptredner der Veranstaltung.
Dr. Leeson erlangte seinen Doktortitel in Geflügelernährung im Jahr 1974 an der Universität von Nottingham. 38 Jahre lang war er Professor des Lehrstuhls für Tier- und Geflügelwissenschaft an der Universität Guelph in Kanada, wo er seit 2014 als emeritierter Professor noch tätig ist. Der hervorragende Autor kann mehr als 400 Veröffentlichungen in referierten Fachzeitschriften und 6 Bücher über unterschiedlichste Aspekte aus Geflügelernährung und -management vorweisen. Er war Gewinner des “American Feed Manufacturer’s Association Nutrition Research Award” (1981), des “Canadian Society of Animal Science Fellowship Award” (2001) und des Novus Lifetime Achievement Award in Poultry Nutrition (2011).
Darmgesundheitsmanagement: eine enorme Herausforderung in antibiotikafreier Hähnchenmast
Von Dr. Ajay Bhoyar, Global Technical Manager Poultry, EW Nutrition
Die Darmgesundheit ist speziell in der antibiotikafreien (ABF) Produktion essenziell, da sie eine entscheidende Rolle für die allgemeine Gesundheit und das Wohlbefinden der Tiere spielt. Antibiotika werden seit langem zur Vorbeugung und Behandlung von Tierkrankheiten eingesetzt, aber ihr übermäßiger Einsatz hat zur Entwicklung von antibiotikaresistenten Bakterien geführt. Viele Landwirte und Erzeuger stellen daher auf antibiotikafreie Produktionsmethoden um. Diese Umstellung stellt eine große Herausforderung dar, da die Erhaltung der Darmgesundheit ohne Antibiotika schwierig sein kann. Es ist jedoch nicht unmöglich.
Eine der schwierigsten Aufgaben in der antibiotikafreien Produktion ist die Verhinderung von bakteriellen Infektionen im Darm. Das Darmmikrobiom spielt eine ausschlaggebende Rolle für das Immunsystem und das Allgemeinbefinden der Tiere. Wenn das Gleichgewicht der Mikroben im Darm gestört ist (Dysbiose), kann dies zu einer schlechten Nährstoffaufnahme führen, die folglich die Leistung der Tiere wie Futterverwertung und Gewichtszunahme beeinträchtigt. Wenn Landwirte und Erzeuger keine Antibiotika verwenden wollen, müssen sie auf andere Methoden zurückgreifen, um ein gesundes Darmmikrobiom zu erhalten.
Reduzierung von Antibiotika – ein wichtiger globaler Trend
In den letzten Jahren tendieren Geflügelproduzenten immer mehr dazu, den Einsatz von Antibiotika zu reduzieren, um die Gesundheit von Mensch und Tier zu fördern und die Nachhaltigkeit ihrer Betriebe zu verbessern. Ausschlaggebend dafür ist die Besorgnis über die Entwicklung antibiotikaresistenter Bakterien, die möglichen Auswirkungen auf die menschliche Gesundheit und die Nachfrage der Verbraucher nach Fleisch, das ohne Antibiotika erzeugt wurde. In vielen Ländern gibt es inzwischen Vorschriften, die den Einsatz von Antibiotika in der Lebensmittel- und Tierproduktion begrenzen.
Herausforderungen für die antibiotikafreie Geflügelproduktion (ABF)
Krankheitsbekämpfung. In der antibiotikafreien Geflügelproduktion müssen Landwirte auf alternative Methoden zur Bekämpfung und Vorbeugung von Krankheiten zurückgreifen, z. B. verstärkte Biosicherheitsmaßnahmen. Dies kann arbeitsintensiv und kostspielig sein.
Höhere Sterblichkeitsraten. Ohne Antibiotika kann es in Geflügelbeständen zu höheren Sterblichkeitsraten aufgrund von Krankheitsausbrüchen und anderen Gesundheitsproblemen kommen. Dies kann zu finanziellen Verlusten für die Landwirte und zu einem geringeren Angebot an Geflügelprodukten für die Verbraucher führen.
Fütterung. Antibiotische Wachstumsförderer (AGPs – aus dem Englischen: Antibiotic Growth Promoters) werden häufig bei Geflügel zur Wachstumsförderung und zur Verhinderung von Darmerkrankungen eingesetzt. Ohne AGPs müssen Geflügelproduzenten alternative Wege finden, um die erwartete Produktionsleistung zu erreichen.
Erhöhte Kosten. Die antibiotikafreie Geflügelproduktion kann teurer sein als konventionelle Produktionsmethoden, da die Landwirte in zusätzliche Ställe, Ausrüstung, Arbeitskräfte usw. investieren müssen.
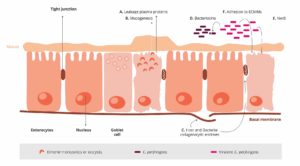
Wenn AGPs nicht mehr eingesetzt werden, wird es wahrscheinlich zu Veränderungen der mikrobiellen Zusammensetzung im Darm kommen. Es besteht die Hoffnung, dass Strategien wie Programme zur Vorbeugung von Infektionskrankheiten und die Verwendung von nicht antibiotischen Alternativen die möglichen negativen Folgen des Antibiotikaentzugs auf den Geflügelbestand minimieren (Yegani und Korver, 2008).
Darmgesundheit – der Schlüssel zu allgemeiner Gesundheit
Ein gesunder Magen-Darm-Trakt ist wichtig für die Erfüllung des maximalen Produktionspotenzials. Darmgesundheit bei Geflügel beschreibt den Allgemeinzustand und die Funktion des Magen-Darm-Trakts und umfasst das Gleichgewicht der nützlichen Bakterien, die Integrität der Darmschleimhaut und die Fähigkeit, Nährstoffe zu verdauen und aufzunehmen. Darmgesundheit ist wichtig, um die Gesundheit insgesamt und das Wohlbefinden der Tiere aufrecht zu erhalten. Ein gesunder Darm trägt zur Verbesserung der Futterverwertung, der Nährstoffaufnahme und der generellen Immunität der Tiere bei.
Der Darm beherbergt mehr als 640 verschiedene Arten von Bakterien und über 20 verschiedene Hormone. Er verdaut und absorbiert den Großteil der Nährstoffe und benötigt dafür fast ein Viertel des Energieverbrauchs des Körpers. Er ist auch das größte Immunorgan des Körpers (Kraehenbuhl und Neutra, 1992). Der Begriff “Darmgesundheit” ist daher sehr komplex und umfasst die makro- und mikrostrukturelle Integrität des Darms, das Gleichgewicht der Mikroflora und den Zustand des Immunsystems (Chot, 2009).
Immunität bei Geflügel kommt vom Darm
Der Darm ist ein wichtiger Bestandteil des Immunsystems, denn er ist die erste Verteidigungslinie gegen Krankheitserreger, die über das Verdauungssystem in den Körper gelangen. Hühner verfügen über ein spezialisiertes Immunsystem im Darm, das so genannte darmassoziierte lymphatische Gewebe (GALT – gut-associated lymphoid tissue), das dazu beiträgt, potenzielle Krankheitserreger zu erkennen und auf sie zu reagieren. Zum GALT gehören die Peyer’schen Plaques (Gruppen von Immunzellen, die sich in der Darmwand befinden) sowie die Darm-assoziierten Lymphozyten (GAL – gut associated lymphocytes), die im gesamten Darm zu finden sind. Diese Immunzellen sind dafür verantwortlich, Krankheitserreger, die in den Darm eindringen, zu erkennen und zu bekämpfen.
Die darmvermittelte Immunreaktion bei Hühnern umfasst mehrere Mechanismen, darunter die Aktivierung von Immunzellen, die Produktion von Antikörpern und die Freisetzung von Entzündungsmediatoren. Das GALT und die GALs spielen bei dieser Reaktion eine entscheidende Rolle, indem sie Krankheitserreger identifizieren, auf sie reagieren und weitere Immunzellen zur Bekämpfung der Infektion aktivieren.
Auch das Darmmikrobiom spielt bei der darmvermittelten Immunität von Hühnern eine entscheidende Rolle. Es besteht aus einer äußerst vielfältigen Gemeinschaft von Mikroorganismen, die einen bedeutenden Einfluss auf die Immunantwort haben können. So können bestimmte nützliche Bakterien dazu beitragen, die Immunreaktion zu stimulieren und den Darm vor Krankheitserregern zu schützen.
Insgesamt arbeiten Darmmikrobiom, GALT und GALs zusammen, um eine für Krankheitserreger feindliche Umgebung zu schaffen, die aber gleichzeitig das Wachstum und die Gesundheit nützlicher Mikroorganismen fördert.
Dysbiose/Dysbakteriose beeinträchtigt die Leistung
Bei einer Dysbiose handelt es sich um ein Ungleichgewicht in der Darmmikrobiota aufgrund einer Belastung des Darms. Dieses Ungleichgewicht kann zu nasser und verkrusteter Einstreu führen und, bei längerem Kontakt der Tiere damit, Pododermatitis (Geschwüren an den Füßen) und Verbrennungen an den Sprunggelenken verursachen. Tierschutzprobleme und eine Abwertung des Schlachtkörpers können die Folge sein (Bailey, 2010). Abgesehen von diesen Auswirkungen ergeben sich die größten wirtschaftlichen Verluste aus den verringerten Wachstumsraten, der schlechteren Futterverwertung und den erhöhten Kosten für tierärztliche Behandlungen. Durch eine Dysbiose können Kokzidiose-Infektionen und andere Darmerkrankungen verschlimmert werden. Tiere mit einer Dysbiose weisen im Allgemeinen hohe Konzentrationen an Clostridien auf, die noch mehr Toxine erzeugen und zu einer nekrotischen Enteritis führen können.
Fig.1: Dysbiose – das Ergebnis einer Belastung des Mikrobioms der Tiere. Quelle: Charisse Petersen und June L. Round.2014
Es wird angenommen, dass sowohl nicht-infektiöse als auch infektiöse Faktoren bei der Dysbakteriose eine Rolle spielen können (DeGussem, 2007). Jede Veränderung der Futtermittel und Futtermittelrohstoffe sowie deren physikalische Qualität hat Einfluss auf das Gleichgewicht der Darmmikrobiota. In der Geflügelproduktion gibt es problematische Phasen, in denen die Tiere besonderen Belastungen ausgesetzt sind, z. B. bei Futterwechsel, Impfungen, Behandlungen, Transport usw. Während dieser Zeiten kann sich das Darmmikrobiom verändern, und in einigen Fällen kann es bei suboptimalem Management zu einer Dysbakteriose kommen.
Zu den infektiösen Erregern, die möglicherweise eine Rolle bei Dysbakteriose spielen, gehören Mykotoxine, Eimeria spp, Clostridium perfringens und andere Bakterien, die toxische Stoffwechselprodukte produzieren.
Faktoren, die die Darmgesundheit beeinträchtigen
Die Faktoren, die die Darmgesundheit von Masthähnchen beeinflussen, lassen sich wie folgt zusammenfassen:
Futter- und Wasserqualität: Form, Art und Qualität des Futters, das Masthähnchen verfüttert wird, können sich erheblich auf die Darmgesundheit auswirken. Die ständige Verfügbarkeit von kühlem und sauberem Trinkwasser ist entscheidend für eine optimale Produktionsleistung.
Stress: Stressige Bedingungen, wie hohe Umgebungstemperaturen oder schlechte Belüftung, können zu einem Ungleichgewicht im Darmmikrobiom und einem erhöhten Krankheitsrisiko führen.
Mikrobielle Belastung: Eine Störung des Mikrobioms und Probleme mit der Darmgesundheit können auftreten, wenn die Tiere Krankheitserregern oder anderen schädlichen Bakterien ausgesetzt sind.
Immunsystem: Ein robustes Immunsystem ist wichtig für die Gesunderhaltung des Darms, da es dazu beiträgt, unmäßiges Wachstum schädlicher Bakterien zu verhindern und die Vermehrung nützlicher Bakterien zu fördern.
Hygiene: Eine saubere und von Krankheitserregern freie Umgebung ist für die Erhaltung der Darmgesundheit von Masthähnchen von entscheidender Bedeutung, da sich Bakterien und andere Krankheitserreger leicht ausbreiten und das Darmmikrobiom stören können.
Management-Praktiken: Richtige Haltungspraktiken wie passendes Fütterungs- und Tränkewasser- sowie ein funktionierendes Einstreu-Management können dazu beitragen, die Darmgesundheit zu erhalten und Darmprobleme zu vermeiden.
Fig. 2. Schlüsselfaktoren, die die Darmgesundheit von Masthähnchen beeinflussen
Zentrale Ansätze für ein Darmgesundheitsmanagement ohne Antibiotika
Zwei Ansätze für das antibiotikafreie Management der Darmgesundheit bei Geflügel sind außerordentlich erfolgreich.
Geeignete Fütterungs- und Managementpraktiken
Für die ABF-Geflügelproduktion ist es wichtig, dass die Tiere Zugang zu sauberem Wasser und hochwertigem Futter haben und in einer stressfreien Umgebung sind. Eine ausgewogene Ration in Bezug auf Eiweiß, Energie sowie essenzielle Vitamine und Mineralien ist für die Erhaltung der Darmgesundheit unerlässlich.
Die Umgebung, in der das Geflügel gehalten wird, spielt eine wichtige Rolle bei der Erhaltung der Darmgesundheit. Angemessene Hygiene und Belüftung sowie die richtige Temperatur und Luftfeuchtigkeit sind entscheidend, um die Ausbreitung von Krankheiten und Infektionen einzudämmen. Außerdem ist eine strikte Umsetzung strenger Biosicherheitsmaßnahmen alternativlos.
Durch die frühzeitige Erkennung und Behandlung von Krankheiten kann verhindert werden, dass diese sich zu ernsthaften Problemen entwickeln und damit die Rentabilität der ABF-Produktion beeinträchtigen. Es ist wichtig, sich die Tiere immer wieder genau hinsichtlich Krankheitsanzeichen wie Durchfall, verminderte Wasser- und Futteraufnahme anzusehen.
Darmgesundheitsfördernde Futtermittelzusätze
Ein weiteres Vorgehen zum Erhalt der Darmgesundheit in einer antibiotikafreien Geflügelproduktion ist die Verwendung von Futtermittelzusätzen, die die Darmgesundheit unterstützen. In der Tierproduktion wird eine Vielzahl von Futtermittelzusätzen zur Förderung der Darmgesundheit eingesetzt, darunter sekundäre Pflanzenstoffe/ätherische Öle, organische Säuren, Probiotika, Präbiotika, exogene Enzyme usw., sowohl einzeln als auch in Kombination. Insbesondere die kosteneffizienten phytogenen Futtermittelzusätze (PFZ) haben mit bereits vielfach nachgewiesener Wirkung auf die Darmgesundheit von Masthühnern an Interesse gewonnen.
Sekundäre Pflanzenstoffe und ätherische Öle (oft auch als Phytogene, Phytochemikalien oder Phytomoleküle bezeichnet) sind biologisch aktive Verbindungen, die in jüngster Zeit als Futtermittelzusätze in der Geflügelproduktion Interesse wecken. Sie verbessern die Futterverwertung durch Förderung der Produktion von Verdauungssekreten und der Nährstoffaufnahme. Dies trägt dazu bei, die Belastung des Darms mit Pathogenen und Radikalen sowie die mikrobielle Belastung des Immunsystems der Tiere zu reduzieren (Abdelli et al. 2021).
