Environmental Stress and Mycotoxins in Breeders: Hidden Losses in Fertility, Egg Quality & Chick Output
Author: Dr. Vaibhav Gawande, Feed Safety and Toxin Control Specialist – South Asia, EW Nutrition
In Tropical countries like India, Bangladesh & Sri Lanka, high temperatures and humidity significantly increase the risk of mycotoxin contamination in animal feed. FAO surveys report mycotoxin contamination in over 70% of cereals and oilseeds used for animal feed, making them a major threat to poultry health and productivity.
The problem becomes particularly severe during the summer & monsoon when:
The new maize crop with high moisture enters the market
Drying is often inadequate
Feed mill humidity remains high
The Science of Mycotoxins & Environmental Risk
Summer temperatures (28–38°C) and high humidity promote fungal growth and mycotoxin production in feed ingredients.
Freshly harvested “new maize” often enters feed mills with unsafe moisture levels (13–15%), exceeding the safe storage limit of 11–12%.
High-moisture grain undergoes self-heating during storage, creating ideal conditions for fungal proliferation.
Poor aeration, condensation, insect damage, and humid storage environments further accelerate contamination.
Common summer mycotoxins include Aflatoxins, Ochratoxins, and T-2 toxins, which may develop during both pre- and post-harvest stages.
Although pelleting and heat treatment may destroy molds, mycotoxins are generally heat-stable and remain toxic to poultry.
Regional Case Studies: South Asia
India
Coastal Humidity Crisis – Andhra Pradesh, Telangana, Tamil Nadu, Odisha & West Bengal
Pre-monsoon humidity promotes Aspergillus flavus growth in stored maize.
Aflatoxin contamination commonly reduces shell quality and egg production.
Fatty Liver Hemorrhagic Syndrome (FLHS) is frequently observed.
Short-Storage Trap – Punjab & Haryana
Wet maize stored during summer heat retains internal moisture, favoring T-2 toxin and Ochratoxin formation.
Broilers commonly show oral lesions, feed refusal, and poor FCR.
Mixed Toxin Challenge – Maharashtra
Co-contamination with Aflatoxin and Fumonisin is common.
Combined toxicity causes immunosuppression, poor vaccine response, and increased mortality during disease outbreaks.
Bangladesh
High Humidity Feed Risk
Persistent humidity and post-flood harvesting increase moisture retention in maize and rice by-products.
Aflatoxin frequently co-occurs with Ochratoxin A.
Causes immunosuppression, uneven flock uniformity, and poor hatchability in breeders.
Sri Lanka
Tropical Storage Challenge
Tropical coastal humidity and prolonged ingredient storage favor fungal proliferation.
Aflatoxin and Fumonisin commonly develop during humid transit and storage.
Causes thin shells, liver damage, and poor FCR.
Nepal
Mountain Moisture Variability
Humid Terai grains stored in cool hill regions favor mixed mycotoxin contamination.
Aflatoxins commonly co-occur with DON and Zearalenone.
DON causes feed refusal, while Zearalenone induces prolapse and false layers.
Mycotoxins and High Temperature Humidity Index (THI): Synergistic effects on poultry health, immunity & productivity
The Immunological “Blackout”
Aflatoxins, Trichothecenes, and Ochratoxins inhibit protein synthesis, reducing the formation of antibodies and immune cells.
Mycotoxins cause atrophy of immune organs (bursa of Fabricius, thymus, and spleen)
Macrophage activity and phagocytosis are reduced, weakening bacterial clearance.
Cytokine signaling is disrupted, delaying immune activation against infections.
Enhanced oxidative stress: Mycotoxins increase the occurrence of reactive oxygen species (ROS), and heat stress weakens antioxidant defenses, resulting in severe cellular and liver damage.
Oxidative stress caused by aflatoxins and trichothecenes leads to immune cell apoptosis and tissue damage.
Gut Health & Barrier Damage
The gut is the first line of defense. Mycotoxins and heat stress act like a “chemical and physical abrasive” on the intestinal lining.
Villi Destruction: T-2 and Aflatoxins cause necrosis (cell death) of the intestinal villi. This reduces the surface area for nutrient absorption, leading to poor FCR.
The “Leaky Gut” Phenomenon: Heat stress causes blood to be diverted from the internal organs to the skin for cooling (vasodilation). As a result, the gut receives less oxygen, causing the tight junctions (the “glue” between intestinal cells) to break down and become permeable. Mycotoxins also have a direct effect, inhibiting tight junctions proteins.
Pathogen Entry: Mycotoxins further erode the protective mucus layer. With the “gates” (tight junctions) open and the “walls” (mucus) gone, bacteria can freely enter the bloodstream.
Disease Susceptibility
Because the immune system is “blind” and the gut is “leaky,” the bird becomes a target for opportunistic infections.
Secondary Bacterial Infections: Normal gut bacteria like E. coli and Salmonella transition from harmless to fatal, causing systemic septicemia.
Viral Synergism: Small viral loads, such as Inclusion Body Hepatitis (IBH) that a healthy bird would normally survive, become highly fatal.
Coccidiosis Flare-ups: Damaged gut linings are more easily colonized by Eimeria, making standard anti-coccidial programs less effective.
Vaccine Failure:
Mycotoxins suppress B-cell and T-cell maturation, reducing vaccine effectiveness.
Low immunoglobulin (IgG, IgA, IgM) production results in poor antibody titers.
Maternal toxin exposure reduces immunity transfer to chicks, increasing early-age disease vulnerability.
Figure 1: How mycotoxins and heat stress cause damage in poultry
Breeder Reproductive Dysfunction and Transgenerational Effects of Mycotoxins
In breeder operations, mycotoxins represent a catastrophic economic threat because they are vertically transmitted. Unlike commercial layers, where the loss is limited to the individual bird’s production, breeder contamination compromises the viability of the entire next generation.
1. Impact on the Reproductive Systems (Male & Female)
Mycotoxins hit both sides of the fertility equation, often exacerbated by summer heat.
Female Reproductive System
Mode of action: Mycotoxins (especially Zearalenone) mimic estrogen. This disrupts the hypothalamic-pituitary-ovarian axis.
Impact: inflammation of the oviduct, cystic ovaries, and reduced synthesis of yolk precursors in the liver, resulting in a sharp drop in egg production and poor internal egg quality.
Male Fertility
Mode of action: Toxins like T-2 and Aflatoxin induce oxidative stress that damages the phospholipid membrane of sperm cells.
Impact: Under heat stress, rooster semen quality already declines; mycotoxins accelerate this by reducing sperm motility, concentration, and increasing morphological abnormalities. This leads to a massive spike in infertility rates.
2. Hatchability & Embryonic Mortality
For breeders, mycotoxins represent a “generational loss” via vertical transmission.
Mode of action (the yolk bridge): Some mycotoxins are highly lipophilic. As the liver assembles the yolk, it deposits toxins directly into the egg.
The “three-wave” mortality:
Early (Days 1–7): Toxins interfere with mitosis (cell division), leading to early deaths often mistaken for “infertility.”
Mid-Term (Days 8–18): As the embryo begins intensive absorption of the toxic yolk, its developing liver and kidneys are compromised. This is the classic “Toxin Fingerprint.”
Late (Days 19–21): Ochratoxins impair the embryo’s ability to mobilize calcium from the eggshell. As a result, the chick becomes too weak to pip and dies fully developed inside the shell (“dead-in-shell”).
3. Chick Quality, Grading, and Settability
The “Chick Quality” starts in the breeder’s gut and kidney health.
Mode of action (nutrient malabsorption): Mycotoxins reduce pancreatic lipase and bile salts. This prevents the mother from absorbing fat-soluble vitamins (A, D, E, K) and pigments. They also lead to lower intestinal adsorption due to a reduced absorption area and lower transporter efficacy.
Impact on chick quality:
“Pale Bird Syndrome”: Chicks lack vital carotenoids for early-stage defense.
Skeletal weakness: Interference with Vitamin D3 metabolism results in weak legs and “rubbery beaks” in Day-Old-Chicks (DOCs).
High first week mortality (FWM): Chicks hatch immunosuppressed, leading to high mortality during the first week.
Impact on egg grading & settability: Mycotoxins (Ochratoxin) are nephrotoxic, damaging the kidneys and disrupting the blood calcium-carbonate balance. This leads to “Sandpaper” shells, misshapen eggs, and a 5–10% drop in the number of settable eggs fit for the incubator.
Figure 2: The impact of mycotoxins on breeder production and economics
Pro tip for breeders: In summers, high-moisture new maize triggers a mycotoxin surge that synergistically destroys the breeder’s kidneys and shell gland, crippling egg settability. A 5% spike in “Dead-in-Shell” embryos during breakout analysis is a definitive indicator of feed toxicity rather than incubator failure.
Integrated Mycotoxin Mitigation Strategies for Poultry Production
To pursue an effective mycotoxin mitigation strategy, it is essential first to identify which mycotoxins are relevant to a given region before implementing measures.
Strict moisture control: Reject any maize arriving with >14% moisture.