Pflanzenextrakte – Ätherische Öle / Phytomoleküle
Phytogene Stoffe sind natürliche, in Pflanzen vorkommende Verbindungen. Viele dieser Stoffe haben nachweislich antimikrobielle Eigenschaften, d. h. sie können das Wachstum von Mikroorganismen wie Bakterien, Viren und Pilzen hemmen oder diese abtöten. Beispiele für sekundäre Pflanzenstoffe mit antimikrobiellen Eigenschaften sind Verbindungen, die in Knoblauch, Thymian und Teebaumöl vorkommen. Ätherische Öle sind rohe Pflanzenextrakte (Blüten, Blätter, Wurzeln, Früchte usw.), während Phytomoleküle die aktiven Inhaltsstoffe von ätherischen Ölen oder anderen Pflanzenmaterialien sind und eindeutig als Wirkstoffe definiert sind. Ätherische Öle sind wichtige aromatische Bestandteile von Kräutern und Gewürzen und werden als natürliche Alternativen zum Ersatz von antibiotischen Wachstumsförderern (AGP – englisch: antibiotic growth promoters) im Geflügelfutter verwendet. Zu den besonderen Wirkungen von ätherischen Ölen gehören die Appetitanregung, Förderung der Enzymsekretion bei der Verdauung und die Aktivierung der Immunantwort (Krishan und Narang, 2014).
Eine Vielzahl von Kräutern und Gewürzen (u. a. Thymian, Oregano, Zimt, Rosmarin, Majoran, Schafgarbe, Knoblauch, Ingwer, grüner Tee, Schwarzkümmel und Koriander) sowie ätherischen Ölen (aus Thymian, Oregano, Zimt, Knoblauch, Anis, Rosmarin, Zitrusfrüchten, Nelken und Ingwer) wurden einzeln oder in Kombination als potenzielle AGP-Alternativen für Geflügel verwendet (Gadde et al., 2017).
Fig. 3: Futtermittelzusatz auf Phytomolekülbasis übertrifft AGPs durch verbesserte Leistung bei Masthähnchen (42-tägige Feldstudie)
Eine der vorrangigen Wirkweisen von ätherischen Ölen hängt mit ihren antimikrobiellen Eigenschaften zusammen, die die Bekämpfung potenzieller Krankheitserreger ermöglichen (Mohammadi und Kim, 2018).
Phytomolekül-Mischung
Clostridium perfringens
Enterococcus cecorum
Enterococcus hirae
Escherichia coli
Salmonella typhimurium
Staphylococcus aureus
Ventar D
1250
2500
5000
2500
5000
2500
Abb. 4: Aktivität eines Futtermittelzusatzes auf Phytomolekülbasis (Ventar D) gegen enteropathogene Bakterien (MHK-Wert – minimale Hemmkonzentration – in ppm)
Phytomoleküle wie z.B. Flavonoide, Polyphenole, Carotinoide und Terpene haben nachweislich entzündungshemmende Eigenschaften. Sie werden unter anderem durch Hemmung der Aktivität von entzündungsfördernden Enzymen und Molekülen hervorgerufen. So blockieren Polyphenole nachweislich die Aktivität des Nuklearfaktors Kappa B (NF-kB), eines Transkriptionsfaktors, der eine Schlüsselrolle bei der Regulierung von Entzündungen spielt.
Phytomoleküle haben auch antioxidative Eigenschaften, die helfen können, Zellen vor Schäden durch reaktive Sauerstoffspezies (ROS) und andere reaktive Moleküle zu schützen, die zu Entzündungen beitragen können. Es wird dementsprechend vorgeschlagen, Pflanzenextrakte als Antioxidantien in Tierfutter zu verwenden, um die Tiere vor oxidativen, durch freie Radikale verursachten, Schäden zu schützen. Phenolische OH-Gruppen in Thymol, Carvacrol und anderen Pflanzenextrakten wirken als Wasserstoffdonator für Peroxyradikale, die während des ersten Schritts der Lipidoxidation entstehen, und verzögern so die Bildung von Hydroxylperoxid (Farag et al., 1989, Djeridane et al., 2006). Thymol und Carvacrol haben eine stark-antioxidative Wirkung (Yanishlieva et al., 1999) und unterdrücken Berichten zufolge die Lipidperoxidation (Hashemipour et.al. 2013).
Insgesamt geht man davon aus, dass die entzündungshemmenden Wirkungen von Phytomolekülen auf die Kombination aus ihrer Fähigkeit, die Aktivität entzündungsfördernder Enzyme und Moleküle zu hemmen und das Immunsystem zu modulieren sowie ihren antioxidativen Eigenschaften zurückzuführen sind. Pflanzenextrakte (z. B. Carvacrol, Zimtaldehyd, Eugenol usw.) hemmen die Produktion von pro-inflammatorischen Zytokinen und Chemokinen aus Endotoxin-stimulierten Immun- und Epithelzellen (Lang et al., 2004, Lee et al., 2005, Liu et al., 2020). Es gibt Hinweise darauf, dass die entzündungshemmende Wirkung teilweise durch die Blockierung des NF-κB-Aktivierungswegs vermittelt wird (Lee et al., 2005).
Fig. 5: Entzündungshemmende Eigenschaft eines Futtermittelzusatzes auf Phytomolekülbasis (Ventar D) – verringerte Aktivität entzündlicher Zytokine
Angemessener Schutz der Phytomoleküle ist der Schlüssel zu optimalen Ergebnissen
Es hat sich gezeigt, dass etliche phytogene Verbindungen größtenteils im oberen Teil des Verdauungstrakts absorbiert werden, was bedeutet, dass die meisten von ihnen ohne angemessenen Schutz nicht im unteren Teil des Darms, wo sie ihre Hauptfunktionen ausüben sollten, ankommen würden (Abdelli et al. 2021). Der Zusatz einer Mischung aus enkapsulierten ätherischen Ölen zu Masthähnchenfutter brachte bessere Ergebnisse als der Zusatz der phytogenen Futterzusätze (PFZ) in pulverisierter, nicht geschützter Form (Hafeez et al. 2016). Neuartige Verabreichungstechnologien wurden entwickelt, um PFZs vor Abbau und Oxidation während der Futtermittelverarbeitung und Lagerung zu schützen, die Handhabung zu erleichtern, eine langsamere Freisetzung zu ermöglichen und auf den unteren Verdauungstrakt abzuzielen (Starčević et al. 2014). Die speziellen Schutztechniken, die bei der kommerziellen Herstellung einer Mischung aus ätherischen Ölen/Phytomolekülen eingesetzt werden, sind entscheidend, um die gesteckten Ziele mit bemerkenswerter Beständigkeit zu erreichen.
Fig. 6: Stabilität von einem auf Phytomolekülen basierenden Futtermittelzusatz (Ventar D) bei der Pelletierung mit hohen Temperaturen und längeren Konditionierungszeiten
Mischung aus Phytomolekülen optimiert die Produktionsleistung
Der Verzicht auf Antibiotika in der Geflügelproduktion kann ein Problem für die Kontrolle der Mortalität und die Aufrechterhaltung der Produktionsleistung der Tiere darstellen. Es hat sich gezeigt, dass phytogene Futterzusätze aufgrund ihrer antimikrobiellen, entzündungshemmenden, antioxidativen und verdauungsfördernden Eigenschaften dieProduktionsleistung von Geflügel verbessern. Die verbesserte Nährstoffverdaulichkeit durch phytogene Futtermittelzusätze (PFZ) könnte auf deren Fähigkeit zurückzuführen zu sein, Appetit, Speichelsekretion, Darmschleimproduktion, Gallensäuresekretion und die Aktivität von Verdauungsenzymen wie Trypsin und Amylase zu stimulieren sowie die Darmmorphologie positiv zu beeinflussen (Oso et al. 2019). Ätherische Öle werden in der Geflügelproduktion als Wachstumsförderer angesehen und zeigen starke antimikrobielle und kokzidiostatische Aktivitäten (Zahi et al., 2018). PFZs beeinflussen Gewichtszunahme und Futterverwertung bei Geflügel in positiver Weise (Khattak et al. 2014, Zhang et el. 2009).
Fig. 7: Futtermittelzusatz auf Basis von Phytomolekülen verbessert im Feldversuchbei Masthähnchen die Futterverwertung und senkt die Mortalität
Schlussfolgerung
Zusammenfassend lässt sich sagen, dass das Darmgesundheitsmanagement eine enorme Herausforderung in der ABF-Hähnchenproduktion darstellt. Dieser Herausforderung muss sich gestellt werden, um das Wohlbefinden der Tiere und damit eine optimale Leistung zu sichern. Der Einsatz von Antibiotika als Präventivmaßnahme in der Masthähnchenproduktion ist immer noch weit verbreitet, doch angesichts der steigenden Nachfrage nach antibiotikafreien Produkten müssen alternative Methoden zur Erhaltung der Darmgesundheit eingesetzt werden. Dazu gehören die Verwendung von Futtermittelzusätzen, die die Darmgesundheit fördern, und angemessene Managementpraktiken, wie die Umsetzung von Biosicherheitsmaßnahmen, die Aufrechterhaltung optimaler Umweltbedingungen, ausreichend Platz und angemessene Belüftung sowie die Reduzierung von Stress. Leider jedoch gibt es keine Einheitslösung für das Darmgesundheitsmanagement in der ABF-Hähnchenmast. Wichtig ist, die Darmgesundheit der Tiere kontinuierlich zu überwachen und einzuschätzen und gegebenenfalls Anpassungen vorzunehmen. Darüber hinaus sollten Forschung und Entwicklung in diesem Bereich gefördert werden, um neue und innovative Wege zur Erhaltung der Darmgesundheit in der ABF-Masthähnchenproduktion zu finden.
Insgesamt ist das Darmgesundheitsmanagement eine erhebliche Herausforderung, die einen vielschichtigen Ansatz und kontinuierliche Überwachung und Management erfordert. Durch die Umsetzung geeigneter Strategien und den Einsatz neuer Technologien können Geflügelhalter Gesundheit und Wohlbefinden ihrer Tiere sicherstellen und gleichzeitig die wachsende Nachfrage nach antibiotikafreien Produkten nachhaltig befriedigen.
Literatur:
Abdelli N, Solà-Oriol D, Pérez JF. Phytogenic Feed Additives in Poultry: Achievements, Prospective and Challenges. Animals (Basel). 2021 Dec 6;11(12):3471.
BaileyR. A. 2010. Intestinal microbiota and the pathogenesis of dysbacteriosis in broiler chickens. PhD thesis submitted to the University of East Anglia. Institute of Food Research, United Kingdom
Choct M. Managing gut health through nutrition. British Poultry Science Volume 50, Number 1 (January 2009), pp. 9—15.
De Gussem M, “Coccidiosis in Poultry: Review on Diagnosis, Control, Prevention and Interaction with Overall Gut Health,” Proceedings of the 16th European Symposium on Poultry Nutrition, Strasbourg, 26-30 August, 2007, pp. 253-261.
Dorman, H.J. and S.G. Deans. Antimicrobial agents from plants: antibacterial activity of plant volatile oils. J Appl Microbiol, 88 (2000), pp. 308-316
Djeridane A., M. Yousfi M, Nadjemi B, Boutassouna D., Stocker P., Vidal N. Antioxidant activity of some Algerian medicinal plant extracts containing phenolic compounds. Food Chem, 97 (2006), pp. 654-660
Farag R. S., Daw Z.Y., Hewedi F.M., El-Baroty G.S.A. Antimicrobial activity of some Egyptian spice essential oils. J Food Prot, 52 (1989), pp. 665-667
Gadde U., Kim W.H., Oh S.T., Lillehoj H.S. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: A review. Anim. Health Res. Rev. 2017; 18:26–45.
Guo, F.C., Kwakkel, R.P., Williams, B.A., Li, W.K., Li, H.S., Luo, J.Y., Li, X.P., Wei, Y.X., Yan, Z.T. and Verstegen, M.W.A., 2004. Effects of mushroom and herb polysaccharides, as alternatives for an antibiotic, on growth performance of broilers. British Poultry Science, 45(5), pp.684-694.
Hafeez A., Männer K., Schieder C., Zentek J. Effect of supplementation of phytogenic feed additives (powdered vs. encapsulated) on performance and nutrient digestibility in broiler chickens. Poult. Sci. 2016; 95: 622–629.
Hammer K.A., Carson C.F., Riley T.V. Antimicrobial activity of essential oils and other plant extracts. J Appl Microbiol, 86 (1999), pp. 985-990
Hashemipour H, Kermanshahi H, Golian A, Veldkamp T. Effect of thymol and carvacrol feed supplementation on performance, antioxidant enzyme activities, fatty acid composition, digestive enzyme activities, and immune response in broiler chickens. Poultry Science. Volume 92. Issue 8. 2013, Pp 2059-2069
Khattak F., Ronchi A., Castelli P., Sparks N. Effects of natural blend of essential oil on growth performance, blood biochemistry, cecal morphology, and carcass quality of broiler chickens. Poult. Sci. 2014; 93: 132–137
Kraehenbuhl, J.P. & Neutra, M.R. (1992) Molecular and cellular basis of immune protection of mucosal surfaces. Physiology Reviews, 72: 853–879.Krishan and Narang J. Adv. Vet. Anim. Res., 1(4): 156-162, December 2014
Lang A., Lahav M., Sakhnini E, Barshack I., Fidder H. H., Avidan B. Allicin inhibits spontaneous and TNF-alpha induced secretion of proinflammatory cytokines and chemokines from intestinal epithelial cells. Clin Nutr, 23 (2004), pp. 1199-1208
Lee S.H., Lee S.Y., Son D.J., Lee H., Yoo H.S., Song S. Inhibitory effect of 2′-hydroxycinnamaldehyde on nitric oxide production through inhibition of NF-kappa B activation in RAW 264.7 cells Biochem Pharmacol, 69 (2005), pp. 791-799
Liu, S., Song, M., Yun, W., Lee, J., Kim, H. and Cho, J., 2020. Effect of carvacrol essential oils on growth performance and intestinal barrier function in broilers with lipopolysaccharide challenge. Animal Production Science, 60(4), pp.545-552.
Mitsch, P., Zitterl-Eglseer, K., Köhler, B., Gabler, C., Losa, R. and Zimpernik, I., 2004. The effect of two different blends of essential oil components on the proliferation of Clostridium perfringens in the intestines of broiler chickens. Poultry science, 83(4), pp.669-675.
Mohammadi Gheisar M., Kim I.H. Phytobiotics in poultry and swine nutrition—A review. Ital. J. Anim. Sci. 2018; 17: 92–99.
Oso A.O., Suganthi R.U., Reddy G.B.M., Malik P.K., Thirumalaisamy G., Awachat V.B., Selvaraju S., Arangasamy A., Bhatta R. Effect of dietary supplementation with phytogenic blend on growth performance, apparent ileal digestibility of nutrients, intestinal morphology, and cecal microflora of broiler chickens. Poult. Sci. 2019; 98: 4755–4766
Oviedo-Rondón, Edgar O., et al. “Ileal and caecal microbial populations in broilers given specific essential oil blends and probiotics in two consecutive grow-outs.” Avian Biology Research 3.4 (2010): 157-169.
Petersen C. and June L. Round. Defining dysbiosis and its influence on host immunity and disease. Cellular Microbiology (2014)16(7), 1024–1033
Starčević K., Krstulović L., Brozić D., Maurić M., Stojević Z., Mikulec Ž., Bajić M., Mašek T. Production performance, meat composition and oxidative susceptibility in broiler chicken fed with different phenolic compounds. J. Sci. Food Agric. 2014; 95: 1172–1178.
Yanishlieva, N.V., Marinova, E.M., Gordon, M.H. and Raneva, V.G., 1999. Antioxidant activity and mechanism of action of thymol and carvacrol in two lipid systems. Food Chemistry, 64(1), pp.59-66.
Yegani, M. and Korver, D.R., 2008. Factors affecting intestinal health in poultry. Poultry science, 87(10), pp.2052-2063.
Zhai, H., H. Liu, Shikui Wang, Jinlong Wu and Anna-Maria Kluenter. “Potential of essential oils for poultry and pigs.” Animal Nutrition 4 (2018): 179 – 186.
Zhang G.F., Yang Z.B., Wang Y., Yang W.R., Jiang S.Z., Gai G.S. Effects of ginger root (Zingiber officinale) processed to different particle sizes on growth performance, antioxidant status, and serum metabolites of broiler chickens. Poult. Sci. 2009; 88: 2159–2166.
Moderne phytogene Futterzusatzstoffe – auf die Verkapselung kommt’s an
Henning Gerstenkorn & Ruturaj Patil, EW Nutrition
Sekundären Pflanzenextrakten wurden eine verbesserte Verdauung, positive Effekte auf die Darmgesundheit, sowie ein Schutz bei oxidativem Stress in diversen wissenschaftlichen Studien der vergangenen Jahre attestiert (Hashemi and Davoodi, 2011). Ihr Einsatz als Futtermittelzusatzstoff hat sich etabliert und verschiedene Mischungen, den entsprechenden Zielstellungen angepasst, sind erhältlich. Deren Verwendung im pelletierten Futter stehen jedoch seit geraumer Zeit kritische Stimmen gegenüber. Eine unbefriedigende Reproduzierbarkeit der positiven Einflüsse auf die Leistungsparameter, insbesondere in ihrem Ausmaß, stehen im Fokus der Kritiker.
Als mögliche Ursachen werden nicht ausreichend standardisierte Rohwaren, sowie nicht kontrollierbare und ungleichmäßige Verluste der wertvollen enthaltenen Phytomoleküle während der Mischfutterherstellung diskutiert. In diesem Artikel beleuchten wir einen weiteren Parameter dessen Leistungsfähigkeit für die ökonomische und ökologische Nachhaltigkeit von phytogenen Futterzusatzstoffen entscheiden ist: die Verkapselungstechnologie.
Qualität phytogener Zusatzstoffe hängt von mehreren Faktoren ab
Das intensive Bestrebens der Nutztierhaltung, deren Bedarf antibiotisch wirksamer Arzneimittel auf ein unverzichtbares Minimum zu reduzieren, hat zu einer intensivere Nutzung von natürlichen und naturidentischen Futtermittelzusatzstoffen zur präventiven Gesunderhaltung der Nutztierbestände geführt. In den Kategorien der zootechnischen und sensorischen Zusatzstoffe sind zahlreiche Stoffe eingeordnet, die in der Humanernährung in dem Bereich der Gewürzpflanzen und Kräuter, oder in der traditionellen Medizin als Heilkräuter bekannt sind.
Die ersten verfügbaren Produkte dieser phytogenen Zusatzstoffe wurden auf einfachem Wege dem Mischfutter beigemengt. Die gewünschten Pflanzenteile wurden, ähnlich wie Gewürze und Kräuter in der Humanernährung, zerkleinert, oder zermahlen dem Premix beigemengt. Alternativ wurden flüssige Pflanzenextrakte vorab auf einen geeigneten Träger (z. B. Kieselgur) gebracht, um diese dann in den Premix einzubringen. An diesen Verfahrensweisen lassen sich zwei Gegebenheiten aufzeigen, die für die zu Beginn genannten schwierigen Reproduzierbarkeit positiver Ergebnisse verantwortlich sein können.
Variabilität aktiver Substanzen in Rohstoffen
Ein nicht zu unterschätzender Störfaktor ist die variierende Konzentration und Zusammensetzung der aktiven Substanzen in der Pflanze. Diese Zusammensetzung ist im Wesentlichen von den Standortbedingungen, wie Witterung, Boden, Lebensgemeinschaft und Erntezeitpunkt abhängen (Ehrlinger, 2007). In einem aus Thymian gewonnenen Öl können daher die Gehalte des relevanten Phenols Thymol zwischen 30 und 70 % variieren (Lindner, 1987). Diese extremen Schwankungen werden bei modernen phytogenen Zusatzstoffen durch den Einsatz naturidentischer Inhaltsstoffe vermieden.
Pelletierung setzt sensiblen Zusatzstoffen zu
Die in Diskussion stehenden Verluste der wertvollen Phytomoleküle können ebenfalls auf die natürliche Herkunft der Rohmaterialien zurückgeführt werden. Einige Phytomoleküle (z. B. Cineol) sind bereits bei niedrigen Temperaturen flüchtig. In der Hausapotheke nutzt man diesen Effekt vorwiegend bei Erkältungsprodukten, um unter Zugabe von heißem Wasser die ätherischen Öle aus Minze und Eukalyptus inhalieren zu können. Im Prozess des Pelletierens in der Mischfutterherstellung herrschen je nach Bauart Temperaturen zwischen 60 °C und bis zu 90 °C, die bis zum Abkühlungsprozess über mehrere Minuten anhalten können. Sensible Zusatzstoffe können in diesem Prozessschritt inaktiviert werden, oder sich verflüchtigen.
Eine stabile Verkapselung als Schlüssel zur Stabilität
Eine technische Lösung zur Erhaltung temperatursensibler Zusatzstoffe stellt die Ummantelung mit einer Schutzhülle dar, welche bei Enzymen bereits etabliert ist. Eine solche sogenannte Verkapselung wird auch bei phytogenen Zusatzstoffen bereits in höherwertigen Produkten erfolgreich genutzt. Die flüchtigen Substanzen sollten durch eine Ummantelung mit Fett oder Stärke geschützt werden, damit der Großteil (>70%) der Inhaltsstoffe auch nach dem Pelletieren wiedergefunden wird. Leider ist mit dieser Kapsel kein kompletter Schutz möglich, da diese einfache Schutzhülle durch mechanischen Druck beim Mahlen und Pelletieren aufgebrochen werden kann. Neue Arten der Mikroverkapselung wirken dem jedoch entgegen: Vergleichbar mit einem Schwamm, wird bei mechanischem Druck auf solch eine Kapsel nur ein kleiner Anteil der mit flüchtigen Phytomolekülen gefüllten Kammern beschädigt.
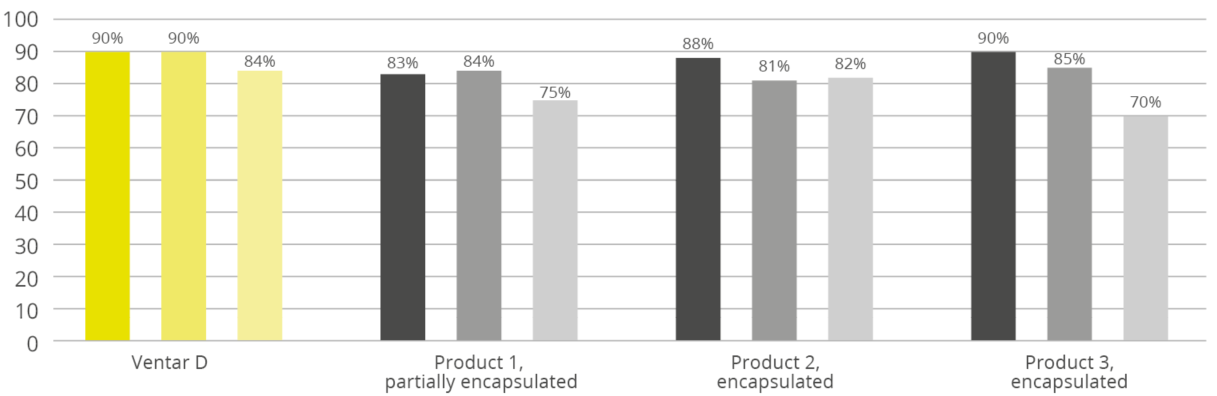
Phytogener Futterzusatz mit dem Schutz für Kontinuität im Ergebnis
Mit dieser Zielsetzung hat EW Nutrition eine Mikroverkapselung speziell für den Einsatz im Futter entwickelt. Erste Ergebnisse zeigen, dass die im Produkt Ventar D realisierte Technologie auch unter anspruchsvollen Pelletierungsbedingungen eine hohe Wiederfindungsraten der sensiblen Phytomoleküle gewährleistet. In einer Vergleichsstudie mit im Markt etablierten verkapselten Produkten konnte Ventar D in allen drei getesteten Szenarien (70 °C, 45 Sek; 80 °C, 90 Sek; 90 °C, 180 Sek) die höchsten Wiederfindungsraten erzielen. In dem Stresstest bei einer Temperatur von 90 °C über eine Dauer von 180 Sekunden konnten mindestens 84 % der wertvollen Phytomoleküle erhalten, während die Vergleichsprodukte zwischen 70 % und 82 % variierten. Unter einfacheren Bedingungen wurde eine konstante Wiederfindungsrate von 90 % erreicht.
Wiederfindungsraten für jedes Produkt, von links nach rechts: 70°C/45 Sekunden; 80°C/90 Sekunden; 90°C/180 Sekunden
Ökonomisch und ökologisch nachhaltig
In der Vergangenheit wurden die genannten Verluste in der Mischfutterproduktion und insbesondere beim Pelletieren als größtenteils unvermeidbar bezeichnet. Die phytogenen Zusatzstoffe mussten in den Mischfutterwerken, abhängig vom vorliegenden Prozess, häufig um 20 % bis 40 % überdosiert werden, um im fertigen Produkt die gewünschte Dosierung der wertvollen Phytomoleküle zu erhalten. Diese Vorgehensweise bedingte einen höheren Produkteinsatz und erhöhte somit die Kosten, sowie den damit verbundenen CO2-Fußabdruck. Die in Ventar D eingesetzte moderne Verkapselungstechnologie bietet nun einen deutlich besseren Schutz der wertvollen Phytomoleküle und bietet neben dem ökonomischen Vorteil auch einen effizienteren Umgang mit den zur Produktion notwendigen Ressourcen.
Referenzen
Hashemi, S. R .; Davoodi, H .; 2011; Herbal plants and their derivatives as growth and health promoters in animal nutrition; Vet Res Commun (2011) 35: 169-180; DOI 10.1007 / s11259-010-9458-2; Springer Science + Business Media BV, 2011
Ehrlinger, M., 2007: Phytogenic additives in animal nutrition. Inaugural dissertation. Munich: Veterinary Faculty of the Ludwig Maximilians University in Munich.
Lindner, U., 1987: Aromatic plants – cultivation and use. Contribution to the special show – Medicinal and Spice Plants (Federal Garden Show 1987), Teaching and Research Institute for Horticulture Auweiler-Friesdorf, Düsseldorf.
Was Sie schon immer über die nekrotisierende Enteritis beim Geflügel wissen wollten. Ein Überblick
by Inge Heinzl, Marisabel Caballero, Ajay Bhoyar, EW Nutrition
Die Eliminierung von nekrotisierender Enteritis (NE) aus Ihrem Bestand beginnt damit, die Krankheit zu verstehen, zu wissen, wie man sie verhindern kann und ob/wie es möglich ist, Auswirkungen von NE auf den Bestand zu mildern.
Nekrotisierende Enteritis ist eine Geflügelkrankheit, die durch ein übermäßiges Wachstum von Clostridium perfringens Typ A und in geringerem Maße Typ C im Dünndarm verursacht wird. Die Toxine dieser Bakterien schädigen die Darmwand. Im Allgemeinen tritt nekrotisierende Enteritis bei Masthähnchen im Alter von 2-6 Wochen auf. Subklinische Formen sind gekennzeichnet durch eine beeinträchtigte Futterverwertung. Klinische Formen führen zu schweren Problemen und einer erhöhten Bestandssterblichkeit in sehr kurzer Zeit.
Nekrotisierende Enteritis ist die Ursache für jährliche Verluste in Höhe von 6 Milliarden US-Dollar weltweit in der Geflügelproduktion – und diese vormals kontrollierbare Krankheit nimmt zu. Ein Grund dafür ist der freiwillige oder gesetzlich vorgeschriebene reduzierte Einsatz von Antibiotika in der Tierproduktion. Dieser Trend ergibt sich aus einem steigenden Auftreten antimikrobieller Resistenzen und aus den Anforderungen der Verbraucher. Ein weiterer Grund für das wieder verstärkte Auftreten von NE ist die Reduktion von Ionophoren, die neben ihrer Aktivität gegen Kokzidien auch Wirksamkeit gegen Clostridien zeigen. Wenn Lebendimpfstoffe gegen Kokzidien verwendet werden, ist die Anwendung dieser Ionophore nicht möglich und Clostridien / nekrotisierende Enteritis nehmen zu (Williams, 2005).
Nekrotisierende Enteritis und Kokzidiose sind weit verbreitet bei allen Geflügelarten, für Masthähnchen im Besonderen stellt diese Krankheit jedoch ein schwerwiegendes Gesundheitsproblem dar.
Die klinische und subklinische Form von NE
Die klinische Form
(c) Rob Moore
… ist gekennzeichnet durch akuten, dunklen Durchfall, der zu nasser Eintreu führt. Die Herdensterblichkeit steigt plötzlich um bis zu 1% pro Tag nach Auftreten der ersten klinischen Symptome (Ducatelle und Van Immerseel, 2010) und summiert sich teilweise auf bis zu 50% (Van der Sluis, 2013). Die Tiere haben zerzauste Federn, sind lethargisch und zeigen keinen Appetit.
Bei der Nekropsie fallen typischerweise aufgeblähte Gedärme mit einer aufgerauten Schleimhautoberfläche, Läsionen und bräunlichen (diphtherischen) Pseudomembranen auf. Größere Mengen an wässriger, brauner, blutdurchsetzter Flüssigkeit sind zu sehen und das Tier verströmt einen üblen Geruch. Die Leber ist dunkel, geschwollen und fest, die Gallenblase ist aufgetrieben (Hofacre et al., 2018).
Bei perakuter nekrotisierender Enteritis können die Tiere ohne vorherige Anzeichen sterben.
Die subklinische Form
Wenn die Tiere unter der subklinischen Form leiden, führen chronische Schäden an der Darmschleimhaut und eine erhöhte Schleimmenge im Dünndarm zu einer Beeinträchtigung der Verdauung. Eine deshalb geringere Absorption von Nährstoffen hat eine schlechtere FCR und damit eine geringere Wachstumsleistung zur Folge, was sich um Tag 35 bemerkbar macht.
Da Futtermittel etwa 65-75% der Produktionskosten bei Masthähnchen ausmachen, erhöht eine schlechte Futterverwertung die Produktionskosten und beeinflusst damit die Rentabilität erheblich. Aus Mangel an klaren Symptomen bleibt diese subklinische Form der Krankheit oft unbehandelt und wirkt sich dauerhaft auf die Produktionseffizienz aus.
Krankheitserreger
Verantwortlich für die nekrotisierende Enteritis sind grampositive, anaerobe Bakterien. Dabei handelt es sich um spezifische Stämme von Clostridium perfringens Typ A und, in geringerem Maße, Typ C (Keyburn et al., 2008).
Clostridien kommen in erster Linie im Boden vor, wo organische Substanzen abgebaut werden, außerdem im Abwasser und im Magen-Darm-Trakt von Tieren und Menschen. Sie produzieren Sporen, die extrem widerstandsfähig gegen Umwelteinflüsse (Wärme, Bestrahlung, Austrocknung) sowie auch einige Desinfektionsmittel sind und mehrere Jahre überleben können. Unter geeigneten Bedingungen können sich diese Bakterien über Sporen im Futter oder in der Einstreu sogar vermehren.
Clostridium perfringens Bakterien sind natürliche Bewohner des Hühnerdarms. In gesunden Tieren findet man normalerweise eine Mischung aus verschiedenen Stämmen in Konzentrationen von 102-104 KBE/g Darminhalt (McDevitt et al., 2006). Die Krankheit beginnt, wenn C. perfringens sich im Dünndarm vermehrt. In der Regel geschieht dies aufgrund einer Kombination verschiedener Faktoren wie z.B hohe Mengen an Protein im Futter, geringe Immunität und ein Ungleichgewicht in der Darmflora. Dann steigt die Zahl auf 107-109 KBE/g Darminhalt (Dahiya et al., 2005).