Rapid screening: Perform rapid mycotoxin screening before unloading raw materials.
Mechanical grain driers: To maintain a safe storage moisture level (<12%)
2. Feed Plant & Storage Hygiene
First-In, First-Out (FIFO): Ensure strict inventory rotation to prevent “pockets” of old, moldy feed from contaminating new batches.
Frequently clean silos and elevators: High temperature and humidity cause moisture condensation on silo walls, leading to localized mold growth.
Antifungal Treatment: Use buffered organic acids (propionic and formic acid) to limit mold proliferation in feed.
3. Broad-Spectrum binders:
Bentonites (for Aflatoxins) and Yeast Cell Walls: These components help bind pathogenic bacteria like E. coli that capitalize on the “leaky gut” caused by toxins and heat stress.
4. Physiological & Gut Health Support
Water acidification: Lower the drinking water pH to 4.5–5.5. This prevents bacterial blooms in the water lines when birds increase water intake by 3 times during heat stress.
Liver & kidney tonics: Supplemental hepatic (milk thistle/silymarin) and renal support to help the bird metabolize and export toxins more efficiently.
Metabolite supplementation: Use 25-hydroxyvitamin D3 in breeder diets to bypass the liver/kidney damage and ensure shell quality remains intact.
Antioxidant boost: Increase levels of Vitamin E, C, and Selenium to counter the oxidative stress caused by the heat-toxin synergy.
5. Monitoring & Diagnostics
Hatchery breakout analysis: Monitor “dead-in-shell” embryos. A spike in mid-term mortality is an immediate indicator that the breeder feed toxin binder needs a dosage increase.
Frequent lab testing: Mycotoxin testing at least weekly during the new maize transition to identify the specific toxin profile.
Solutions are available to support toxin risk management
In the challenging climate where high-moisture “new maize” and summer humidity create a complex cocktail of mycotoxins, endotoxins, and pesticide residues, traditional, single-ingredient binders often fall short. Modern poultry production requires a proactive solution that does more than just “bind”; it must protect the bird’s internal integrity.
Solis Max – The effective myco- and endotoxin solution for sustained profitability
Solis Max is engineered to meet customers’ demand for an effective solution, offering a multi-pronged defense mechanism that targets the root causes of performance collapse. Solis Max uses a synergistic blend of five key components to ensure the flock’s safety.
Trials prove the effectiveness of Solis Max
Solis Max shows dose-dependent adsorbing capacity against multiple mycotoxins:
Figure 4: Mycotoxin Binding Capacity Of Solis Max
SOLIS MAX shows endotoxin adsorbing capacity – 1mg of SOLIS MAX absorbs 20 endotoxin units (EU) of E. coli endotoxin (80% adsorption rate):
Figure 5: Endotoxin Binding Capacity Of Solis Max
Solis Max demonstrates high pesticide binding efficiency across multiple compounds:
Figure 6: Pesticide Net Binding Capacity Of Solis Max (%)
Conclusion:
The convergence of a high Temperature–Humidity Index (THI) and mycotoxicosis represents a critical, multisystem challenge in poultry production, precipitating severe pathology across the hepatic, renal, and gastrointestinal systems. In breeding operations, this crisis exhibits a transgenerational impact: lipophilic mycotoxins are vertically transmitted to the yolk, inducing mid-term embryonic mortality and compromising post-hatch progeny immunity.
Mitigation demands stringent control of raw material moisture alongside advanced, broad-spectrum interventions. Utilizing an advanced multi-pronged solution like Solis Max counters this synergy by providing physicochemical adsorption and targeted organ protection. By neutralizing the concurrent threats of mycotoxins, endotoxins, and pesticides, it preserves cellular integrity, mitigates systemic pathology, and maintains optimal performance under extreme environmental stress.
References available upon request.
Learning from AGP mechanisms to advance poultry nutrition
By Ilinca Anghelescu, Dr. Andreas Michels, Predrag Persak
Our understanding of how nutrition influences growth and resilience in poultry has greatly expanded in recent years. It is now clear that animal performance stems to a large extent from a balance between metabolism, immune function, and the gut microbiome. These systems interact continuously, and even small nutritional or environmental changes can shift the animals’ physiological response. This growing knowledge has encouraged the development of nutritional strategies and feed components that work through adaptive, non-antibiotic mechanisms. One recent proposed explanation for these responses has rapidly gained ground: hormetic modeling.
Hormetic modeling describes how small or moderate doses of nutritional components can activate beneficial adaptive responses (improved resilience or metabolic efficiency), while excessive doses become harmful. This idea parallels, largely speaking, Paracelsus’s famous principle: “The dose makes the poison.” In poultry nutrition, such hormetic patterns are well recognized in nutrients like trace elements (selenium, zinc) and specific amino acids (for example, arginine). At optimal levels, these nutrients support antioxidant defense, growth, and immune balance, whereas excessive intake may cause oxidative or metabolic stress
This review examines the hormetic principle and its application to modern poultry/swine feeding concepts, exploring how balanced nutrient design and controlled inclusion of bioactive compounds can strengthen cellular adaptation, improve stress tolerance, and enhance production efficiency.
How do AGPs actually work?
Despite AGP’s widespread historical use, the precise mechanisms by which subtherapeutic doses of antibiotics enhance animal productivity remained poorly understood. Recent advances in systems biology and mitochondrial research propose new answers, much needed to develop future advanced nutritional systems.
The traditional explanations for AGP efficacy have focused primarily on antimicrobial effects:
reducing nutrient competition from microorganisms
decreasing harmful bacterial metabolites
improving gut wall morphology (thinner gut wall ➡ better nutrient absorption)
preventing subclinical infections
However, these mechanisms alone could not fully explain why different classes of antibiotics with diverse mechanisms of action produce similar growth-promoting effects (Gutierrez-Chavez et al., 2025).
Niewold (2007) hypothesized that the primary mechanism of AGPs is non-antibiotic anti-inflammatory activity, reducing the energetic costs of chronic low-grade inflammation. Inflammation diverts nutrients from growth toward immune responses, with cytokine production (particularly IL-1β, IL-6, and TNF-α) suppressing anabolic pathways (Kogut et al., 2018). AGPs appear to selectively inhibit pro-inflammatory cytokine production without completely suppressing immune function.
A paper published in 2024 by Fernandez Miyakawa et al. proposes that antibiotics at subtherapeutic levels act primarily through mitochondrial hormesis and adaptive stress responses, and not simply through antimicrobial activity. In this model, mitochondria act as bioenergetic hubs and signaling centers. Low-dose antibiotics trigger mild mitochondrial stress, which triggers the activation of adaptive protective pathways.This in turn induces mitokine release, leading to systemic adaptive responses improving growth, feed efficiency, and disease tolerance.
Mechanism of action in the hormetic model of AGP efficiency
Hormesis is a biphasic mechanism whereby high doses are toxic, but low doses stimulate adaptive responses and are beneficial. In the case of AGPs, Fernandez Miyakawa et al. propose that low doses stimulate growth, stress resistance, and cellular repair.
Key signaling pathways
As Bottje et al. (2006, 2009) shows, efficient animals often have mitochondrial inner membranes that are less permeable to protons and other ions, allowing for more effective coupling between electron transport and ATP synthesis, which reduces energy loss through proton leak and maximizes the production of ATP per oxygen molecule consumed. Lower membrane permeability is influenced by factors like decreased membrane surface area per protein mass, specific membrane protein content (such as adenine nucleotide translocase), and fatty acid composition in the membrane phospholipids, all contributing to a tighter barrier that prevents unregulated electron or proton flow and supports higher energetic efficiency. Such features make mitochondria in efficient species more capable of maintaining membrane integrity and ATP generation, especially when facing environmental stress, as seen in freeze-tolerant animals whose mitochondria do not undergo damaging permeability transitions under extreme conditions.
Nrf2
Many AGPs interfere with mitochondrial protein synthesis and electron transport chain. At subtherapeutic levels, they cause a mild ROS increase, which triggers the activation of redox-sensitive transcription factor Nrf2. Since Nrf2 regulates over 250 antioxidant, detoxification, and anti-inflammatory genes, the result is improved cell survival, redox balance, and tolerance to stress.
Figure 1 From Zhang et al., 2024
Mitokine production
Mitokines are “signaling molecules that enable communication of local mitochondrial stress to other mitochondria in distant cells and tissues” (Burtscher 2023). Through fibroblast growth factor 21 (FGF21), growth differentiation factor 15 (GDF15), adrenomedullin2 (ADM2) etc, these stress signals are released systemically and coordinate tissue-wide responses, leading to improved growth and resilience.
Inflammation and disease defense
While the negative side of antibiotic growth promoters is well researched and understood (Rahman et al., 2022), science can advance by isolating the positive effects and attempting to offer different pathways to the same benefits. One such lesson can be derived from understanding inflammation pathways and responses.
Chronic low-grade intestinal inflammation is common in modern poultry production, due to diet, microbiota shifts, high metabolic demands etc. This inflammation diverts energy from growth to immune responses.