NetB, ein wichtiger Virulenzfaktor für NE
Um sich in einem Wirt anzusiedeln sind Clostridium Spp. und andere Krankheitserreger auf Virulenzfaktoren angewiesen (siehe Infobox). Zu diesen Virulenzfaktoren gehören beispielsweise die Voraussetzungen, um sich im Wirt festzusetzen, dem Immunsystem zu entgehen oder es zu unterdrücken, um an Nährstoffe zu gelangen und um sich in die Darmzellen einzuschleusen. Jahrelang wurde angenommen, dass das von C. perfringens produzierte α-Toxin an der Entwicklung der Krankheit beteiligt und ein wichtiger Virulenzfaktor ist. Im Jahr 2008 fanden Keyburn und Mitarbeiter jedoch einen weiteren wichtigen Virulenzfaktor. Sie verwendeten für ihren Versuch eine Mutante von C. perfringens. Diese Mutante ist nicht in der Lage, α-Toxin zu produzieren, verursacht aber dennoch nekrotisierende Enteritis.
Der Versuch identifizierte ein weiteres Toxin, das nur bei Hühnern zu finden ist, die an nekrotisierender Enteritis leiden: Das von Clostridium perfringens produzierte NetB (necrotic enteritis B-like toxin). NetB ist ein porenbildendes Toxin. Porenbildende Toxine sind Exotoxine, die in der Regel von pathogenen Bakterien produziert werden, es gibt jedoch auch andere Mikroorganismen, die dazu in der Lage sind. Diese Toxine zerstören die Membranintegrität von Darmwandzellen. Ausfließende Zellinhalte können von den Bakterien als Nährstoffe genutzt werden. Wenn Immunzellen auf diese Weise zerstört werden, könnte das eine Beeinträchtigung der Immunreaktion zur Folge haben (Los et al., 2013).
Darüber hinaus produzieren pathogene Stämme von C. perfringens Bakteriozine, um die Vermehrung harmloser Clostridium Spp.-Stämme zu hemmen und die normale Darmflora von Hühnern zu verdrängen (Riaz et al., 2017). Das wichtigste davon ist Perfrin (Timbermont et al., 2014).
Beispiele für Virulenzfaktoren
Adhäsine
Ermöglichen es dem Erreger, sich im Wirt festzusetzen, sich anzuheften, z. B. über Fimbrien. Pili ermöglichen den Austausch von RNA oder DNA zwischen Krankheitserregern.
Invasionsfaktoren
Erleichtern dem Erreger das Eindringen und die Verbreitung (Invasions- und Verbreitungsenzyme). Beispiele: Hyaluronidase, die die Hyaluronsäure des Bindegewebes angreift oder Geißeln zur Fortbewegung der Erreger.
Giftstoffe
Beeinträchtigen die Funktion der Wirtszellen oder zerstören sie; z.B. Endotoxine (Lipopolysaccharide), Exotoxine
Maßnahmen zur Umgehung der Immunabwehr Ermöglichen es dem Erreger, die Abwehrstrategien des Wirtes zu unterlaufen (z. B. Antiphagozytosefaktoren bieten Schutz vor einem Angriff durch Phagozyten; Inaktivierung spezifischer Antikörper durch Enzyme ).
Ein Huhn mit einer optimalen Darmgesundheit ist weniger anfällig für NE. Zusätzliche prädisponierende Faktoren sind notwendig: Nährstoffe müssen bereitgestellt und die Darmumgebung entsprechend vorbereitet werden, um eine Verbreitung dieser Krankheitserreger und damit den Ausbruch der Krankheit zu ermöglichen (Van Immerseel et al., 2008; Williams, 2005).
Prädisponierende Faktoren
Futter: Zusammensetzung und Partikelgröße
Die Rolle der Futtermittel bei der Entwicklung von nekrotisierender Enteritis sollte nicht unterschätzt werden. Hier sind Stoffe zu nennen, die eine für C. perfringens günstige Darmumgebung schaffen.
Mykotoxinkontamination
Mykotoxine schädigen die Darmintegrität und schaffen ideale Bedingungen für die Proliferation von Clostridium perfringens.
Mykotoxine haben keine direkte Wirkung auf die Verbreitung von C. Perfringens, die Toxinproduktion oder die NetB-Transkription. Mykotoxine beeinträchtigen jedoch die Darmgesundheit und schaffen eine günstige Umgebung für den Erreger. Beispiele:
Durch Angriff der Darmbarriere und die Beschädigung des Epithels schafft DON gute Bedingungen für die Proliferation von C. Perfringens. Die dadurch möglicherweise resultierende Durchlässigkeit des Epithels und eine verminderte Absorption von Proteinen aus dem Futter können zu einer höheren Restmenge an Proteinen im Dünndarm führen. Diese Proteine können als Nährstoffe für den Erreger dienen (Antonissen et al., 2014).
Fusariumtoxine verringern außerdem die Anzahl an Milchsäure produzierenden Bakterien, was zu einer Verschiebung des mikrobiellen Gleichgewichts führt (Antonissen et al., 2016.).
Eimeria ssp.
Ein intaktes Darmepithel ist die beste Abwehr gegen potenzielle Krankheitserreger wie C. perfringens. Hier kommen Kokzidien ins Spiel. Moore (2016) zeigte, dass Eimeria-Arten durch Schädigung des Darmepithels C. perfringens einen Zugang zu den Darmbasaldomänen des Schleimhautepithels liefern. Dort findet dann die erste Phase des pathologischen Prozesses statt. C. perfringens dringt in die Lamina propria ein und schädigt das Darmepithel (Olkowski et al., 2008). Die austretenden Plasmaproteine und der produzierte Schleim, sind reichhaltige Nährstoffquellen (Van Immerseel et al., 2004; Collier et al., 2008). Zusätzlich bewirkt die Kokzidiose eine Verschiebung des mikrobiellen Gleichgewichts im Darm. Die Anzahl von z.B. Candidatus savagella wird verringert, was eine Aktivierung der angeborenen Immunabwehr zur Folge hat.
.
A. Eimeria verursachen den Austritt von Plasmaproteinen durch Zerstörung von Epithelzellen
B. Sie erhöhen die Schleimproduktion im Darm
A+B führen zu einer Erhöhung der verfügbaren Nährstoffe und schaffen ein günstiges Umfeld für die Proliferation von C. perfringens
Nicht nur Eimeria spp., auch andere Krankheitserreger (z.B. Salmonella Spp., Ascaris- Larven, Viren) und die schon erwähnten Mykotoxine, die die Darmschleimhaut schädigen, können den Weg für eine C. perfringens-Infektion ebnen. Prädisponierende Faktoren wie nasse Einstreu, deren Feuchtigkeit für die Sporulation von Eimeria Spp. Oozysten wichtig ist, müssen auch als fördernd für nekrotisierende Enteritis betrachtet werden (Williams, 2005).
Immunsuppressive Faktoren
Neben den bereits beschriebenen Einflussfaktoren Futter, Mykotoxine und Kokzidien müssen noch andere erwähnt werden. Im Allgemeinen kann man sagen, dass alles, was Stress bei den Tieren auslöst, das Gleichgewicht der Darmflora stört. Die daraus resultierende Unterdrückung des Immunsystems erhöht das Risiko einer nekrotisierenden Enteritis (Tsiouris, 2016). Zu diesen Faktoren gehören:
Bakterien: Shivaramaiah und Mitarbeiter (2011) ermittelten eine Salmonella Typhimurium-Infektion bei frisch geschlüpften Küken als potentiellen prädisponierenden Faktor für NE. Die frühe Infektion verursacht erhebliche Schäden am Darm (Porter et al., 1998). Weiterhin zeigten Hassan et al. (1994), dass eine Infektion mit Salmonella typhimurium die Entwicklung der Lymphozyten negativ beeinflusst, was sich zusätzlich fördernd auf eine Kolonisierung mit Clostridium perfringens auswirken könnte.
Viren: Infektiöse Bursitis ist dafür bekannt, die Schwere von Infektionen mit Salmonellen, Staphylokokken, aber auch Clostridien zu erhöhen. Auch die Marek-Krankheit gilt als eine Clostridien fördernde Viruserkrankung.
Stress: Der Darmtrakt reagiert besonders empfindlich auf jede Art von Stress. Stress wird z.B. verursacht durch zu hohe Temperaturen, hohe Besatzdichten oder einen abrupten Futterwechsel.
Behandlung
In akuten Fällen sollte der Landwirt auf jeden Fall einen Tierarzt konsultieren und seine Tiere behandeln.
Da eine Behandlung in diesem Fall immer über Futter oder Wasser erfolgt, können dementsprechend nur Tiere behandelt werden, die noch Wasser oder Futter aufnehmen.
Antibiotika
Zur Behandlung von akuter NE werden normalerweise Antibiotika, die sich gegen grampositive Bakterien richteten, verwendet. Die Wahl des Antibiotikums sollte dabei dem Tierarzt überlassen werden. Er kennt den Wirkmechanismus und kann eventuelle Resistenzprobleme auf dem Betrieb / im Bestand berücksichtigen.
Ein prophylaktischer Einsatz von Antibiotika wird nicht empfohlen. In vielen Ländern ist dieser Einsatz bereits verboten, um Antimikrobielle Resistenz (AMR) zu reduzieren.
Antimikrobielle Resistenz (AMR)
Einige Bakterien reagieren aufgrund genetischer Mutationen weniger empfindlich auf bestimmte Antibiotika. Sie sind in der Lage:
die Produktion von Enzymen stimulieren, die Antibiotika abbauen oder modifizieren und damit inaktivieren (1).
die Eintrittspforten von Antibiotika zu beseitigen oder die Entwicklung von Pumpen zu fördern, die das Antibiotikum ausschleusen bevor es wirken kann (2).
Moleküle, an die das Antibiotikum bindet (die “Zielobjekte“) zu verändern oder zu beseitigen.
Das bedeutet, dass bei Verwendung eines bestimmten Antibiotikums Bakterien, die gegen dieses Antibiotikum resistent sind, überleben. Da sie jetzt keine Konkurrenten mehr haben, können sie sich bestens vermehren.
Leider kann diese Resistenz mit Hilfe von “Resistenzgenen” übertragen werden
an Tochterzellen
über deren Aufnahme von abgestorbenen Bakterien (3)
durch Gentransfer von Bakterium zu Bakterium (4)
durch Viren (5)
Jede Anwendung von Antibiotika fördert die Entwicklung von Resistenzen (Robert Koch Institut, 2019). Eine kurze Anwendung oder eine Anwendung mit niedriger Dosierung gibt den Bakterien die Möglichkeit, sich anzupassen.
Bakteriophagen
Experimente mit Phagen zeigten ein langsameres Fortschreiten der Krankheit und eine verminderte Ausprägung der Symptome von nekrotisierender Enteritis (Miller et al., 2010). Durch die orale Anwendung eines Bakteriophagencocktails konnten Miller et al. die Sterblichkeit bei C. perfringens im Vergleich zur unbehandelten Kontrolle um 92 % senken.
Wirkungsweise: Mit Hilfe von hochentwickelten Enzymen, den Endolysinen, sind Bakteriophagen in der Lage, die bakterielle Zellwand zu verdauen, und ihre Nachkommen in die Wirtszelle einzuschleusen (Fischetti, 2010). Eine Genehmigung für die Anwendung von Phagen seitens der EFSA steht noch aus.
Prävention
Eine Krankheit zu verhindern ist immer besser – und kostengünstiger – als sie zu behandeln.
Aber wie?
Am besten senkt man die Wahrscheinlichkeit für NE, indem man alle Faktoren, die die Vermehrung von Clostridium perfringens begünstigen und die Immunantwort der Tiere beeinträchtigen, möglichst eliminiert.
Neben der Elimination dieser prädisponierenden Faktoren liegt der Fokus auch auf:
Es gibt Hinweise darauf, dass die meisten Clostridium-Stämme, die von an nekrotisierender Enteritis leidenden Vögeln isoliert wurden, die Krankheit auch im Experiment auslösen können. Stämme, die von gesunden Vögeln isoliert wurden, sind dazu nicht in der Lage. Dies bestätigt, dass nur bestimmte Stämme problematisch sind (Ducatelle und Van Immerseel, 2010).
Daher ist es von höchster Bedeutung, die Einschleusung dieser pathogenen Stämme in den Betrieb auf jeden Fall zu vermeiden.
Separate Kleidung / Stiefel und in jedem Geflügelstall die Möglichkeit, Hände zu waschen und zu desinfizieren
Mindestens 14 Tage Leerstand bis zur Neubelegung der Ställe
Spezifische Maßnahmen gegen Kokzidiose
Impfung
Laut Parasitologen können 7 bis 9 Eimeria-Arten bei Hühnern nachgewiesen werden, eine Kreuzreaktivität besteht nicht. Eine wirksame Impfung muss dementsprechend sporulierte Oozysten der kritischsten pathogenen Eimeria-Arten enthalten (E. acervulina, E. maxima,E. tenella, E. necatrix und E. brunetti). Je mehr Arten im Impfstoff enthalten sind, desto besser. Durch eine inkorrekte Anwendung kann die Impfung jedoch unwirksam sein oder sogar Reaktionen bei den Tieren hervorrufen, die zu NE führen können (Mitchell, 2017).
Kokzidiostatika
Bei der Anwendung von Kokzidiostatika ist es wichtig, zwischen chemischen (synthetischen Verbindungen) und ionophoren (Polyether-Antibiotika) Kokzidiostatika mit unterschiedlichen Wirkungsweisen abzuwechseln, um die Entwicklung von Resistenzen zu vermeiden.
Ionophore haben eine spezielle Wirkungsweise: sie töten die Oozysten ab, und verhindern damit eine infektion des Geflügels. Da sie sehr klein sind, können die Moleküle der Ionophoren aufgenommen und in die äußere Membran des Sporozoits diffundiert werden. Dort senken sie den Konzentrationsgradienten. Die folgende Ansammlung von Wasser führt zum Bersten des Sporozoits.
Fütterung
Den Einsatz von Nicht-Stärke-Polysacchariden (NSPs) aus Getreide minimieren
Um eine “Fütterung” von Clostridium perfringens zu verhindern, sollte ein hoher Gehalt an wasserlöslichen, aber unverdaulichen NSPs und damit an Weizen, Weizennebenprodukten und Gerste vermieden oder zumindest minimiert werden. Darüber hinaus ist es von Vorteil Xylanasen in die Futterformulierung einzubeziehen, um die schädlichen Auswirkungen der NSPs zu reduzieren und die Nutzung der Energie aus dem Futter zu verbessern. Anstelle der oben genannten Getreidearten könnte Mais im Futter verwendet werden. Mais gilt als eine perfekte Komponente für Masthähnchen-Rationen aufgrund seines hohen Energiegehalts und seiner hohen Nährstoffverfügbarkeit.
Formulierung von eiweißarmen Rationen/Rationen mit hochverdaulichen Aminosäuren
Zur Minderung des Risikos einer nekrotisierenden Enteritis könnte die Fütterung von proteinarmen Rationen, die mit kristallinen Aminosäuren ergänzt werden, vorteilhaft sein (Dahiya et al., 2007). Der Zusatz von Proteasen verbessert die Proteinverdaulichkeit und reduziert damit die Proliferation von C. perfringens.
Tierische Fette und Fette minderer Qualität im Futter vermeiden bzw. minimieren
Durch die Fütterung von tierischen Fetten und Fetten minderer Qualität erhöht sich oft die Anzahl von Clostridium perfringens, daher sollten sie, wenn möglich, durch pflanzliche bzw. hochwertigere Fette ersetzt werden.
Futterbeschaffenheit
In Bezug auf die Futterbeschaffenheit stellten Engberg et al. (2002) fest, dass Tiere mit Pelletfütterung eine geringere Anzahl von Clostridium perfringens im Zäkum und Rektum aufwiesen als mit Maischefütterung. Branton und Mitarbeiter (1987) berichteten von einer niedrigeren Mortalitätsrate durch die Fütterung von Futtermitteln aus der Walzenmühle (grob gemahlen) als aus der Hammermühle.
Zusatzstoffe
Additive können entweder verwendet werden, um direkt eine Vermehrung von Clostridium perfringens zu verhindern oder um die Umweltbedingungen für eine Vermehrung von C. perfringens zu verschlechtern.
Probiotika
Diese lebenden Mikroben werden eingesetzt, um eine neue Darmflora zu etablieren, eine bestehende zu erhalten oder die Darmflora wiederherzustellen.
Wirkungsweise:
Probiotika konkurrieren mit pathogenen Bakterien um Substrate und Bindungsstellen
sie produzieren antimikrobielle Substanzen, die das Wachstum pathogener Bakterien hemmen (Gillor et al., 2008)
sie binden und neutralisieren Enterotoxine (Mathipa und Thantsha, 2017)
sie fördern die Immunfunktion des Wirts (Yang et al., 2012)
Präbiotika
Diese Futtermittelzusätze dienen als Substrate, um nützliche Bakterien im Darm zu fördern.
Wirkungsweise:
D-Mannose oder Fruktose, Stärkearten, die von Vögeln nicht verdaut werden können, stimulieren selektiv das Wachstum und die Aktivität der “guten” Darmflora
Fructooligosaccharide reduzieren C. perfringens und E. coli im Darm und erhöhen die Vielfalt von Lactobacillus Spp. (Kim et al., 2011)
Galactooligosaccharide, in Kombination mit einem B. lactis-basierten Probiotikum, sollen nach Berichten von Jung et al. (2008), selektiv die Proliferation von Bifidobacterium spp. fördern.
Organische Säuren
Organische Säuren werden oft in Futterrationen verwendet, um die Darmgesundheit zu verbessern.
Wirkungsweise:
niedriger pH-Wert fördert nützliche Bakterien
Caprylsäure unterdrückt C. perfringens, aber auch Salmonella Spp. indem sie deren Nutzung von Glucose hemmt (Skrivanova et al., 2006)
Lauric-, Zitronen-, Öl- und Linolsäure sowie mittelkettige Fettsäuren (C8-C14) behindern das Wachstum von C. perfringens
Viele Studien haben die Wirksamkeit von Tanninen gegen verschiedene Krankheitserreger wie Helminthen, Eimeria,, Viren und Bakterien gezeigt
Extrakte aus Kastanien- und Quebrachobäumen wirken nicht nur gegen C. perfringens, sondern auch gegen deren Toxine (Elizando et al., 2010)
Tannine zeigen Wirkung gegen Eimeria (Cejas et al., 2011) und Salmonella Sp., zwei prädisponierende Faktoren für NE.
Ätherische Öle
Da ätherische Öle hydrophob sind, können sie mit den Lipiden der Membran von C. perfringens interagieren.
Sie können sich in die bakterielle Membran einlagern und ihre Integrität stören.
Dies erhöht die Durchlässigkeit der Zellmembran für Ionen und andere kleine Moleküle wie ATP, was zur Abnahme des elektrochemischen Gradienten über der Zellmembran und zum Verlust von Energieäquivalenten führt.
Neben ihrer direkten Wirkung auf Clostridium Spp. verbessern viele Phytomoleküle die Darmgesundheit und helfen, eine Proliferation von Clostridium Spp. und damit nekrotisierende Enteritis zu verhindern.
Binder für Mykotoxine/bakterielle Toxine
Diese Binder wirken auf zweierlei Weise:
Durch Bindung von Mykotoxinen können Schäden am Darmepithel – eine Voraussetzung für die Vermehrung von Clostridien – reduziert oder sogar verhindert werden.
Durch Bindung der von Clostridium perfringens produzierten Toxine können Auftreten oder Schweregrad von Läsionen vermindert werden:
Alpha-Toxin (Phospholipase C) hydrolysiert Membranphospholipide, schädigt damit Erythrozyten, Leukozyten, Myozyten und Endothelzellen und verursacht ihre Zerstörung (Songer, 1996). Dies führt zu Nekrose und Gewebeschäden.
Auch die Bindung NetB Toxin, dem wichtigsten Virulenzfaktor, könnte die Schwere der nekrotisierenden Enteritis reduzieren.
Schlussfolgerung
Der ständig wachsende Trend, den Einsatz von Antibiotika und Ionophoren in der Geflügelproduktion zu reduzieren, trägt zu einem erhöhten Auftreten von nekrotisierender Enteritis bei.
Die subklinische Form der nekrotisierenden Enteritis bleibt in der Regel unbemerkt. Die resultierende schlechtere Futterverwertung ist eine der Hauptursachen der finanziellen Verluste in der Geflügelproduktion.
Um nekrotisierende Enteritis zu verhindern, ist die Aufrechterhaltung einer optimalen Darmgesundheit unumgänglich. Im Zeitalter einer antibiotikafreien Geflügelproduktion müssen Alternativen zur Bekämpfung dieses pathogenen Bakteriums und seiner prädisponierenden Faktoren in Betracht gezogen werden, um diese verheerende Krankheit wieder unter Kontrolle zu bekommen.
References
Annett, C.B., J. R. Viste, M. Chirino-Trejo, H. L. Classen, D. M. Middleton, and E. Simko. “Necrotic enteritis: effect of barley, wheat and corn diets on proliferation of Clostridium perfringens type A.” Avian Pathology 31 (2002): 599– 602. https://doi.org/10.1080/0307945021000024544
Antonissen G, F. Van Immerseel, F. Pasmans, R. Ducatelle, F. Haesebrouck, L. Timbermont, M. Verlinden, G.P.J. Janssens, V. Eeckhaut, M. Eeckhout, S. De Saeger, S. Hessenberger, A. Martel, and S. Croubels. “The mycotoxin deoxynivalenol predisposes for the development of Clostridium perfringens-Induced necrotic enteritis in broiler chickens. PLoS ONE 9 no. 9 (2014): e108775. https://doi.org/10.1371/journal.pone.0108775
Antonissen, G., V. Eeckhaut, K. Van Driessche, L. Onrust , F. Haesebrouck, R. Ducatelle, R.J. Moore, and F. Van Immerseel. “Microbial Shifts Associated With Necrotic enteritis.” Avian Pathol. 45 no. 3 (2016): 308-312. https://doi.org/10.1080/03079457.2016.1152625
Branton, S.L., F.N. Reece, and W.M. Hagler. “Influence of a wheat diet on mortality of broiler chickens associated with necrotic enteritis.” Poultry Sci. 66 (1987): 1326-1330. https://doi.org/10.3382/ps.0661326
Cejas, E., S. Pinto, F. Prosdócimo, M. Batalle, H. Barrios, G. Tellez, and M. De Franceschi. “Evaluation of quebracho red wood (Schinopsis lorentzii) polyphenols vegetable extract for the reduction of coccidiosis in broiler chicks.” International Journal of Poultry Science 10 no. 5 (2011): 344–349. https://doi.org/10.3923/ijps.2011.344.349
Collier, C.T., C.L. Hofacre, A.M. Payne, D.B. Anderson, P. Kaiser, R.I. Mackie, and H.R. Gaskins. “Coccidia-induced mucogenesis promotes the onset of necrotic enteritis by supporting Clostridium perfringens growth.” Veterinary Immunology and Immunopathology 122 (2008):104–115.
Dahiya, J.P., D. Hoehler, A.G. Van Kessel, and M.D. Drew. “Effect of different dietary methionine sources on intestinal microbial populations in broiler chickens.” Poultry Science 86 (2007):2358–2366
Dahiya, J.P., D. Hoehler, D.C. Wilkie, A.G. van Kessel, and M.D. Drew. “Dietary glycine concentration affects intestinal Clostridium perfringens and Lactobacilli populations in broiler chickens.” Poultry Science 84 no.12 (2005):1875-85. https://doi.org/10.1093/ps/84.12.1875
Diaz Carrasco, J.M., L.M. Redondo, E.A. Redondo, J.E. Dominguez, A.P. Chacana, and M.E. Fernandez Miyakawa. “Use of plant extracts as an effective manner to control Clostridium perfringens induced necrotic enteritis in poultry.” BioMed Research International (2016): Article ID 3278359. https://dx.doi.org/10.1155/2016/3278359
Ducatelle, R. and F. van Immerseel. “Necrotic enteritis: emerging problem in broilers.” WATTAgNet.com – Poultry Health and Disease (April 9, 2010).
Elizondo, A.M., E.C. Mercado, B.C. Rabinovitz, and M.E. Fernandez-Miyakawa. “Effect of tannins on the in vitro growth of Clostridium perfringens.” Veterinary Microbiology 145 no. 3-4 (2010): 308–314. https://doi.org/10.1016/j.vetmic.2010.04.003
Engberg, R.M., M.S. Hedemann, and B.B. Jensen. “The influence of grinding and pelleting of feed on the microbial composition and activity in the digestive tract of broiler chickens.” · British Poultry Science 43 no. 4 (2002):569-579. https://doi.org/10.1080/0007166022000004480
Fischetti, V.A. “Bacteriophage endolysins: A novel anti-infective to control Gram-positive pathogens.” J Med Microbiol. 300 no. 6 (2010): 357–362. https://doi.org/10.1016/j.ijmm.2010.04.002
Gillor, O., A. Etzion and M.A. Riley. “The dual role of bacteriocins as anti- and probiotics.” Appl Microbiol Biotechnol. 81 no. 4 (2008): 591–606. https://doi.org/10.1007/s00253-008-1726-5
Hassan, J. O., and R. Curtiss III. “Virulent Salmonella typhimurium induced lymphocyte depletion and immunosuppression in chickens.” Infect. Immun. 62 (1994):2027–2036 https://doi.org/10.1128/IAI.62.5.2027-2036.1994
Hofacre, C.L., J.A. Smith, and G.F. Mathis. “Invited Review. An optimist’s view on limiting necrotic enteritis and maintaining broiler gut health and performance in today’s marketing, food safety, and regulatory climate.” Poultry Science 97 (2018):1929–1933. https://dx.doi.org/10.3382/ps/pey082
Jung, S.J., R. Houde, B. Baurhoo, X. Zhao, and B. H. Lee. “Effects of galacto-oligosaccharides and a bifidobacteria lactis-based probiotic strain on the growth performance and fecal microflora of broiler chickens.” Poultry Science 87 (2008):1694–1699. https://doi.org/10.3382/ps.2007-00489
Kaldhusdal and Skjerve. “Association between cereal contents in the diet and incidence of necrotic enteritis in broiler chickens in Norway.” Preventive Veterinary Medicine 28 (1996):1-16. https://doi.org/10.1016/0167-5877(96)01021-5
Keyburn, A. L., S. A. Sheedy, M. E. Ford, M. M. Williamson, M. M. Awad, J. I. Rood, and R. J. Moore. “Alpha-toxin of Clostridium perfringens is not an essential virulence factor in necrotic enteritis in chickens.” Infect. Immun. 74 (2006): 6496–6500. https://doi.org/10.1128/IAI.00806-06
Keyburn, A.L., J.D. Boyce, P. Vaz, T.L. Bannam, M.E. Ford, D. Parker, A. Di Rubbo, J.I. Rood, and R.J. Moore. “NetB, a new toxin that is associated with avian necrotic enteritis caused by Clostridium perfringens.” PLoS Pathog 4 no. 2, e26 (2008): 0001-0011. https://doi.org/10.1371/journal.ppat.0040026
Kim, G.-B., Y. M. Seo , C. H. Kim , and I. K. Paik. “Effect of dietary prebiotic supplementation on the performance, intestinal microflora, and immune response of broilers.” Poultry Science 90 (2011):75–82. https://doi.org/10.3382/ps.2010-00732
Knap, I., B. Lund, A. B. Kehlet, C. Hofacre, and G. Mathis. “Bacillus licheniformis prevents necrotic enteritis in broiler chickens.” Avian Diseases 54 no. 2 (2010):931-935. https://doi.org/10.1637/9106-101509-ResNote.1
Knarreborg, A., M.A. Simon, R.M. Engberg, B.B. Jensen, and G.W. Tannock. “Effects of Dietary Fat Source and Subtherapeutic Levels of Antibioticon the Bacterial Community in the Ileum of Broiler Chickensat Various Ages.” Applied and Environmental Microbiology 68 no. 12 (2002): 5918-5924. https://doi.org/0.1128/AEM.68.12.5918–5924.2002
Kocher, A. and M. Choct. “Improving broiler chicken performance. The efficacy of organic acids, prebiotics and enzymes in controlling necrotic enteritis.” Australian Government-Rural Industries Research and Development Corporation. Publ. no. 08/149 (2008).
Kondo, F. “In vitro lecithinase activity and sensitivity to 22 antimicrobial agents of Clostridium perfringens isolated from necrotic enteritis of broiler chickens.” Research in veterinary Science 45 (1988): 337-340. https://doi.org/10.1016/S0034-5288(18)30961-5
Kubena, L.F., J.A. Byrd, C.R. Young, and D.E. Corrier. “Effects of tannic acid on cecal volatile fatty acids and susceptibility to Salmonella typhimurium colonization in broiler chicks.” Poultry Science 80, no. 9 (2001): 1293–1298. https://doi.org/10.1093/ps/80.9.1293
Los, F.C.O., T.M. Randis, R.V. Aroian, and A.J. Ratner. “Role of pore-forming toxins in bacterial infectious diseases.” Microbiology and Molecular Biology Reviews 77 (2013): 173-207 https://doi.org/10.1128/MMBR.00052-12
M’Sadeq S.A., Shubiao Wu, Robert A. Swick, Mingan Choct. “Towards the control of necrotic enteritis in broiler chickens with in-feed antibiotics phasing-out worldwide.” Animal Nutrition 1 (2015): 1-11. https://dx.doi.org/10.1016/j.aninu.2015.02.004
Mathipa, M.G. and M.S. Thantsha. “Probiotic engineering: towards development of robust probiotic strains with enhanced functional properties and for targeted control of enteric pathogens.” Gut Pathog. 9 no. 28 (2017). https://doi.org/10.1186/s13099-017-0178-9
McDevitt, R.M., J.D. Brooker, T. Acamovic, and N.H.C. Sparks. “Necrotic enteritis, a continuing challenge for the poultry industry.” World’s Poultry Science Journal 62; World’s Poultry Science Association (June 2006). https://doi.org/10.1079/WPS200593
Miller, R.W., J. Skinner, A. Sulakvelidze, G.F. Mathis, and C.L. Hofacre. “Bacteriophage therapy for control of necrotic enteritis of broiler chickens experimentally infected with Clostridium perfringens.” Avian Diseases 54 no. 1 (2010): 33-40. https://doi.org/10.1637/8953-060509-Reg.1
Mitsch, P., K. Zitterl-Eglseer, B. Köhler, C. Gabler, R. Losa, and I. Zimpernik. “The Effect of Two Different Blends of Essential Oil Components on the Proliferation of Clostridium perfringens in the Intestines of Broiler Chickens.” Poultry Science 83 (2004):669–675. https://doi.org/10.1093/ps/83.4.669
Olkowski, A.A., C. Wojnarowicz, M. Chirino-Trejo, B. Laarveld, and G. Sawicki. “Sub-clinical necrotic enteritis in broiler chickens: Novel etiological consideration based on ultra-structural and molecular changes in the intestinal tissue.” Veterinary Science 85 (2008): 543–553. https://doi.org/10.1016/j.rvsc.2008.02.007
Pan, D. and Z. Yu. “Intestinal microbiome of poultry and its interaction with host and diet.” Gut Microbes 5 no. 1 (2014): 108–119. https://dx.doi.org/10.4161/gmic.26945
Rougière, N. and B. Carré. “Comparison of gastrointestinal transit times between chickens from D + and D- genetic lines selected for divergent digestion efficiency.” Animal 4 no. 11 (2010): 1861-1872. https://doi.org/10.1017/S1751731110001266
Santos, F.B.O., B.W. Sheldon, A.A. Santos Jr., and P.R. Ferket. ”Influence of housing system, grain type, and particle size on Salmonella colonization and shedding of broilers fed triticale or corn-soybean meal diets.” Poultry Science 87 (2008): 405-420. https://dx.doi.org/10.3382/ps.2006-00417
Schiavone, A. , K. Guo, S. Tassone, L .Gasco, E. Hernandez, R. Denti, and I. Zoccarato. “Effects of a Natural Extract of Chestnut Wood on Digestibility, Performance Traits, and Nitrogen Balance of Broiler Chicks.” Poult Sci. 87 no. 3 (2008): 521-527. https://doi.org/10.3382/ps.2007-00113
Shivaramaiah, S., R. E. Wolfenden, J. R. Barta, M. J. Morgan, A. D. Wolfenden, B. M. Hargis, and G. Téllez. „The role of an early Salmonella typhimurium infection as a predisposing factor for necrotic enteritis in a laboratory challenge model.” Avian Diseases 55 (2011): 319-323. https://doi.org/10.1637/9604-112910-ResNote.1
Singh, Y., V. Ravindran, T.J. Wester, A.L. Molan, and G. Ravindran. “ Influence of feeding coarse corn on performance, nutrient utilization, digestive tract measurements, carcass characteristics, and cecal microflora counts of broilers.” Poultry Science 93 (2014): 607–616. https://dx.doi.org/10.3382/ps.2013-03542
Skrivanova, E., M. Marounek, V. Benda, and P. Brezina. “Susceptibility of Escherichia coli, Salmonella sp. and Clostridium perfringens to organic acids and monolaurin.” Veterinarni Medicina 51 no. 3 (2006): 81–88. https://doi.org/10.17221/5524-VETMED
Stanley D., Wu S.-B., Rodgers N., Swick R.A., and Moore R.J. “Differential Responses of Cecal Microbiota to Fishmeal, Eimeria and Clostridium perfringens in a Necrotic Enteritis Challenge Model in Chickens.” PLoS ONE 9 no. 8 (2014): e104739. https://doi.org/10.1371/journal.pone.0104739
Tan, L., D. Rong, Y. Yang, and B. Zhang. “Effect of Oxidized Soybean Oils on Oxidative Status and Intestinal Barrier Function in Broiler Chickens.” Brazilian Journal of Poultry Science 18 no. 2 (2018): 333-342. http://dx.doi.org/10.1590/1806-9061-2017-0610
Tan, L., D. Rong, Y. Yang, and B. Zhang. “The Effect of Oxidized Fish Oils on Growth Performance, Oxidative Status, and Intestinal Barrier Function in Broiler Chickens.” J. Appl. Poult. Res. 28 (2019): 31-41. http://dx.doi.org/10.3382/japr/pfy013
Timbermont L., A. Lanckriet, J. Dewulf, N. Nollet, K. Schwarzer, F. Haesebrouck, R. Ducatelle, and F. Van Immerseel. “Control of Clostridium perfringens-induced necrotic enteritis in broilers by target-released butyric acid, fatty acids and essential oils.” Avian Pathol. 39 no. 2 (2010): 117-21. https://doi.org/10.1080/03079451003610586
Tsiouris, V. “Poultry management: a useful tool for the control of necrotic enteritis in poultry.” Avian Pathol. 45 no. 3 (2016):323-325. https://doi.org/10.1080/03079457.2016.1154502
Van der Most, P.J., B. de Jong, H.K. Parmentier and S. Verhulst. “Trade-off between growth and immune function: a meta-analysis of selection experiments.” Functional Ecology 25 (2011): 74-80. https://doi.org/0.1111/j.1365-2435.2010.01800.x
Van der Sluis, W. “Clostridial enteritis is an often underestimated problem.” Worlds Poult. Sci. J. 16 (2000):42–43.