AGPs reduce the energy costs of this inflammation in three main ways:
Reduces inflammation through adaptive stress response
Raising the threshold to trigger inflammation
Promoting overall resilience, rather than simply killing pathogens
Fernandez Miyakawa et al. suggest, in this emerging model, that disease defense can operate two different actions: resistance to health challenges through reduction of the pathogen load (which is driven by the immune system and is energy costly); and overall resilience by reducing host damage without reducing the pathogen load. AGPs, the authors claim, mainly promote resilience by enhancing mitochondrial stress responses and tissue repair, i.e. more precisely:
Direct mitochondrial stimulation in intestinal epithelial cells
Metabolic optimization supporting growth and feed efficiency
Figure 2 From Fernandez Miyakawa et al., 2024.
In this context, “metabolic optimization” refers to the enhancement of metabolic processes within livestock or poultry to support efficient growth, feed conversion, and physiological resilience, without relying on immune-mediated pathways that are energetically costly. Scientific evidence shows that metabolic optimization involves improving nutrient assimilation, promoting more efficient energy production in tissues (such as mitochondrial ATP synthesis), and minimizing wasteful metabolic byproducts, resulting in reduced feed intake per unit of growth and better utilization of dietary nutrients (Rauw 2025, El-Hack 2025).
Function of feed additives and feed components
Feed additives and feed components in many ways represent the complete other side of the spectrum from antibiotics, but are there some features where antibiotics and feed additives come close in their functions? There is a good case to be made for certain feed additives ultimately working in the animal to achieve similar benefits to the desirable, non-medicinal usage of AGP´s. Especially with the emergent model of AGP mechanism described above, it is worth discussing how certain feed additives can support the same end goal: promoting animal resilience.
Lillejhoj et al (2018), Gutierrez-Chavez et al. (2025) and others outline the end-results such products must achieve:
Growth performance & feed conversion efficiency
Promotion of animal productivity under real-world conditions
Support gut homeostasis
Non-adverse effect on the immune system
Reduction of oxidative stress
Support organism in mitigation of enteric inflammatory consequences
Within the hormetic model, possibly the most important systemic benefit is, in one phrase, promoting resilience. Phytomolecules have long been used, in human and animal medicine, for the same end goal. The mechanisms described below should naturally be seen with caution, as phytomolecule microbiome effects can be subtler and context-dependent. However, the substantiating literature has been increasingly accumulating on these specific topics.
1. Immunometabolic regulation
Phytomolecules demonstrate remarkably similar anti-inflammatory effects to what Niewold (2007) suggested was a primary mechanism of AGPs: non-antibiotic anti-inflammatory activity, reducing the energetic costs of chronic low-grade inflammation. Inflammation diverts nutrients from growth toward immune responses, with cytokine production (particularly IL-1β, IL-6, and TNF-α) suppressing anabolic pathways (Kogut et al., 2018). AGPs appear to selectively inhibit pro-inflammatory cytokine production without completely suppressing immune function. A similar effect can be observed with various types of phytomolecules, which significantly reduced pro-inflammatory and/or increased anti-inflammatory cytokine expression in animals challenged with several pathogens. The anti-inflammatory mechanism appears to involve inhibition of NF-κB activation and modulation of MAPK signaling pathways (Kim et al., 2010; Long et al., 2021).
2. Mitochondrial hormesis and energy metabolism
Fernández Miyakawa et al. (2024, see above) proposed that AGPs exert growth-promoting effects through mitochondrial hormesis – subtherapeutic antibiotic doses induce mild mitochondrial stress, triggering adaptive responses that enhance mitochondrial function, energy metabolism, and cellular resilience. This mechanism, while requiring further validation, explains why different antibiotics with diverse targets produce similar growth outcomes.
The mitochondrial stress response involves activation of the IL-6 receptor family signaling cascade, which regulates metabolism, growth, regeneration, and homeostasis in liver and other tissues (Perry et al., 2024). Subtherapeutic antibiotic exposure activates proteins involved in growth and proliferation through IL-6R gp130 subunit signaling, including JAK, STAT, mTOR, and MAPK pathways.
Phytomolecules demonstrate similar mitochondrial effects. Perry et al. (2024) showed that increased activity of AMPK, mTOR, PGC-1α, PTEN, HIF, and S6K can also be available via phytomolecule activity, suggesting enhanced anabolic metabolism.
Capsicum oleoresin supplementation in broilers increased jejunal lipase and trypsin activity, enhanced ileal amylase activity, improved jejunal morphology, and modulated immune organ development, indicating enhanced digestive efficiency and nutrient utilization (Li et al., 2022).
Compounds such as vanillin, thymol, eugenol have been shown to improve glucose and lipid metabolism through TRPV1 activation and mitochondrial function enhancement (Gupta et al., 2022; Zhang et al., 2017).
3. Gut microbiota modulation
AGPs selectively reduce specific microbial populations, particularly Lactobacillus species that produce bile salt hydrolase (BSH). Since BSH reduces fat digestibility and thus weight gain, AGP-mediated reduction of BSH-producing bacteria enhances energy extraction and growth (Lin, 2014; Bourgin et al., 2021).
Recent research by Zhan et al. (2025) using single-molecule real-time 16S rRNA sequencing demonstrated that therapeutic antibiotic doses (lincomycin, gentamicin, florfenicol, benzylpenicillin, ceftiofur, enrofloxacin) significantly altered chicken gut microbiota composition, with Pseudomonadota and Bacillota becoming dominant phyla after exposure. Different antibiotics produced distinct temporal effects on microbial diversity and community structure.
Phytomolecules exert targeted antimicrobial effects while promoting beneficial bacteria. Dietary supplementation with 800 mg/kg Capsicum extract in Japanese quails reduced cecal counts of pathogenic bacteria (Salmonella spp., E. coli, coliforms) while modulating Lactobacilli populations (Reda et al., 2020).
In pigs, 80 mg/kg natural capsicum extract increased cecal propionic acid and total volatile fatty acid concentrations, with increased butyric acid in the colon – indicating enhanced fermentation by beneficial bacteria (Long et al., 2021).
Capsicum and Curcuma oleoresins altered intestinal microbiota composition in commercial broilers challenged with necrotic enteritis, reducing disease severity through microbiome modulation (Kim et al., 2015).
Capsaicin demonstrates selective antimicrobial activity, inhibiting pathogenic Gram-negative bacteria while favoring development of certain Gram-positive bacteria. The antibacterial mechanism involves induction of osmotic stress and membrane structure damage (Adaszek et al., 2019; Rosca et al., 2020).
4. Intestinal barrier function and gut health
AGPs have been associated with improved intestinal morphology, including increased villus height and reduced crypt depth, which enhance absorptive capacity (Gaskins et al., 2002).
Phytomolecules produce similar or superior effects. Capsicum extract (80 mg/kg) in pigs increased ileal villus height and upregulated MUC-2 gene expression, indicating enhanced gut barrier function and integrity. The improved barrier function correlated with reduced diarrhea incidence (Liu et al., 2013; Long et al., 2021).
Allium hookeri extract increased expression of tight junction proteins (claudins, occludins, ZO-1) in LPS-challenged broiler chickens, demonstrating direct enhancement of barrier integrity (Lee et al., 2017).
5. Oxidative stress mitigation
Oxidative stress impairs growth by damaging cellular components and triggering inflammatory responses. AGPs reduce oxidative stress indirectly through anti-inflammatory effects and microbiota modulation (Bortoluzzi et al., 2021).
Phytomolecules possess direct antioxidant properties. Capsicum extract (50 mg/kg) in heat-stressed quails reduced serum and ovarian malondialdehyde (MDA) while increasing superoxide dismutase (SOD) and catalase (CAT) activities. Ovarian transcription factors showed decreased NF-κB and increased Nrf2 and HO-1 expression (Sahin et al., 2016).
A mixture of herbal extracts including pepper reduced thiobarbituric acid reactive substances and MDA in broiler liver and muscle, while increasing glutathione peroxidase (GSH-Px) activity and improving antioxidant enzyme expression (Saleh et al., 2018).
Capsicum extract (80 mg/kg) in pigs increased total antioxidant capacity, SOD, and CAT while reducing MDA levels, demonstrating robust antioxidant effects (Long et al., 2021).
Standardization and controlled release: Critical success factors
A major criticism of phytomolecules has been inconsistent efficacy across studies. However, this variability largely reflects differences in:
Active compound concentrations
Bioavailability and stability
Dosing precision
Product quality and standardization
Microencapsulation is one of the technologies that address the standardization and bioavailability challenges. It protects volatile compounds from degradation during feed processing and storage, with encapsulated essential oils showing significantly higher retention compared to unprotected forms (Stevanović et al., 2018). By creating a protective barrier around active ingredients, microencapsulation enables controlled release in specific regions of the gastrointestinal tract, improving absorption efficiency and reducing dose variability (Bringas-Lantigua et al., 2011). The technology also masks unpalatable flavors that can reduce feed intake while standardizing active ingredient concentrations through precise manufacturing processes (Gharsallaoui et al., 2007). Studies demonstrate that spray-dried microencapsulated essential oils achieve encapsulation efficiencies exceeding 93% with minimal loss during storage (Hu et al., 2020), and can be engineered for enzyme-mediated release to ensure bioactive delivery at optimal intestinal sites (Elolimy et al., 2025).