Van Immerseel, F., J. De Buck, F. Pasmans, G. Huyghebaert, F. Haesebrouck, and R. Ducatelle. “Clostridium perfringens in poultry: an emerging threat of animal and public health.” Avian Pathology 33 (2004): 537-549. https://doi.org/10.1080/03079450400013162
Van Immerseel, F., J.I. Rood, R.J. Moore, and R.W. Titball. “Rethinking our understanding of the pathogenesis of necrotic enteritis in chickens.” Trends in Microbiology 17 no. 1 (2008):32-36. https://doi.org/10.1016/j.tim.2008.09.005
Wade, B., A.L. Keyburn, T. Seemann, J.I. Rood, and R.J. Moore. “Binding of Clostridium perfringens to collagen correlates with the ability to cause necrotic enteritis in chickens.” Veterinary Microbiology 180 no. 3–4 (2015): 299-303. https://doi.org/10.1016/j.vetmic.2015.09.019
Williams, R.B. “Intercurrent coccidiosis and necrotic enteritis of chickens: rational, integrated disease management by maintenance of gut integrity.” Avian Pathology 34 no. 3 (2005):159-180. https://doi.org/10.1080/03079450500112195
Yang , C.M., G.T. Cao, P.R. Ferket, T.T. Liu, L. Zhou, L. Zhang, Y.P. Xiao, and A. G. Chen. “ Effects of probiotic, Clostridium butyricum, on growth performance, immune function, and cecal microflora in broiler chickens.” Poultry Science 91 (2012): 2121–2129. https://dx.doi.org/10.3382/ps.2011-02131
Stehen Endotoxine hinter Ihrer geringen Tierproduktivität?
Machen Stresssituationen Ihre Tiere krank oder verschlimmert sich eine Krankheit nach Antibiotikabehandlung? Endotoxine könnten die Ursache sein!
Was sind Endotoxine?
Herkunft
Endotoxine wie auch Exotoxine sind bakterielle Toxine. Im Gegensatz zu Exotoxinen, die von lebenden Bakterien aktiv ausgeschieden werden, sind Endotoxine („Endotoxin“ kommt aus dem Griechischen; Endo = Innen; Toxin = Gift) Bestandteile der äußeren Zellmembran von gramnegativen Bakterien wie Escherichia coli, Salmonella, Shigella, und Cyanobakterien (Blaualgen). Sie werden nur freigegeben im Falle von
bakteriellem Tod aufgrund eines wirksamen Abwehrmechanismus des Wirts oder der Wirkung bestimmter Antibiotika
Biochemisch gehören Endotoxine zu den Lipopolysacchariden (LPS). Sie bestehen aus einer relativ einheitlichen Lipidfraktion (Lipid A) und einer speziesspezifischen Polysaccharidkette. Ihre Toxizität ist hauptsächlich auf das Lipid A zurückzuführen. Der Polysaccharidteil ist verantwortlich für ihre Aktivität. Im Gegensatz zu den Bakterien sind ihre Endotoxine sehr hitzebeständig und halten einer Sterilisation stand. Die Namen Endotoxin und Lipopolysaccharide werden synonym verwendet, wobei bei “Endotoxin” das Augenmerk auf dem Vorkommen und der biologischen Aktivität liegt, bei “Lipopolysaccharid” mehr die chemische Struktur hervorgehoben wird (Hurley, 1995).
Auswirkungen
Endotoxine gehören zu den fiebererzeugenden Substanzen. Sie aktivieren die Signalwege mehrerer immunkompetenter Zellen. Ein früher Kontakt mit Endotoxinen führt zur Aktivierung und Reifung des erworbenen Immunsystems. Braun-Fahrländer und Mitarbeiter (2002) stellten fest, dass Kinder, die mit Endotoxinen in Berührung kamen, weniger Probleme mit Heuschnupfen, atopischem Asthma und atopischer Sensibilisierung hatten. Dies passt zusammen mit der Tatsache, dass in menschlichen Populationen nach Erhöhung der Hygienestandards eine Zunahme von Allergien beobachtet werden konnte.
Verschiedene Tierarten zeigen unterschiedliche Empfindlichkeiten gegenüber Endotoxininfusionen, z. B. (gesunde) Hunde, Ratten, Mäuse, Hühner tolerieren Konzentrationen ≥ 1 mg / kg Körpergewicht, während (gesunde) Wiederkäuer, Schweine und Pferde bereits bei Konzentrationen <5 μg/kg Körpergewicht sehr empfindlich reagieren (Olson et al., 1995, zitiert in Wilken, 2003).
Wann ist ein Organismus einer höheren Belastung mit Endotoxinen ausgesetzt?
Endotoxine kommen natürlicherweise im Darm vor, da die Mikroflora auch gramnegative Bakterien enthält. Sie richten erst Schaden an, wenn sie in den Blutkreislauf gelangen. Solange der Endotoxinspiegel niedrig ist, kann das Immunsystem damit zurechtkommen, höhere Konzentrationen können jedoch kritisch werden. Ein Anstieg der Endotoxine im Organismus resultiert aus einem höheren Input und / oder einer geringeren Clearance- bzw. Entgiftungsrate.
Wie kommen mehr Endotoxine in den Organismus
„Normale“, geringe Mengen an Endotoxinen, die aufgrund regelmäßiger bakterieller Aktivität im Darm entstehen und in den Organismus übertragen werden, haben keine negativen Auswirkungen, solange die Leber ihre Clearance-Funktion erfüllt. Auch die im Fettgewebe gespeicherten Endotoxine sind unproblematisch. Einige Faktoren können jedoch zu einer Freisetzung der Endotoxine oder deren Translokation in den Organismus führen:
Stress
Stresssituationen wie Geburt, Operationen, Verletzungen können zu einer Ischämie (mangenlde Blutversorgung) im Darm und zur Übertragung von Endotoxinen in den Organismus führen (Krüger, 1997). Andere Stresssituationen in der Tierproduktion, wie hohe Temperaturen und hohe Besatzdichten, tragen auch zu höheren Endotoxinspiegeln im Blutkreislauf bei: Stress führt zu einem höheren Stoffwechselbedarf an Wasser, Natrium und energiereichen Substanzen. Für eine höhere Verfügbarkeit dieser Substanzen wird die Permeabilität der Darmbarriere erhöht, was möglicherweise auch zu einer höheren Translokation von Bakterien und ihren Toxinen in den Blutkreislauf führt.
Beispiele:
In einer experimentellen Studie von Seidler (1998) (zitiert in Wilken (2003)) wurden höhere Endotoxinwerte bei Schweinen festgestellt, die unter Stress aufgrund von Belastung, Transport und damit verbundenen erhöhten Temperaturen litten.
Marathonläufer (Brock-Utne et al., 1988) und Rennpferde (Baker et al., 1988) zeigten ebenfalls höhere Endotoxinkonzentrationen im Blut proportional zum Laufstress. Dabei liegen bei trainierten Pferden die Konzentrationen niedriger als bei nicht trainierten.
Abbau von Körperfett zur Energiemobilisierung
Wenn Endotoxine aufgrund von anhaltendem Stress ständig in den Blutkreislauf gelangen, können sie im Fettgewebe gespeichert werden. Der SR-B1 (Scavenger-Rezeptor B1, ein Membranrezeptor, der zur Gruppe der Mustererkennungsrezeptoren (pattern-recognition receptor – PRR) gehört) bindet sowohl an Lipide als auch an Lipopolysaccharide und fördert damit wahrscheinlich den Einbau von LPS in Chylomikronen. Von Chylomikronen auf andere Lipoproteine übertragen, gelangt LPS schließlich ins Fettgewebe (Hersoug et al., 2016). Die Mobilisierung von Energie durch Fettabbau, z. B. zu Beginn der Laktation, führt beispielsweise zu einer Wiedereinspeisung von Endotoxinen in den Blutkreislauf.
Beschädigung der Darmbarriere
Wie bereits beschrieben, sind Endotoxine aufgrund bakterieller Aktivität unter normalen Bedingungen im Darm vorhanden. Eine Beschädigung der Darmbarriere ermöglicht dementsprechend den Übergang von Endotoxinen (und Bakterien) in den Blutkreislauf
Zerstörung von gramnegativen Bakterien
Eine weitere „Quelle“ für Endotoxine ist die Zerstörung der Bakterien. Dies kann zum einen durch das Immunsystem des Organismus oder durch die Behandlung mit bakteriziden Substanzen gegen gramnegative Bakterien erfolgen (Kastner, 2002). Um eine erhöhte Freisetzung von Endotoxinen zu verhindern, sollten dementsprechend gramnegative Bakterien mit bakteriostatischen Substanzen, die nur das Wachstum hemmen und die Bakterien nicht zerstören, oder mit bakteriziden Substanzen in Kombination mit LPS-Bindemitteln behandelt werden (Brandenburg, 2014).
Vermehrung von gramnegativen Bakterien
Da gramnegative Bakterien auch beim Wachstum geringe Mengen an Endotoxinen freisetzen, führt alles, was ihre Proliferation fördert, auch zu einem Anstieg der Endotoxine:
Ungleichgewicht in der Fütterung
Hochleistungskühe z. B. erhalten für die Milchproduktion Futter mit hohen Gehalten an Stärke, Fett und Eiweiß. Hohe Gehalte an Kohlenhydraten führen zu einem Anstieg von gramnegativen Bakterien. Gleichzeitig führt aber diese Fütterung auch zu einer Absenkung des pH (Azidose), was dann zur Abtötung von gramnegativen Bakterien und damit zur Freisetzung von Endotoxinen führen kann. Futter mit hohen Fettgehalten führt zu einer höheren Konzentration von Endotoxinen im Organismus, da derselbe „Transporter“ (Scavenger-Rezeptor Klasse B Typ 1, SR-BI) für die Absorption von Fett (Hersoug et al., 2016) aber von Endotoxinen verwendet werden kann.
In einer Studie von Deopurkar et al. (2010) mit Menschen als Vertretern der Monogastrier wurden gesunden Teilnehmern drei verschiedene Getränke (Glukose – 100% Kohlenhydrate, Orangensaft – 92% Kohlenhydrate und Sahne – 100% Fett) zum Trinken gegeben. Nur das Sahnegetränk erhöhte den Gehalt an Lipopolysacchariden im Plasma.
Infektionskrankheiten
Infektionskrankheiten wie Mastitis, Metritis und andere Infektionen, die durch gramnegative Bakterien wie E. coli, Salmonellen usw. verursacht werden, können als Ursache für die Freisetzung von Endotoxinen angesehen werden
Eingeschränkte Entgiftung, verminderter Abbau
Hauptverantwortliches Organ: die Leber
Aufgabe: Entgiftung und Abbau von in den Blutkreislauf gelangtem Endotoxin. Die Leber produziert Substanzen wie Lipopolysaccharid-Bindungsproteine (LBP), die zur Bindung und Neutralisierung von Lipopolysaccharid-Strukturen notwendig sind.
Während der postpartalen Periode befindet sich der Organismus in einer katabolen Phase, mit einem erhöhten Fettabbau versucht der Organismus, den hohen Energiebedarf für die Milchproduktion zu decken. Eine gesteigerte Lipolyse führt, wie bereits erwähnt, zu einer Freisetzung von Endotoxinen aus dem Fettgewebe, aber auch zu einer Verfettung der Leber. Eine verfettete und damit degenerierte Leber kann bei der Endotoxin-Clearance nicht die gleiche Leistung erbringen wie eine normale (Andersen, 2003; Andersen et al., 1996; Harte et al., 2010; Wilken, 2003). In einer Studie von Andersen et al. (1996) konnte bei Kühen mit Fettleber keine vollständige Elimination der Endotoxine erreicht werden. Das Auftreten einer Fettleber nimmt nach der Geburt zu (Reid und Roberts, 1993; Wilken, 2003).
Auch andere Erkrankungen der Leber beeinflussen die Endotoxin-Clearance in der Leber. Hanslin et al. (2019) fanden eine beeinträchtigte Endotoxin-Elimination bei Schweinen mit schon bestehendem, systemischem inflammatorischem Response-Syndrom (SIRS).
Probleme durch Endotoxine
Endotoxine in geringer Konzentration können das Immunsystem positiv stimulieren (Sampath, 2018). Nach McAleer und Vella (2008) werden Lipopolysaccharide als natürliche Adjuvantien verwendet, um bei einer Impfung die Immunreaktion durch Beeinflussung der CD4+T-Zellantwort zu stärken. Problematisch an Endotoxinen ist ihre Beteiligung an der Entwicklung schwerwiegender Probleme wie des MMA-Komplexes (Pig Progress) oder eines septischen Schocks (Sampath, 2018).