Mechanistic synthesis: An integrated model
The evidence indicates that both AGPs and phytomolecules operate through an integrated network of effects:
Primary Level: Selective antimicrobial effects modify gut microbiota composition
This integrative model explains why multiple antibiotics with different mechanisms produce similar growth outcomes: they converge on common pathways regulating immunometabolism and mitochondrial function (Fernández Miyakawa et al., 2024).
Phytomolecules operate through the same mechanistic framework but with potential advantages:
Safety and antimicrobial resistance considerations
Antibiotic exposure significantly disrupts gut microbiota diversity and stability, with effects persisting beyond withdrawal periods. The study by Zhan et al. (2025) demonstrated that different antibiotics produce varying degrees of microbiota disruption, with florfenicol and gentamicin showing the strongest and most persistent effects.
In contrast, phytomolecules generally do not generate resistance through the same mechanisms as antibiotics. Some phytochemicals may actually enhance antibiotic efficacy and resensitize resistant bacteria through structural modifications of bacterial membranes (Khameneh et al., 2021; Suganya et al., 2022).
However, one study reported increased correlation between antibiotic resistance genes (ARGs) and mobile genetic elements in pig feces after mushroom powder supplementation, suggesting that certain phytogenic compounds may increase ARG mobility (Muurinen et al., 2021). This emphasizes the need for continued surveillance of phytomolecule effects on resistance gene dynamics.
Capsaicinoids and capsinoids have well-established safety profiles. Capsiate, a non-pungent analogue of capsaicin, exhibits substantially lower toxicity while maintaining similar metabolic and growth-promoting effects (Gupta et al., 2022). No adverse effects on animal health or product quality have been reported at recommended dosages in reviewed studies.
Future directions and research needs
Despite substantial progress, several areas require further investigation:
Mechanistic refinement: Detailed characterization of signaling pathways, particularly the IL-6R/gp130 cascade and mitochondrial stress responses
Precision formulation: Development of combinations optimized for specific production stages, environmental conditions, and disease pressures
Bioavailability optimization: Advanced delivery systems ensuring consistent active compound release and absorption
Microbiome-host interaction mapping: High-resolution characterization of microbial community shifts and their functional consequences
Economic validation: Large-scale production trials assessing cost-effectiveness compared to AGPs and disease management costs
Conclusions
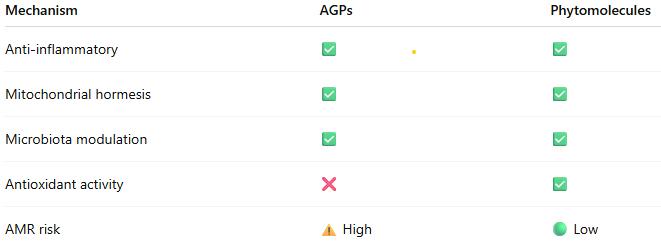
The scientific evidence demonstrates that standardized phytomolecules operate through well-characterized biological mechanisms that substantially replicate those of AGPs:
Anti-inflammatory effects reducing energetic costs of immune activation
Mitochondrial hormesis enhancing energy metabolism and cellular resilience
Selective microbiota modulation supporting beneficial bacteria while controlling pathogens
Intestinal barrier enhancement improving nutrient absorption and reducing translocation
Antioxidant activity mitigating oxidative stress and supporting immune function
When properly standardized and formulated for controlled release, phytomolecules deliver growth promotion, feed efficiency improvements, and disease resistance comparable to AGPs, while potentially offering advantages in AMR risk profile, stress resilience, and consumer acceptance.
The mechanistic convergence between AGPs and phytomolecules, coupled with demonstrated efficacy in controlled trials, provides producers with confidence that science-based phytomolecular interventions represent legitimate alternatives to AGPs. Success depends on product standardization, appropriate dosing, and understanding that phytomolecules work through fundamental biological pathways rather than undefined or mystical mechanisms.
As the livestock industry continues to navigate the post-AGP era, standardized phytomolecules offer a scientifically sound, mechanistically validated approach to maintaining animal performance, health, and welfare while addressing antimicrobial resistance concerns.
References
Adaszek, Ł., et al. “Properties of Capsaicin and Its Utility in Veterinary and Human Medicine.” Research in Veterinary Science, vol. 123, 2019, pp. 14 – 19.
Bottje, W., et al. “Mitochondrial proton leak kinetics and relationship with feed efficiency within a single genetic line of male broilers”. Poultry Science, Volume 88, Issue 8, 1 August 2009, p. 1683-1693.
Bortoluzzi, C., et al. “A Protected Complex of Biofactors and Antioxidants Improved Growth Performance and Modulated the Immunometabolic Phenotype of Broiler Chickens Undergoing Early Life Stress.” Poultry Science, vol. 100, 2021, p. 101176.
Bourgin, M., et al. “Bile Salt Hydrolases: At the Crossroads of Microbiota and Human Health.” Microorganisms, vol. 9, no. 1122, 2021.
Bravo, D., et al. “A Mixture of Carvacrol, Cinnamaldehyde, and Capsicum Oleoresin Improves Energy Utilization and Growth Performance of Broiler Chickens Fed Maize-Based Diet.” Journal of Animal Science, vol. 92, 2014, pp. 1531 – 1536.
Bringas-Lantigua, M., et al. “Influence of Spray-Dryer Air Temperatures on Encapsulated Mandarin Oil.” Drying Technology, vol. 29, 2011, pp. 520–526.
Burtscher, J., et al. “Mitochondrial Stress and Mitokines in Aging.” Aging Cell, vol. 22, no. 2, 2023, e13770.
El-Hack, M. et al. “Integrating metabolomics for precision nutrition in poultry: optimizing growth, feed efficiency, and health”. Frontiers in Veterinary Science, Sec. Animal Nutrition and Metabolism, Volume 12 – 2025. https://doi.org/10.3389/fvets.2025.1594749
Elolimy, Ahmed A., et al. “Effects of Microencapsulated Essential Oils and Seaweed Meal on Growth Performance, Digestive Enzymes, Intestinal Morphology, Liver Functions, and Plasma Biomarkers in Broiler Chickens.” Journal of Animal Science, vol. 103, 2025, p. skaf092, https://doi.org/10.1093/jas/skaf092.
Fernández Miyakawa, Mariano E., et al. “How Did Antibiotic Growth Promoters Increase Growth and Feed Efficiency in Poultry?” Poultry Science, vol. 103, no. 2, 2024, article 103136. https://doi.org/10.1016/j.psj.2023.103136
Gaskins, H. Rex, C. T. Collier, and D. B. Anderson. “Antibiotics as Growth Promotants: Mode of Action.” Animal Biotechnology, vol. 13, no. 1, 2002, pp. 29 – 42.
Gharsallaoui, A., et al. “Applications of Spray-Drying in Microencapsulation of Food Ingredients: An Overview.” Food Research International, vol. 40, no. 9, 2007, pp. 1107-21.
Gutiérrez-Chávez, Vanesa, et al. “Capsaicinoids and Capsinoids of Chilli Pepper as Feed Additives in Livestock Production: Current and Future Trends.” Animal Nutrition, vol. 22, 2025, pp. 483 – 501. https://doi.org/10.1016/j.aninu.2025.03.014.
Gupta, A., et al. “Capsaicin and Capsinoids: Recent Updates on Their Health Benefits and Mechanisms of Action.” Phytotherapy Research, vol. 36, no. 5, 2022, pp. 1898 – 1912.
Hu, Q., Li, X., Chen, F., Wan, R., Yu, C.-W., Li, J., McClements, D. J., & Deng, Z. (2020). “Microencapsulation of an essential oil (cinnamon oil) by spray drying: Effects of wall materials and storage conditions on microcapsule properties“. Journal of Food Processing and Preservation, 44(11). https://doi.org/10.1111/jfpp.14805
Khameneh, B., et al. “Mechanisms of Antibiotic Resistance Resensitization by Phytochemicals: Review.” Phytomedicine, vol. 85, 2021, p. 153529.
Kim, D. K., et al. “Effects of Capsicum and Curcuma on Necrotic Enteritis in Broilers.” Poultry Science, vol. 94, 2015, pp. 2314 – 2321.
Kim, J. S., et al. “Anti-inflammatory Effects of Plant-Derived Molecules via NF-κB and MAPK Pathways.” International Immunopharmacology, vol. 10, no. 3, 2010, pp. 306 – 314.
Lee, S. H., et al. “Allium Hookeri Extract Enhances Tight Junction Proteins in Broilers.” Journal of Animal Physiology and Animal Nutrition, vol. 101, no. 1, 2017, pp. e48 – e56.
Li, X., et al. “Capsicum Oleoresin Supplementation Improves Digestive Enzyme Activity and Gut Morphology in Broilers.” Poultry Science, vol. 101, no. 7, 2022, p. 101844.
Lin, J. “Effect of Antibiotics on the Intestinal Microbiota and Their Role in Animal Growth.” Animal Biotechnology, vol. 25, no. 3, 2014, pp. 149 – 157.
Lillehoj, H., et al. “Phytochemicals as Antibiotic Alternatives to Promote Growth and Enhance Host Health.” Veterinary Research, vol. 49, no. 76, 2018.
Liu, Y., et al. “Dietary Capsicum Extract Enhances Intestinal Barrier Function and Growth in Pigs.” Journal of Animal Science, vol. 91, 2013, pp. 518 – 525.
Long, L., et al. “Phytogenic Feed Additives Modulate Intestinal Immunity and Antioxidant Status in Pigs and Poultry.” Frontiers in Veterinary Science, vol. 8, 2021, p. 620998.
Muurinen, J., et al. “Mushroom Powder Supplementation Increases Antibiotic Resistance Gene Mobility in Pig Feces.” Frontiers in Microbiology, vol. 12, 2021, p. 676678.
Niewold, T. A. “The Non-antibiotic Anti-inflammatory Effect of Antimicrobial Growth Promoters, the Real Mode of Action? A Hypothesis.” Poultry Science, vol. 86, 2007, pp. 605 – 609.
Perry, F., C. N. Johnson, L. Lahaye, E. Santin, D. R. Korver, M. H. Kogut, and R. J. Arsenault. “Protected Biofactors and Antioxidants Reduce the Negative Consequences of Virus and Cold Challenge by Modulating Immunometabolism via Changes in the Interleukin-6 Receptor Signaling Cascade in the Liver.” Poultry Science, vol. 103, no. 9, 2024, article 104044. https://doi.org/10.1016/j.psj.2024.104044
Rahman, Md, et al. “Insights in the Development and Uses of Alternatives to Antibiotic Growth Promoters in Poultry and Swine Production.” Antibiotics, vol. 11, no. 6, 2022, p. 766, https://doi.org/10.3390/antibiotics11060766.
Reda, F. M., et al. “Capsicum Extract Supplementation Modulates Gut Microbiota and Performance in Japanese Quails.” Animal Feed Science and Technology, vol. 265, 2020, p. 114507.
Rosca, I., et al. “Capsaicin Induces Osmotic Stress in Gram-negative Pathogens.” Veterinary Sciences, vol. 7, no. 4, 2020, p. 172.
Sahin, K., et al. “Dietary Capsicum Extract Reduces Oxidative Stress in Heat-stressed Japanese Quails.” Poultry Science, vol. 95, no. 2, 2016, pp. 231 – 240.
Saleh, A. A., et al. “Herbal Extract Mixtures Improve Antioxidant Status and Performance in Broilers.” Poultry Science, vol. 97, no. 11, 2018, pp. 3927 – 3936.
Stevanović, Z. D., et al. „Essential oils as feed additives—Future perspectives”. Molecules, 23(7), 2018, pp1717.
Suganya, R., et al. “Phytochemicals in Combination with Antibiotics: Antimicrobial Resistance Breakers.” Antibiotics, vol. 11, 2022, p. 123.
Zhang, Benyuan et al. “Mitochondrial Stress and Mitokines: Therapeutic Perspectives for the Treatment of Metabolic Diseases.” Diabetes & Metabolism Journal vol. 48,1, 2024, pp. 1-18.
Zhan, Ru, et al. “Effects of Antibiotics on Chicken Gut Microbiota: Community Alterations and Pathogen Identification.” Frontiers in Microbiology, vol. 16, 2025, article 1562510. https://doi.org/10.3389/fmicb.2025.1562510
Zhang, Y., et al. “Effects of Vanillin, Thymol, and Eugenol on Glucose and Lipid Metabolism via TRPV1 Activation.” Journal of Agricultural and Food Chemistry, vol. 65, no. 13, 2017, pp. 2719 – 2727.
Phytomolecules: Sustainability and Efficiency in Pig Production
Conference Report
By M. Rosenthal, Global Application Manager Swine, EW Nutrition GmbH
Sustainability is essential for the long-term survival of our planet. In pig production, sustainability involves maintaining economically viable outputs while simultaneously safeguarding animal health and welfare and minimizing environmental impact. The goal is to produce pork that is profitable, ethical, and has a minimal ecological footprint.
Phytomolecules, the bioactive constituents of plant-derived essential oils, play a promising role in advancing this goal. With multifunctional gut health benefits including antimicrobial, anti-inflammatory, antioxidant, and digestive-supportive properties, phytomolecules help maintain gut health and reduce the need for antibiotics. By improving feed efficiency, enhancing resilience, and supporting intestinal integrity, phytomolecules contribute to both sustainability and efficiency in pig production systems.
Targeting sustainability in pig production
Achieving sustainability in pig production requires a balanced approach that considers three key perspectives: those of the producer, the pig, and the environment.
For the producer, sustainable pig production must be profitable to ensure the long-term viability of the industry. This includes factors such as efficient feed conversion, optimized production practices, and fair market prices.
Another aspect is the maintenance of animal health and well-being, which is essential for optimal pig performance and can be achieved by providing appropriate housing, nutrition, and veterinary care, as well as minimizing stress and disease.
From an environmental perspective, minimizing negative impacts, such as greenhouse gas emissions, water pollution, and land degradation, is a key objective. Various strategies, such as improved manure management, efficient nutrient utilization, reuse of farm resources like manure and water, and the use of by-products from other industries as feed ingredients, can be applied.
Strategy for efficient pig production
Historically, pig production has relied heavily on the use of antibiotics to control enteric pathogens, promote gut health, and enhance growth. While effective in the short term, this practice led to unintended consequences, including the emergence of antimicrobial resistance (amr), disruption of microbiota across multiple organ systems, difficulties in manure management, and environmental contamination.
These outcomes triggered societal concern, regulatory interventions, and economic pressure, prompting a shift away from routine antibiotic use. The industry now faces increasing expectations for environmentally responsible practices, reduced dependence on antibiotics, and cost-effective, sustainable solutions.
Achieving both efficiency and sustainability in pig production requires a holistic, system-wide approach that includes an innovative, solution-oriented mindset, optimized management practices, and the adoption of effective gut health antibiotic alternatives.
The foundation of efficiency – the gut
The pigs gastrointestinal tract is the largest and most vulnerable interface between the pig and its external environment. It is a highly organized ecosystem comprised of epithelial cells, the mucosal immune system, and a diverse microbiome consisting of both beneficial commensal microbes and potentially harmful pathogens.
The functions of the gut include nutrient absorption, chemosensing of nutrients and other compounds, immune defence, and balancing the highly diverse microbiome within this complex environment (Furness et al. , 2013). Disruption of this ecosystems homeostasis can impair not only gut function and health but also negatively affect the overall well-being and growth efficiency of the pig.
When evaluating antibiotic alternatives to support this ecosystems homeostasis in the face of challenges, considerations include safety for humans, animals, and the environment, cost-effectiveness, antimicrobial efficacy, the ability to increase nutrient availability, and to modulate immune activation and inflammation.
Functional feed additives commonly utilized in pig nutrition, alone or in combination, include organic acids, probiotics, immunoglobulins, medium-chain fatty acids, and phytomolecules.
Phytomolecules: supporting gut health and performance
Phytomolecules are the bioactive components of plant-derived essential oils. Due to the variability in phytomolecule content and the presence of volatile and astringent components in essential oil extracts, utilizing commercial phytomolecule products is recommended. Proprietary formulations utilize encapsulation or matrix technology to protect the phytomolecules from damage or loss during storage, processing, and passage through the stomach.
Extensive research in humans and animals has identified phytomolecules as having antimicrobial, anti-inflammatory, antioxidative, and coccidiostatic properties. They enhance digestibility and immunity, promote gut health through differential modulation of bacterial populations, and reduce inflammation and oxidative stress (Brenes et al., 2010; Puvaca et al. , 2013; Chitprasert et al., 2014). Phytomolecules most researched and utilized in pig feed additives to date include terpenes (e. G., carvacrol and thymol) and phenylpropenes (e.g., cinnamaldehyde and eugenol).
1. Direct antimicrobial activity of phytomolecules
Phytomolecules such as carvacrol and thymol provide broad-spectrum antimicrobial activities against Gram- and Gram+ bacteria, fungi, and yeast and are regarded as promising alternatives to antibiotics in swine production systems (Lambert et al., 2001; Delaquis et al., 2002; Abbaszadeh et al., 2014).
Phytomolecules directly target bacterial cells through multiple mechanisms, with the cell wall and membrane being major sites of action. The lipophilic structure of phytomolecules enables their entry through bacterial membranes among the fatty acid chains, causing the cell wall and membranes to expand and become more fluid. This damage collapses the cell wall and cytoplasmic membrane, resulting in the destruction of membrane proteins, the coagulation of the cytoplasm, and a reduction in proton motive force. The result is leakage of vital intracellular contents and death of the bacterial cell (Cox et al., 1998; Faleiro, 2011; Nazzaro et al., 2013; Yap et al., 2014). For example, thymol and carvacrol can damage the outer membrane of Salmonellatyphimurium and Escherichia coli o157: h7 (Helander et al., 1998).