MMA-Komplex bei Sauen
MMA bei Sauen ist eine multifaktorielle Erkrankung, die kurz (12 Stunden bis drei Tage) nach dem Abferkeln auftritt und durch verschiedene Faktoren (Krankheitserreger wie E. coli, Klebsiella spp., Staph. Spp. und Mycoplasma spp., aber auch Stress, Ernährung) verursacht werden kann. MMA ist auch als puerperales Syndrom, puerperale Septikämie, Milchfieber oder Toxämie bekannt. Die letzte Bezeichnung legt nahe, dass einer der Faktoren, die die Krankheit mit verursachen, bakterielle Endotoxine sind. Während der perinatalen Phase findet zur Unterstützung der Laktation ein massiver Fettabbau statt. Die Sauen leiden häufig unter Verstopfung. Dies kann zu einer höheren Durchlässigkeit der Darmwand und zu einer Übertragung von Bakterien bzw. Endotoxinen in den Blutkreislauf führen. Eine weitere „Quelle“ für Endotoxine kann das Euter sein, da gramnegative Bakterien in erhöhtem Maße in den Brustdrüsen vorkommen (Morkoc et al., 1983).
Die Endotoxine können zu einer endokrinen Dysfunktion führen: ↑ Cortisol, ↓ PGF2α, ↓ Prolactin, ↓ Oxytocin. MMA steht für:
Mastitis, eine bakterielle Infektion des Euters
Mastitis kann von zwei Seiten provoziert werden: Einerseits führt Endotoxämie zu einer Erhöhung der Zytokine (IL1, 6, TNFα). Niedrigere Ca- und K-Spiegel führen dazu, dass der Zitzenschließmuskel weniger funktionsfähig ist, was den Eintritt von Umweltpathogenen ins Euter erleichtert und zu Mastitis führt. Andererseits steigt die Cortisolkonzentration als Reaktion auf den Abferkelstress. Die resultierende Immunsuppression ermöglicht es E. coli, sich im Euter zu vermehren.
Metritis, eine Infektion der Gebärmutter mit Ausfluss im Vulvabereich
Die Gebärmutterentzündung führt zu reduzierten Kontraktionen und damit zu einem sich länger hinziehenden und/oder komplizierten Abferkeln oder zu toten Ferkeln. Stress und die damit verbundene Abnahme der Oxytocin- und Prostaglandin-F2α-Sekretion kann eine Infektion der Gebärmutter fördern. Morkoc et al. (1983) fanden keinen Zusammenhang zwischen Metritis und Endotoxinen.
Agalaktie, eine verringerte oder komplett fehlende Milchproduktion
In vielen Fällen wird eine Agalaktie erst erkannt, wenn die Saugferkel Anzeichen von Hunger und / oder Gewichtsverlust zeigen. Im schlimmsten Fall steigt die Sterblichkeitsrate bei den Ferkeln. Wahrscheinlich wird ein Milchmangel durch zu niedrigere Konzentrationen der an der Laktation beteiligten Hormone verursacht. Der Prolaktinspiegel kann z. B. schon durch geringe Mengen an Endotoxin dramatisch gesenkt werden (Smith und Wagner, 1984). Der Oxytocinspiegel ist oft nur halb so hoch wie bei normalen Sauen (Pig Progress, 2020).
Endotoxinschock
Ein septischer Schock kann die Reaktion auf die Freisetzung einer hohen Menge an Endotoxinen sein.
Im Falle einer Infektion mit gramnegativen Bakterien werden die Tiere mit (oft bakteriziden) Antibiotika behandelt. Außerdem findet eine Eliminierung der Bakterien durch das Immunsystem statt. Durch das Absterben vieler Bakterien werden Endotoxine massiv freigesetzt. Im ungebundenen Zustand aktivieren sie das Immunsystem, einschließlich Makrophagen, Monozyten und Endothelzellen. Das führt zur Freisetzung großer Mengen an zellulären Mediatoren wie TNFα, Interleukin 1 (IL-1), IL-6 und Leukotrienen. Hohe Mengen an proinflammatorischen Zytokinen aktivieren die Komplement- und Gerinnungskaskade. Bei einigen Tieren wird dann die Produktion von Prostaglandinen und Leukotrienen stimuliert. Hohes Fieber, verringerter Blutdruck, Bildung von Thromben im Blut, Kollaps, Schädigung mehrerer Organe und tödlicher (endotoxischer) Schock sind die Folge.
Ein endotoxischer Schock tritt nur bei wenigen anfälligen Tieren auf, obwohl die gesamte Herde möglicherweise immunstimuliert wurde. Ein weit größeres Problem stellt der Leistungsabfall der übrigen Ferkel dar: Aufgrund der Endotoxämie wird Energie anstatt für Wachstum für die Immunabwehr eingesetzt.
Verstärkter Durchfall
Lipopolysaccharide führen zu einer erhöhten Freisetzung von Prostaglandinen, die die Magen-Darm-Funktionen beeinflussen. Zusammen mit Leukotrienen und entzündungsfördernden Mediatoren in der Schleimhaut verringern sie die Darmabsorption (Munck et al., 1988; Chiossone et al., 1990), lösen aber auch einen pro-sekretorischen Zustand im Darm aus. Liang et al. (2005) beobachteten eine dosisabhängige Ansammlung von reichlich Flüssigkeit im Dünndarm, die bei Ratten zu einer erhöhtem Durchfall und einer verminderten Darmperistaltik führte.
Fazit
Maßnahmen gegen gramnegative Bakterien können zu einem noch schwerwiegenderen Problem führen – der Endotoxämie. Endotoxine wirken sich nicht nur direkt negativ auf den Organismus aus, sie fördern auch die Entwicklung einiger Krankheiten. Die Unterstützung der Darmgesundheit durch verschiedene Ansätze, einschließlich der Bindung von Toxinen, trägt dazu bei, Tiere gesund zu erhalten.
By Inge Heinzl, EW Nutrition
References
Andersen, P.H. “Bovine endotoxicosis – some aspects of relevance to production diseases. A review.” Acta vet. scand. Suppl. 98 (2003): 141-155. DOI: 10.1186/1751-0147-44-S1-P57
Andersen, P.H., N. Jarløv, M. Hesselholt, and L. Bæk. “Studies on in vivo Endotoxin Plasma Disappearance Times in Cattle.” Zentralblatt für Veterinärmedizin. Reihe A 43 no. 2(1996): 93-101. DOI: 10.1111/j.1439-0442.1996.tb00432.x
Baker, B., S.L. Gaffin, M. Wells, B.C. Wessels and J.G. Brock-Utne. “Endotoxaemia in racehorses following exertion.” Journal of the South African Veterinary Association June (1988): 63-66. https://journals.co.za/docserver/fulltext/savet/59/2/1341.pdf?expires=1598542211&id=id&accname=guest&checksum=E50C766D318776E09CA41DA912F14CAD
Beutler, B. and T. Rietschel. “Innate immune sensing and its roots: The story of endotoxin.” Nature Reviews / Immunology 3(2003): 169-176. DOI: 10.1038/nri1004
Braun-Fahrländer, C., J. Riedler, U. Herz, W. Eder, M. Waser, L. Grize, S. Maisch, D. Carr, F. Gerlach, A. Bufe, R.P. Lauener, R. Schierl, H. Renz, D. Nowak and E. von Mutius. „Environmental exposure to endotoxin and its relation to asthma in school-age children. ”The New England Journal of Medicine 347 (2002): 869-877. DOI: 10.1056/NEJMoa020057.
Chiossone, D. C., P.L. Simon, P.L. Smith. “Interleukin-1: effects on rabbit ileal mucosal ion transport in vitro.” European Journal of Pharmacology 180 no. 2-3 (1990): 217–228. DOI: 10.1016/0014-2999(90)90305-P.
Deopurkar R., H. Ghanim, J. Friedman, et al. “Differential effects of cream, glucose, and orange juice on inflammation, endotoxin, and the expression of Toll-like receptor-4 and suppressor of cytokine signaling-3.” Diabetes care 33 no. 5 (2010):991–997.
Erridge, C., E. Bennett-Guerrero, and I.R. Poxton. “Structure and function of lipopolysaccharides.” Microbes and Infection 4 no. 8 (2002): 837-851. DOI: 10.1016/s1286-4579(02)01604-0
Fritsche, D. “Endotoxinpromovierte bakterielle Translokationen und Besiedelung von Uterus und Euter beim Hochleistungsrind im peripartalen Zeitraum.“ Dissertation. Leipzig, Univ., Veterinärmed. Fak. (1998)
Hanslin, K., J. Sjölin, P. Skorup, F. Wilske, R. Frithiof, A. Larsson, M. Castegren, E. Tano, and M. Lipcsey. “The impact of the systemic inflammatory response on hepatic bacterial elimination in experimental abdominal sepsis.” Intensive Care Medicine Experimental 7 (2019): art. 52. https://doi.org/10.1186/s40635-019-0266-x
Harte, A.L., N.F. da Silva, S.J. Creely, K.C. McGee, T. Billyard, E.M. Youssef-Elabd, G. Tripathi, E. Ashour, M.S. Abdalla, H.M. Sharada, A.I. Amin, A.D. Burt, S. Kumar, C.P. Day and P.G. McTernan. “Research Elevated endotoxin levels in non-alcoholic fatty liver disease.” Journal of Inflammation 7 (2010): 15-24. DOI: 10.1186/1476-9255-7-15
Hersoug, L.-G., P. Møller, and S. Loft. “Gut microbiota-derived lipopolysaccharide uptake and trafficking to adipose tissue: implications for inflammation and obesity.” Obesity Reviews 17 (2016): 297–312. DOI: 10.1111/obr.12370
Hurley, J. C. “Endotoxemia: Methods of detection and clinical correlates.” Clin. Microbiol. Rev. 8 (1995): 268–292. DOI: 10.1128/CMR.8.2.268
Kastner, A. “Untersuchungen zum Fettstoffwechsel und Endotoxin-Metabolismus bei Milchkühen vor dem Auftreten der Dislocatio abomasi.“ Inaug. Diss. Universität Leipzig, Veterinärmed. Fak. (2002). https://d-nb.info/967451647/34
Krüger M. “Escherichia coli: Problemkeim in der Nutztierhaltung.“ Darmflora in Symbiose und Pathogenität. Ökologische, physiologische und therapeutische Aspekte von Escherichia coli. 3. Interdisziplinäres Symposium. Alfred-Nissle-Gesellschaft (Ed.). Ansbach, 28.-29. Nov. (1997): 109-115.
Liang, Y.-C., H.-J. Liu, S.-H. Chen, C.-C. Chen, L.-S. Chou, and L. H. Tsai. “ Effect of lipopolysaccharide on diarrhea and gastrointestinal transit in mice: Roles of nitric oxide and prostaglandin E2.” World J Gastroenterol. 11 no. 3 (2005): 357–361. DOI: 10.3748/wjg.v11.i3.357
McAleer, J.P. and Vella, A.T. “Understanding how lipopolysaccharide impacts CD4 T cell immunity.” Crit. Rev. Immunol. 28 no. 4 (2008): 281-299. DOI:10.1615/CRITREVIMMUNOL.V28.I4.20
Morkok, A., L. Backstrom, L. Lund, A.R.Smith. “Bacterial endotoxin in blood of dysgalactic sows in relation to microbial status of uterus, milk, and intestine.” JAVMA 183 (1983): 786-789. PMID: 6629987
Munck, L.K., A. Mertz-Nielsen, H. Westh, K. Buxhave, E. Beubler, J. Rask-Madsen. “Prostaglandin E2 is a mediator of 5-hydroxytryptamine induced water and electrolyte secretion in the human jejunum.” Gut 29 no. 10 (1988): 1337-1341
Sampath, V.P. “Bacterial endotoxin-lipopolysaccharide; structure, function and its role in immunity in vertebrates and invertebrates.” Agriculture and Natural Resources 52 no. 2 (2018): 115-120. https://doi.org/10.1016/j.anres.2018.08.002
Seidler, T. “Freies Endotoxin in der Blutzirkulation von Schlachtschweinen: eine Ursache für bakterielle Translokationen?“ Diss. Universität Leipzig, Veterinärmed. Fak. (1998).
Smith, B.B. and W.C. Wagner. “Suppression of prolactin in pigs by Escherichia coli endotoxin.“ Science 224 no. 4649 (1984): 605-607
Wilken, H. “Endotoxin-Status und antioxidative Kapazität sowie ausgewählte Stoffwechselparameter bei gesunden Milch- und Mutterkühen.“ Inaugural Diss. Universität Leipzig (2003).
Ein komplexes Schlachtfeld: Mykotoxine im Magen-Darm-Trakt
Die meisten als Futterrohstoffe verwendeten Körner sind anfällig für Mykotoxin-Kontaminationen. Diese toxischen Sekundärmetaboliten werden vor oder nach der Ernte von Pilzen produziert und verursachen entlang der gesamten landwirtschaftlichen Wertschöpfungsketten schwere wirtschaftliche Verluste. Zu den negativen Folgen für Nutztiere zählen akute Auswirkungen wie Leber- und Nierenfunktionsstörungen, Erbrechen oder Anorexie sowie chronische Auswirkungen wie Immunsuppression, Wachstumsverzögerung und Reproduktionsprobleme. Das Management von Mykotoxinen hat daher für Tierproduzenten weltweit höchste Priorität.
Aber wie kommt es, dass Mykotoxine überhaupt solche Schäden verursachen? Dieser Artikel befasst sich mit den komplexen Prozessen, die auftreten, wenn Mykotoxine mit dem Magen-Darm-Trakt (GIT) in Kontakt kommen. Das Darmepithel ist das erste Gewebe, das Mykotoxinen ausgesetzt ist, und häufig in höheren Konzentrationen als andere Gewebe. Ein tieferes Verständnis darüber, wie Mykotoxine die GIT beeinflussen, ermöglicht es uns, die kaskadierenden Auswirkungen auf die Tiergesundheit und -leistung einzuschätzen, warum solche Schäden bereits bei Kontaminationsniveaus auftreten, die weit unter den offiziellen Sicherheitsschwellen liegen – und was wir dagegen tun können.
Das Darmepithel: die belebte Triage-Stelle für Nährstoffe und Schadstoffe
Wenn Mykotoxine aufgenommen werden, treffen sie auf das Darmepithel des GIT (Abbildung 1). Diese einzelne Zellschicht, die das Darmlumen auskleidet, erfüllt zwei widersprüchliche Funktionen: Erstens muss sie durchlässig genug sein, um die Aufnahme von Nährstoffen zu ermöglichen. Andererseits stellt es die primäre physiologische Barriere gegen Schadstoffe wie Viren, Mikroorganismen und Toxine dar.
Innerhalb des Darmepithels sind verschiedene Arten hochspezialisierter Zellen an der Epithelregeneration, der Nährstoffaufnahme, der angeborenen Abwehr, dem Transport von Immunglobulinen und der Immunüberwachung beteiligt. Die selektive Barrierefunktion bleibt aufgrund der Bildung komplexer Netzwerke von Proteinen erhalten, die benachbarte Zellen verbinden und den Interzellularraum versiegeln. Außerdem ist das Darmepithel mit Schleim bedeckt, der von Becherzellen produziert wird, die seine Oberfläche isolieren und die Adhäsion von Krankheitserregern an die Enterozyten (intestinale Absorptionszellen) verhindern.
Aufgrund seiner doppelten Beteiligung an Verdauungs- und Immunprozessen spielt das Darmepithel eine entscheidende Rolle für die allgemeine Gesundheit des Tieres. Wichtig ist, dass das Epithel direkt der gesamten Belastung mit aufgenommenen Mykotoxinen ausgesetzt ist. Daher können ihre Auswirkungen bereits bei geringen Konzentrationen problematisch sein.