A further direct antimicrobial action involves phytomolecules acting as trans-membrane carriers, exchanging a hydroxyl proton for a potassium ion, resulting in dissipation of the ph gradient and electrical potential over the bacterial cytoplasmic membrane. The result is a reduced proton motive force and the depletion of the intracellular adenosine triphosphate (APT) pools. Loss of potassium further inhibits bacterial function as it is needed for the activation of cytoplasmic enzymes to maintain osmotic pressure and regulate intracellular pH. (Wendakoon et al., 1995).
In summary, the primary direct antimicrobial mechanism of action for terpene and phenylpropene phytomolecules is related to their effects on cell walls and cytoplasmic membranes, and energy metabolism of pathogenic bacteria.
2. Indirect antimicrobial activity of phytomolecules
Phytomolecules indirectly impact the physiological functioning and virulence capability of pathogenic bacteria through the interference of quorum-sensing (QS). QS involves pathogenic bacteria producing signaling molecules that are released based on cell numbers. The detection of these molecules regulates pathogen population behavior such as attachment, biofilm formation, and motility, i. e. , virulence (Greenberg, 2003; Joshi et al., 2016).
QS mechanisms require signal synthesis, signal accumulation, and signal detection, providing three opportunities for QS inhibitors to disrupt pathogenic bacteria from causing disease (Czajkowski and Jafra, 2009; Lasarre and Federle, 2013). Eugenol and carvacrol have been extensively studied for their QS inhibition activities (Zhou et al., 2013; Burt et al., 2014).
3. Combinations increase efficacy
Additional antimicrobial effects can be seen when different phytomolecules are combined, and/or applied with other functional additives such as organic acids (Souza et al., 2009; Hulankova and Borilova, 2011). Zhou et al. (2007) reported that carvacrol or thymol in combination with acetic or citric acid had a better efficacy against S. typhimurium when compared to the individual phytomolecule or organic acid. In recent studies, results have shown in vivo efficacy of such synergistic dietary strategies in pigs (Diao et al., 2015; Balasubramanian et al., 2016). The combined inclusion of phytomolecules and organic acids in pig diets before slaughter may hinder Salmonella shedding and seroprevalence (Walia et al., 2017; Noirrit et al., 2016).
4. Phytomolecules are more than antimicrobials
In addition to acting as antimicrobials, phytomolecules enhance production efficiency through multiple complementary mechanisms, including direct anti-inflammatory, antioxidative, digestive, and gut barrier-supportive effects.
Anti-inflammatory effects: Gut inflammation in pigs not only compromises intestinal function and barrier integrity but also has a direct negative impact on growth performance and overall health. Chronic or excessive immune activation diverts energy away from productive processes such as growth and feed efficiency.
Phytomolecules have demonstrated the ability to modulate immune responses by influencing key cell-signalling pathways involved in inflammation. For example, compounds such as cinnamaldehyde and carvacrol can modulate the activity of critical transcription factors, including nuclear factor erythroid 2 2-related factor 2 (Nrf2) and nuclear factor kappa B (NF-κB). Through this dual action, phytomolecules can simultaneously activate antioxidant defences and suppress pro-inflammatory signalling, thereby reducing intestinal inflammation and supporting improved performance outcomes (Krois-mayr et al., 2008; Wondrak et al., 2010; Zou et al., 2016).
Antioxidant effects: oxidative stress is a major biological challenge in modern swine production systems, where high-performance animals are frequently exposed to stressors such as weaning, disease challenges, heat stress, mycotoxin exposure, transport, and overcrowding. These stressors promote the generation of reactive oxygen species (ROS), and when ROS production exceeds the capacity of the pig’s antioxidant defence systems, oxidative stress occurs.
This imbalance can negatively affect growth, immunity, muscle integrity, feed intake, milk yield, and reproductive performance, including increased abortion rates in gestating sows (Zhou et al., 2013; Burt et al., 2014). As a result, there is growing interest in the use of natural antioxidant compounds, particularly phytomolecules, to counteract these detrimental effects. For example, carvacrol and thymol (1:1 ratio) at 100 mg/kg dietary supplementation reduced weaning-associated oxidative stress by decreasing TNF-α mRNA expression in the intestinal mucosa (Wei et al., 2017).
Additionally, carvacrol supplementation in the diets of late gestation and lactating sows under oxidative stress conditions significantly improved piglet performance (Tan et al., 2015).
Digestive function: The gastrointestinal tract functions not only as a site for nutrient absorption but also as a sensory organ. Specialized chemosensors in the gut monitor the concentration and composition of nutrients, playing a crucial role in the regulation of digestive enzyme secretion, gut peptide release, feed intake, and nutrient absorption and metabolism.
Studies in weaner piglets have shown that certain phytomolecules can stimulate the secretion of digestive enzymes and enhance gastrointestinal function (Maenner et al., 2011; Li et al., 2012).
Tight junctions and gut barrier integrity: The intestinal epithelium functions as a highly dynamic and selective barrier, facilitating the absorption of fluids and solutes while preventing the translocation of pathogens and toxins into underlying tissues. This barrier function occurs through intercellular tight junctions. During episodes of mucosal inflammation, the integrity of these junctions can be compromised, leading to increased intestinal permeability, reduced nutrient absorption, and systemic immune activation and inflammation.
Research has shown that phytomolecules can enhance transepithelial electrical resistance and upregulate the expression of tight junction proteins, reducing epithelial permeability and maintaining a functional barrier, even under inflammatory conditions (Yu et al., 2020; Kim and Kim, 2019).
Sustainable efficiency in pig production supported by in-feed phytomolecules
As the pig industry moves away from reliance on in-feed antibiotics, the need for sustainable, efficient, and health-focused production strategies has never been greater. Modern pig production systems must respond to societal expectations, regulatory mandates, and environmental pressures, while still maintaining profitability and high animal welfare standards.
Central to this transformation is a holistic approach-one that includes a shift in mindset among stakeholders, optimized management across all production domains, and the strategic use of effective antibiotic alternatives. The gastrointestinal tract, as the core of nutrient absorption and immune defence, is a critical control point for supporting health and performance.
Phytomolecules and other functional feed additives have demonstrated potential to enhance gut integrity, reduce inflammation, combat oxidative stress, and improve nutrient utilization. While no single solution can fully replace antibiotics, targeted combinations of these compounds have shown the most consistent success in promoting gut health and sustainable performance.
With continued innovation, collaboration, and science-based application of these alternatives, the industry is well-positioned to achieve its goals of profitable, ethical, and ecologically responsible pork production for the future.
References
Abbaszadeh, S., A. Sharifzadeh, H. Shokri, A. Khosravi, and A. Abbaszadeh. 2014. “Antifungal Efficacy of Thymol, Carvacrol, Eugenol and Menthol as Alternative Agents to Control the Growth of Food-Relevant Fungi.” Journal de Mycologie Médicale 24 (2): 51–56. Balasubramanian, B., J. W. Park, and I. H. Kim. 2016. “Evaluation of the Effectiveness of Supplementing Micro-Encapsulated Organic Acids and Essential Oils in Diets for Sows and Suckling Piglets.” Italian Journal of Animal Science 15 (4): 626–33. Baschieri, A., M. D. Ajvazi, J. L. F. Tonfack, L. Valgimigli, and R. Amorati. 2017. “Explaining the Antioxidant Activity of Some Common Non-Phenolic Components of Essential Oils.” Food Chemistry 232: 656–63. Berchieri-Ronchi, C., S. Kim, Y. Zhao, C. Correa, K.-J. Yeum, and A. Ferreira. 2011. “Oxidative Stress Status of Highly Prolific Sows During Gestation and Lactation.” Animal 5 (11): 1774–79. Brenes, A., and E. Roura. 2010. “Essential Oils in Poultry Nutrition: Main Effects and Modes of Action.” Animal Feed Science and Technology 158 (1): 1–14. Burt, S. A., V. T. Ojo-Fakunle, J. Woertman, and E. J. Veldhuizen. 2014. “The Natural Antimicrobial Carvacrol Inhibits Quorum Sensing in Chromobacterium violaceum and Reduces Bacterial Biofilm Formation at Sub-Lethal Concentrations.” PLoS One 9 (4): e93414. Chitprasert, P., and P. Sutaphanit. 2014. “Holy Basil (Ocimum sanctum Linn.) Essential Oil Delivery to Swine Gastrointestinal Tract Using Gelatine Microcapsules Coated with Aluminium Carboxymethyl Cellulose and Beeswax.” Journal of Agricultural and Food Chemistry 62 (52): 12641–48. Cox, S., J. Gustafson, C. Mann, J. Markham, Y. Liew, and R. Hartland, et al. 1998. “Tea Tree Oil Causes K⁺ Leakage and Inhibits Respiration in Escherichia coli.” Letters in Applied Microbiology 26 (5): 355–58. Czajkowski, R., and S. Jafra. 2009. “Quenching of Acyl-Homoserine Lactone-Dependent Quorum Sensing by Enzymatic Disruption of Signal Molecules.” Acta Biochimica Polonica 56 (1): 1–16. Delaquis, P. J., K. Stanich, B. Girard, and G. Mazza. 2002. “Antimicrobial Activity of Individual and Mixed Fractions of Dill, Cilantro, Coriander and Eucalyptus Essential Oils.” International Journal of Food Microbiology 74 (1): 101–9. Diao, H., P. Zheng, B. Yu, J. He, X. Mao, J. Yu, et al. 2015. “Effects of Benzoic Acid and Thymol on Growth Performance and Gut Characteristics of Weaned Piglets.” Asian-Australasian Journal of Animal Sciences 28 (6): 827–35. Faleiro, M. 2011. “The Mode of Antibacterial Action of Essential Oils.” In Science Against Microbial Pathogens: Communicating Current Research and Technological Advances, vol. 2, 1143–56. Badajoz, Spain: Formatex Research Center. Furness, J., L. Rivera, and H. J. Cho, et al. 2013. “The Gut as a Sensory Organ.” Nature Reviews Gastroenterology & Hepatology 10: 729–40. Greenberg, E. P. 2003. “Bacterial Communication and Group Behavior.” Journal of Clinical Investigation 112 (9): 1288–90. Helander, I. M., H.-L. Alakomi, K. Latva-Kala, T. Mattila-Sandholm, I. Pol, E. J. Smid, et al. 1998. “Characterization of the Action of Selected Essential Oil Components on Gram-Negative Bacteria.” Journal of Agricultural and Food Chemistry 46 (9): 3590–95. Hulankova, R., and G. Borilova. 2011. “In Vitro Combined Effect of Oregano Essential Oil and Caprylic Acid Against Salmonella Serovars, Escherichia coli O157:H7, Staphylococcus aureus and Listeria monocytogenes.” Acta Veterinaria Brno 80 (4): 343–48. Joshi, J. R., N. Khazanov, H. Senderowitz, S. Burdman, A. Lipsky, and I. Yedidia. 2016. “Plant Phenolic Volatiles Inhibit Quorum Sensing in Pectobacteria and Reduce Their Virulence by Potential Binding to ExpI and ExpR Proteins.” Scientific Reports 6: 38126. Kim, M. S., and J. Y. Kim. 2019. “Cinnamon Subcritical Water Extract Attenuates Intestinal Inflammation and Enhances Intestinal Tight Junction in a Caco-2 and RAW264.8 Co-Culture Model.” Food & Function 20: 4350–60. Kroismayr, A., J. Sehm, M. Pfaffl, K. Schedle, C. Plitzner, and W. Windisch.2008. “Effects of Avilamycin and Essential Oils on mRNA Expression of Apoptotic and Inflammatory Markers and Gut Morphology of Piglets.” Czech Journal of Animal Science 53: 377–87. Lambert, R., P. N. Skandamis, P. J. Coote, and G. J. Nychas. 2001. “A Study of the Minimum Inhibitory Concentration and Mode of Action of Oregano Essential Oil, Thymol and Carvacrol.” Journal of Applied Microbiology 91 (3): 453–62. LaSarre, B., and M. J. Federle. 2013. “Exploiting Quorum Sensing to Confuse Bacterial Pathogens.” Microbiology and Molecular Biology Reviews 77 (1): 73–111. Li, P., X. Piao, Y. Ru, X. Han, L. Xue, and H. Zhang. 2012. “Effects of Adding Essential Oil to the Diet of Weaned Pigs on Performance, Nutrient Utilization, Immune Response and Intestinal Health.” Asian-Australasian Journal of Animal Sciences 25 (11): 1617–26. Maenner, K., W. Vahjen, and O. Simon. 2011. “Studies on the Effects of Essential-Oil-Based Feed Additives on Performance, Ileal Nutrient Digestibility, and Selected Bacterial Groups in the Gastrointestinal Tract of Piglets.” Journal of Animal Science 89 (7): 2106–12. Nazzaro, F., F. Fratianni, L. De Martino, R. Coppola, and V. De Feo. 2013. “Effect of Essential Oils on Pathogenic Bacteria.” Pharmaceuticals 6 (12): 1451–74. Noirrit, M., and F. Philippe. 2016. “Reduction of Salmonella Prevalence on Sows and Finishing Pigs by Use of a Protected Mix of Organic Acids and Essential Oils in the Feed of Lactating Sows and Weaned Piglets.” Journées Recherche Porcine 48: 351–52. Puvaca, N., V. Stanacev, D. Glamocic, J. Levic, L. Peric, and D. Milic. 2013. “Beneficial Effects of Phytoadditives in Broiler Nutrition.” World’s Poultry Science Journal 69 (1): 27–34. Souza, E. L., J. C. Barros, M. L. Conceiçao, N. J. Gomes Neto, and A. C. V. Costa.2009. “Combined Application of Origanum vulgare L. Essential Oil and Acetic Acid for Controlling the Growth of Staphylococcus aureus in Foods.” Brazilian Journal of Microbiology 40 (2): 387–93. Tan, C., H. Wei, H. Sun, J. Ao, G. Long, S. Jiang, et al. 2015. “Effects of Dietary Supplementation of Oregano Essential Oil to Sows on Oxidative Stress Status, Lactation Feed Intake of Sows, and Piglet Performance.” BioMed Research International 2015: Article ID 941754. Walia, K., H. Argüello, H. Lynch, F. C. Leonard, J. Grant, D. Yearsley, et al. 2017. “Effect of Strategic Administration of an Encapsulated Blend of Formic Acid, Citric Acid, and Essential Oils on Salmonella Carriage, Seroprevalence, and Growth of Finishing Pigs.” Preventive Veterinary Medicine 137: 28–35. Wei, H. K., H. X. Xue, Z. Zhou, and J. Peng. 2017. “A Carvacrol-Thymol Blend Decreased Intestinal Oxidative Stress and Influenced Selected Microbes Without Changing the Messenger RNA Levels of Tight Junction Proteins in Jejunal Mucosa of Weaning Piglets.” Animal 11 (2): 193–201. Wendakoon, C. N., and M. Sakaguchi. 1995. “Inhibition of Amino Acid Decarboxylase Activity of Enterobacter aerogenes by Active Components in Spices.” Journal of Food Protection 58 (3): 280–83. Wondrak, G. T., N. F. Villeneuve, S. D. Lamore, A. S. Bause, T. Jiang, and D. D. Zhang. 2010. “The Cinnamon-Derived Dietary Factor Cinnamic Aldehyde Activates the Nrf2-Dependent Antioxidant Response in Human Epithelial Colon Cells.” Molecules 15 (5): 3338–55. Yap, P. S. X., B. C. Yiap, H. C. Ping, and S. H. E. Lim. 2014. “Essential Oils, a New Horizon in Combating Bacterial Antibiotic Resistance.” Open Microbiology Journal 8 (1). Yu, J., Y. Song, B. Yu, J. He, P. Zheng, X. Mao, Z. Huang, Y. Luo, J. Luo, H. Yan, Q. Wang, H. Wang, and D. Chen. 2020. “Tannic Acid Prevents Post-Weaning Diarrhea by Improving Intestinal Barrier Integrity and Function in Weaned Piglets.” Journal of Animal Science and Biotechnology 11: 87. Zhou, F., B. Ji, H. Zhang, H. Jiang, Z. Yang, J. Li, et al. 2007. “Synergistic Effect of Thymol and Carvacrol Combined with Chelators and Organic Acids Against Salmonella Typhimurium.” Journal of Food Protection 70 (7): 1704–9. Zhou, L., H. Zheng, Y. Tang, W. Yu, and Q. Gong. 2013. “Eugenol Inhibits Quorum Sensing at Subinhibitory Concentrations.” Biotechnology Letters 35 (4): 631–37. Zou, Y., J. Wang, J. Peng, and H. Wei. 2016. “Oregano Essential Oil Induces SOD1 and GSH Expression Through Nrf2 Activation and Alleviates Hydrogen Peroxide-Induced Oxidative Damage in IPEC-J2 Cells.” Oxidative Medicine and Cellular Longevity 2016: Article ID 5987183.
The Gut: A Main Component of Poultry’s Immune System
By Dr. Inge Heinzl, Editor EW Nutrition
Gut health is a critical factor in poultry production, influencing growth performance, feed efficiency, and overall bird health. A well-functioning digestive system ensures optimal nutrient absorption and ultimately contributes to economic sustainability in poultry farming.
However, another essential function of the gut is its significant role in immune defense, as evidenced by the fact that 80% of all active immune cells are in the gut. It is essential for the organism to keep a sensitive balance by eliminating invading pathogens while maintaining self-tolerance to avoid autoimmunity. Being 1.5 to 2.3 m long and with a big contact area to the external environment, the gut is the first line of defense when pathogens have orally entered the organism. For this purpose, the intestine has several specialized cells and a plethora of diverse microorganisms – the microbiome.