Abbildung 1: Das Darmepithel
Problematische Wirkungen von Mykotoxinen auf das Darmepithel
Die meisten Mykotoxine werden im proximalen Teil des Magen-Darm-Trakts absorbiert (Tabelle 1). Diese Absorption kann hoch sein, wie im Fall von Aflatoxinen (~ 90%), aber auch sehr begrenzt sein, wie im Fall von Fumonisinen (<1%); außerdem hängt es von der Art ab. Wichtig ist, dass ein erheblicher Teil der nicht absorbierten Toxine im Lumen des Magen-Darm-Trakts verbleibt.
Einige der Mykotoxine, die in das Darmlumen gelangen, können durch die Wirkung bestimmter Bakterien in weniger toxische Verbindungen umgewandelt werden. Diese Wirkung findet jedoch überwiegend im Dickdarm statt – daher findet vor der Resorption in den oberen Teilen des GIT keine Entgiftung statt. Ein Teil der absorbierten Mykotoxine kann auch wieder in den Darm gelangen und die Zellen von der basolateralen Seite über den Blutkreislauf erreichen. Darüber hinaus treten sie durch den enterohepatischen Kreislauf (den Kreislauf von Substanzen zwischen Leber und Dünndarm) wieder ein. Beide Wirkungen erhöhen die Gesamtexposition des Magen-Darm-Trakts gegenüber den Toxinen.
Tabelle 1: Geschwindigkeits- und Absorptionsstellen verschiedener Mykotoxine
Die schädliche Wirkung von Mykotoxinen auf das Darmepithel erfolgt zunächst durch:
Eine Abnahme der Proteinsynthese, die die Barriere und die Immunfunktion verringert ( Van de Walle et al., 2010 )
Erhöhter oxidativer Stress auf zellulärer Ebene, der zu einer Lipidperoxidation führt und die Zellmembranen beeinflusst ( Da Silva et al., 2018 )
Veränderungen der Genexpression und der Produktion chemischer Botenstoffe (Zytokine) mit Auswirkungen auf das Immunsystem sowie das Zellwachstum und die Zelldifferenzierung ( Ghareeb et al., 2015 )
Die Induktion des programmierten Zelltods (Apoptose), die die Reposition von Immun- und Absorptionszellen beeinflusst ( Obremski & Poniatowska-Broniek, 2015 )
Wichtig ist, dass Studien, die auf realistischen Mykotoxinherausforderungen basieren (z. B. Burel et al., 2013 ), zeigen, dass die zur Auslösung dieser Prozesse erforderlichen Mykotoxinwerte niedriger sind als die von der EFSA, der Agentur für Lebensmittelsicherheit der Europäischen Union , als sicher gemeldeten Werte. Die letztendlichen Folgen reichen von einer verminderten Nährstoffaufnahme über Entzündungsreaktionen bis hin zu pathogenen Störungen beim Tier (Abbildung 2).
Figur 2: Auswirkungen von Mykotoxinen auf die GIT und Folgen für monogastrische Tiere
1. Veränderung der Morphologie und Funktionalität der Darmbarriere
Die Mykotoxine DON, Fumonisin und T2 induzieren eine Verringerung der Proliferations- und Differenzierungsrate der Epithelzellen. Dies führt zu einer Verringerung der Höhe und der Oberfläche der Darmzotten, was wiederum zu einer Verringerung der Nährstoffaufnahme führt. Zusätzlich werden einige Nährstofftransporter durch die Wirkung von Mykotoxinen wie DON und T2 gehemmt, die beispielsweise den Glucosetransport negativ beeinflussen.
Mehrere Studien weisen darauf hin, dass Mykotoxine wie Aflatoxin B1, DON, Fumonisin B1, Ochratoxin A und T2 die Permeabilität des Darmepithels von Geflügel und Schweinen erhöhen können (z. B. Pinton & Oswald, 2014 ). Dies ist hauptsächlich eine Folge der Hemmung der Proteinsynthese. Infolgedessen nimmt die Passage von Antigenen in den Blutkreislauf (z. B. Bakterien, Viren und Toxine) zu. Dies erhöht die Anfälligkeit des Tieres für infektiöse Darmerkrankungen. Darüber hinaus führt die Schädigung der Darmbarriere durch Mykotoxine dazu, dass sie auch schneller absorbiert werden.
2. Beeinträchtigte Immunfunktion im Darm
Der Darm ist eine sehr aktive Immunstelle, an der mehrere Immunregulationsmechanismen gleichzeitig den Körper vor Schadstoffen schützen. Immunzellen werden durch Mykotoxine durch die Einleitung von Apoptose, die Hemmung oder Stimulation von Zytokinen und die Induktion von oxidativem Stress beeinflusst. Studien zeigen, dass Aflatoxin, DON, Fumonisin, T2 und Zearalenon mit dem intestinalen Immunsystem derart interagieren, dass die Anfälligkeit des Tieres für virale und bakterielle Infektionen zunimmt (z. B. Burel et al., 2013 ). Darüber hinaus wird durch die Erhöhung der Kotausscheidung die horizontale Übertragung von Krankheitserregern verlängert.
Für die Geflügelproduktion ist eine der schwerwiegendsten enterischen Probleme bakteriellen Ursprungs die nekrotische Enteritis, die durch Clostridium perfringens- Toxine verursacht wird. Jedes Mittel, das das Magen-Darm-Epithel stören kann – z. B. Mykotoxine wie DON, T2 und Ochratoxin – fördert die Entwicklung einer nekrotischen Enteritis. Die durch Mykotoxine wie Aflatoxin, DON und T2 verursachte Hemmung des intestinalen Immunsystems wirkt sich ebenfalls auf die Entwicklung dieser Krankheit aus.
3. Veränderung der Darmflora
Der Magen-Darm-Trakt beherbergt eine vielfältige Gemeinschaft von Bakterien, Pilzen, Protozoen und Viren, die die Wände des distalen Teils des Darms auskleidet. Diese Mikrobiota verhindert das Wachstum pathogener Bakterien durch kompetitiven Ausschluss und die Sekretion natürlicher antimikrobieller Verbindungen, flüchtiger Fettsäuren und organischer Säuren.
Jüngste Studien zur Wirkung verschiedener Mykotoxine auf die Darmmikrobiota zeigen, dass DON und andere Trichothecene die Besiedlung coliformer Bakterien bei Schweinen begünstigen. DON und Ochratoxin A induzieren auch bei Vögeln und Schweinen eine stärkere Invasion von Salmonellen und deren Translokation in den Blutkreislauf und die lebenswichtigen Organe – selbst bei nicht zytotoxischen Konzentrationen. Es ist bekannt, dass Fumonisin B1 Veränderungen im Gleichgewicht von Sphingolipiden auf zellulärer Ebene induzieren kann, auch bei Magen-Darm-Zellen. Dies erleichtert die Adhäsion pathogener Bakterien, erhöht ihre Populationen und verlängert Infektionen, wie für den Fall von E. coli gezeigt wurde.
Aus Sicht der menschlichen Gesundheit ist die Besiedlung des Darms von Tieren, die Lebensmittel produzieren, durch pathogene Stämme von E. coli und Salmonellen von besonderer Bedeutung. Eine Mykotoxinexposition kann die Übertragung dieser Krankheitserreger erhöhen und ein Risiko für die menschliche Gesundheit darstellen.
4. Wechselwirkung mit bakteriellen Toxinen
Wenn Mykotoxine Veränderungen in der Darmmikrobiota induzieren, kann dies zu einer Erhöhung der Endotoxinkonzentration im Darmlumen führen. Endotoxine oder Lipopolysaccharide (LPS) sind Fragmente der Zellwände gramnegativer Bakterien. Sie werden während des Zelltods, des Wachstums und der Teilung von Bakterien freigesetzt. Daher sind Endotoxine auch bei gesunden Tieren immer im Darm vorhanden. Endotoxine fördern die Freisetzung mehrerer Zytokine, die eine verstärkte Immunantwort auslösen, Entzündungen verursachen und so den Futterverbrauch und die Leistung der Tiere, die Schädigung lebenswichtiger Organe, die Sepsis und in einigen Fällen den Tod der Tiere verringern.
Die Synergie zwischen Mykotoxinen und Endotoxinen kann zu einer Überstimulation des Immunsystems führen. Die Wechselwirkung zwischen Endotoxinen und Östrogenen wie beispielsweise Zearalenon führt zu chronischen Entzündungen und Autoimmunerkrankungen, da Immunzellen Östrogenrezeptoren aufweisen, die durch das Mykotoxin stimuliert werden. Andererseits wurde gezeigt, dass die Kombination von DON in geringen Konzentrationen und Endotoxinen im Darm zu einer Abnahme der transepithelialen Resistenz führt und das Gleichgewicht der Mikrobiota verändert.
Was ist zu tun? Proaktives Toxin-Risikomanagement
Um die schädlichen Folgen von Mykotoxinen für die Tiergesundheit und -leistung zu verhindern, sind proaktive Lösungen erforderlich, die die Verdauungs- und Immunfunktion des Darmepithels unterstützen und zur Aufrechterhaltung eines ausgewogenen Mikrobioms im GIT beitragen. Darüber hinaus ist es entscheidend, dass jedes Anti-Mykotoxin-Produkt sowohl Anti-Mykotoxin- als auch antibakterielle Toxineigenschaften aufweist und die Organe unterstützt, auf die Mykotoxine am meisten abzielen, z. B. die Leber. Die Mastersorb® Gold-Vormischung von EW Nutrition basiert auf der synergistischen Kombination von natürlichen Tonmineralien, Hefezellwänden und Phytomolekülen. Seine Wirksamkeit wurde ausführlich getestet, auch als Mittel zum Umgang mit E. coli- Endotoxinen.
Mastersorb® Gold: Die Anti-Mykotoxin-Aktivität stabilisiert die Leistung und stärkt die Gesundheit der Leber
Ein in Deutschland an männlichen Ross 308-Broilern durchgeführter Feldversuch zeigte, dass bei Broilern, die eine mit DON und Zearalenon kontaminierte Diät erhielten, die Zugabe von 1 kg Mastersorb® Gold pro Tonne Futter zu einer signifikanten Leistungssteigerung führte. Sie haben nicht nur den Mykotoxin-induzierten Gewichtsverlust (6% Anstieg gegenüber der Gruppe, die nur die Herausforderung erhielt) wiederhergestellt, sondern auch gegenüber der Kontrollgruppe (die weder die Herausforderung noch Mastersorb® Gold erhielt) zugenommen. Die Futterverwertung verbesserte sich auch um 3% im Vergleich zu der mit Mykotoxinen belasteten Gruppe.
Eine wissenschaftliche Studie an weiblichen Kreuzungsschweinen zeigte, dass Mastersorb® Gold die schädlichen Auswirkungen der Fumonisin-Kontamination im Futter signifikant reduzierte. Die Abnahme der Gewichtszunahme und die Abnahme der Futterverwertung konnten um 6,7% bzw. 13 FCR-Punkte gemildert werden (Abbildung 3). Auch das Sphinganin / Sphingosin (Sa / So) -Verhältnis, ein Biomarker für das Vorhandensein von Fumonisin im Blutserum, konnte um 22,5% verringert werden.
Figur 3: Mastersorb® Gold steigert die Leistung von Schweinen, die mit Fumonisin kontaminiert gefüttert werden
Eine weitere Studie an weiblichen Kreuzungsferkeln, die an einer deutschen Universität durchgeführt wurde, untersuchte, ob Mastersorb® Gold die Leistung sowie die Gesundheit der Leber unter einer natürlich vorkommenden Herausforderung von ZEA (~ 370ppb) und DON (~ 5000ppb) unterstützen kann. Mastersorb® Gold verbesserte die Gewichtszunahme und Futterverwertung bei Ferkeln, die die mit Mykotoxin kontaminierte Nahrung erhielten, signifikant: Die tägliche Körpergewichtszunahme war 75 g höher als bei einer Gruppe, die nur die Herausforderung erhielt, und die FCR verbesserte sich um 24% (1,7 vs. 2,25 für die Gruppe ohne Mastersorb® Gold). Darüber hinaus verbesserte Mastersorb® Gold das Lebergewicht (insgesamt und relativ) und die AST-Werte der Ferkel (Aspartat-Aminotransferase, ein Enzym, das auf eine Leberschädigung hinweist) signifikant. Es zeigte sich auch eine Tendenz zur Verbesserung des Milzgewichts und der GGT-Spiegel (Gamma-Glutamyltransferase, ein weiteres Enzym, das auf Leberprobleme hinweist), was darauf hinweist, dass Mastersorb® Gold den schädlichen Auswirkungen der Mykotoxin-Kontamination auf die Leberfunktionalität wirksam entgegenwirkt.
In-vitro-Studien belegen die Wirksamkeit von Mastersorb® Gold sowohl gegen Mykotoxine als auch gegen bakterielle Toxine
Tierfutter ist häufig mit zwei oder mehr Mykotoxinen kontaminiert, weshalb es wichtig ist, dass ein Anti-Mykotoxin-Mittel gegen eine Vielzahl verschiedener Mykotoxine wirksam ist. Um zu verhindern, dass Mykotoxine die GIT schädigen, sollte ein wirksames Produkt idealerweise die meisten Mykotoxine im ersten Teil des Darms des Tieres (unter sauren Bedingungen) adsorbieren. In-vitro-Experimente in einer unabhängigen Forschungseinrichtung in Brasilien zeigten, dass eine Anwendung von 0,2% Mastersorb® Gold alle getesteten Mykotoxine mit Raten von 95 bis 97% bei einem pH-Wert von 3 unter Verwendung realistischer Herausforderungen von 1000 ppb (Aflatoxin B1 und ZEA) bindet. und 2500 ppb (Fumonisin B1 und DON). Besonders ermutigend sind die Bindungsergebnisse für Fumonisin und DON, die unter schwierigen, nahezu neutralen Bedingungen (pH 6) häufig als „nicht bindend“ angesehen werden.
Figur 4: Mastersorb® Gold-Bindungskapazität gegen verschiedene Mykotoxine (%)
In Bezug auf die Wirksamkeit gegen Endotoxine hat eine In-vitro- Studie, die unter anderem an der Universität Utrecht durchgeführt wurde, gezeigt, dass Mastersorb® Gold ein starkes Instrument gegen das von E. coli freigesetzte LPS ist . Für den Test wurden vier Premium-Mykotoxin-Bindemittel in einer Phosphatpufferlösung in Konzentrationen von 0,25% und 1% suspendiert. E. coli LPS wurden in jeder Probe von 50 ng / ml bis zu einer Endkonzentration suspendiert. Gegen diese besonders hohe Herausforderung erreichte Mastersorb® Gold eine Bindungsrate von 75% bei einer Einschlussrate von 1%: eine deutliche Outperformance gegenüber Konkurrenzprodukten, die bestenfalls eine Bindungsrate von 10% aufwiesen.
Fazit
Ein gesunder Magen-Darm-Trakt ist für die allgemeine Gesundheit der Tiere von entscheidender Bedeutung: Er sorgt für eine optimale Nährstoffaufnahme, schützt durch seine Immunfunktion wirksam vor Krankheitserregern und ist der Schlüssel zur Aufrechterhaltung einer ausgewogenen Mikroflora. Selbst in Mengen, die von der Europäischen Union als sicher eingestuft werden, können Mykotoxine verschiedene Darmfunktionen wie Absorption, Permeabilität, Immunität und Mikrobiota-Gleichgewicht beeinträchtigen, was zu einer geringeren Produktivität und Anfälligkeit für Krankheiten führt.
Um die Leistung der Tiere zu gewährleisten, ist es wichtig, kontinuierlich nach einer geringen Kontamination der Futterrohstoffe zu streben – und zu verhindern, dass die unvermeidbaren Mykotoxinbelastungen das Darmepithel durch die Verwendung eines wirksamen Antimykotoxin-Mittels schädigen, das auch Tiere gegen Endotoxine unterstützt und steigert die Leberfunktion. Untersuchungen zeigen, dass Mastersorb® Gold ein leistungsstarkes Werkzeug für proaktive Produzenten ist, die eine stärkere Tiergesundheit, Tierschutz und Produktivität suchen.
Von Marisabel Caballero und Sabria Regragui Mazili