A balanced gut environment, therefore, enhances resistance to diseases, helps prevent infections, and reduces the need for antibiotics.
Which tools are available in the gut to counteract pathogenic attacks?
The gut wall, per se, has several fixed tools to fight pathogenic offenses, such as the mucus layers and the epithelium with highly specialized cells. Figure 1 shows in detail the different parts of the gut immune system.
Figure 1: Structure of the intestinal wall with its specialized cells (Kong et al., 2018)
1. Mucus layers
The mucus layers form the first host-derived line of defense. They help trap invasive bacteria and facilitate their removal via luminal flow. The protective properties may depend on whether the mucin is neutral or acidic, sialylated or sulfated (Broom and Kogut, 2018). The glycoprotein mucins forming the mucus layer (mainly MUC2 in the small and large intestine and MUC5ac in the proventriculus) are produced by the goblet cells, part of the intestinal epithelium just beneath.
2. Intestinal epithelium
The one-layered intestinal epithelium represents a physical barrier and consists of normal enterocytes, as well as specialized cells. All the cells are closely linked by tight junctions, consisting of claudin, occludin, and junctional adhesion molecules (JAM).
The following diverse specialized cells protect the organism from pathogenic attacks:
2.1 Proliferating stem cells
These cells are ready to replace damaged epithelial cells in the case of inflammation.
2.2 Paneth cells
Paneth cells are situated at the bottom of the Lieberkühn crypts, neighboring the stem cells in the jejunum and the ileum. Paneth cells have different tasks:
In normal conditions, they maintain homeostasis by regulating the microbiome’s composition via the secretion of antimicrobial peptides, which are accumulated in apically oriented secretory granules, performing phagocytosis and efferocytosis. Additionally, the Paneth cells provide niche factors for the intestinal stem cell compartment, absorb heavy metals, and preserve the integrity of the intestinal barrier. If one or more of these functions are impaired, intestinal and systemic inflammations or infections can develop (Wallaeys et al., 2022). The number of Paneth cells and their diameter can be enhanced via feeding. Agarwal et al. (2022) noticed a significant increase in the number and diameter of Paneth cells after feeding quinoa soluble fiber and/or quercetin 3-glucoside.
2.3 M cells
M cells (M coming from microfold and indicating the structure) are specialized epithelial cells localized along the antimesenteric border in the epithelium of the ileum. They are crucial for the immune system and an essential part of the gut-associated lymphoid tissue (GALT), a sub-system of the mucosa-associated lymphoid tissue (MALT).
M cells play an important role in the function of the immune system. They act as a transport system for antigens. They sample antigens (macromolecules, bacteria, viruses, small parasites) via the apical membrane. After the phagocytosis of the foreign organism/substance, the antigen gets through the cell and is consigned to cells of the adaptive immune system (e.g., the B-cells) at the basal side. The exact transport and the handover to the cells of the adaptive immune system are still unclear. It is also not clarified whether the antigens are processed inside the cells.
2.4 Dendritic cells
Dendritic cells are a kind of leucocyte derived from the bone marrow. Immature dendritic cells have a star-like shape. They are specialized to identify, uptake, transport, process, and present antigens to other immune system cells on their surface. To identify and uptake harmful substances/microbes, they carry receptors on their surface that recognize the attributes often occurring in pathogenic viruses, bacteria, and fungi. After contact with the antigen, the cell moves to secondary lymphoid tissue, and in the intestine, this is predominantly Mucosa-Associated Lymphoid Tissue (MALT). Arriving as mature and not phagocytizing dendritic cells, they present the antigens of the pathogens to the T-lymphocytes. For this purpose, they use cell surface proteins (MHC proteins). This presentation, together with co-stimulators and cytokines, activates naïve T-lymphocytes to develop into the relevant T-cell (fighting viruses, bacteria…) and proliferate, leading to the clearance of the pathogen.
On the other hand, dendritic cells can also suppress an immune reaction if the “suspicious subjects” are harmless or belong to the organism. Dendritic cells are the most potent antigen-presenting cells of the immune system.
2.5 Goblet cells
Goblet cells originate from pluripotent stem cells and are located between the enterocytes in the inner mucus layer of the intestine. Goblet cells develop and mature rapidly after hatching due to external stimuli such as environmental and dietary factors, but also intestinal microbiota (Duangnumsawang et al., 2021). They derive their name from their goblet-like appearance. The basal site is thin, but the cell gets thicker toward the apical side. In the thicker cell organisms, vesicles with mucins are stored and explosively released to the surface by exocytosis.
Figure 2: Goblet cells
The mucins (MUC2) are viscous, slime-forming substances consisting of a protein string bound to many sugar chains. Due to their oligosaccharide chain structure, they offer adhesion binding sites for intestinal commensal bacteria and enhance probiotic colonization (Liu et al, 2020). They have a high water-binding capacity, which is responsible for their slimy and protective characteristics. In the case of inflammation, mucin production can increase strongly.
By providing bicarbonate for proper mucin unfolding in the small intestine, goblet cells help maintain homeostasis and the intestinal barrier function. Furthermore, goblet cells can form goblet cell-associated passages (GAPs) and deliver luminal substances to the antigen-presenting cells in the underlying lamina propria that can start an adaptive immune response (Knoop and Newberry, 2018).
As with Paneth cells, the number of goblet cells also increases by feeding quinoa soluble fibers.
2.6 Neuroendocrine cells
Enterochromaffin cells are neuroendocrine cells found in the epithelium of the whole digestive tract, mainly in the small intestine, the colon, and the ceca. They belong to the enteric endocrine system, are part of the diffuse neuroendocrine system, and produce 95% of the serotonin in the organism. Enterochromaffin cells act as chemo- and mechanosensors. They react to free fatty acids, amino acids, and other chemicals as well as physical forces occurring during peristaltic activity in the gut, thus modulating the secretion of water and electrolytes as well as gut motility and visceral sensation of pain (Linan-Rico et al., 2016; Diwakarla et al., 2018).
Serotonin, on its side, has been shown to affect the composition of the gut microbiota (Kwon et al., 2019) and to modulate bacterial physiology (Knecht et al., 2016). Gut-derived serotonin is responsible for immune responses (Baganz and Blakely, 2012) but also for the regulation of other functions such as bone development (Chabbi-Achengli et al., 2012), gut motility, and platelet aggregation (Berger et al., 2009). A deficient serotonergic system can cause psychopathological behaviors such as feather pecking.
3. Last but not least – the microbiome
The poultry gut microbiome consists of bacteria, fungi, protozoa, and viruses. Beneficial microbes, such as Lactobacillus, Bifidobacterium, and Bacteroides, contribute to gut health and immunity.
On the one hand, microbes are involved in digestion and nutrient synthesis. They assist in breaking down fiber, producing short-chain fatty acids, and synthesizing essential vitamins. On the other hand, they contribute to immune defense:
Beneficial bacteria (BB) prevent the colonization of harmful microbes: The bacteria inhabiting the poultry gut act against pathogens by competing with them for nutrients and binding sites at the intestinal mucosa.
Beneficial bacteria prevent/reduce inflammation and stabilize the intestinal mucosa Abaidullah et al. (2019) showed in their review how beneficial bacteria influence the immune response to diverse viruses (AIV, IBDV, MDV, NDV). Bacteria such as Collinsella, Faecalibacterium, Oscillibacter, etc., increase the release of IFN-α, IFN-β, and IL-22. These substances control virus replication and repair mucosal tissue damage. Other bacteria, such as Clostridium XIVa or Firmicutes, provoke T-cells to produce anti-inflammatory cytokines to suppress inflammation. By promoting the antimicrobial peptides such as MUC, TFF, ZO, and tight junction proteins comprised of claudins, occludin, and zona occludens mRNA expression, Bacteroides, Candidatus, SMB53, Parabacteroides, Lactobacillus, Paenibacillus, Enterococcus, and Streptococcus spp. inhibit pathobiont colonization and translocation, and suppress inflammation. Butyrate succinate and lactate, produced by Faecalibacterium and Blautia spp., provide energy and reduce inflammation. Bacteroides fragilis produce bacterial polysaccharides that communicate with the immune system and influence the transformation of CD4+ (T-helper cells) and Foxp3+ cells (the master transcription factor of regulatory T cells in mammals, but also present in chicken (Burkhardt et al., 2022)).
“Negative” bacteria increase inflammation and enhance viral shedding Clostridium Cluster XI, Salmonella, and Shigella downregulate the anti-inflammatory and tight junction-stabilizing substances, which would be increased by the beneficial bacteria and increase IFN-γ and IF-17A to cause mucosal inflammation and tissue damage, as well as increased virus replication and fecal shedding. Further bacteria, which enhance mucosal and GIT inflammation, are Desulfovibrionaceae, producing hydrogen sulfides, Vampirovibrio, Clostridium cluster XIVb, and the genus Rumicoccus. They induce the pro-inflammatory cytokines IL-6 and IL-1β. The latter three bacteria also increase viral shedding. Salmonella typhimurium and Campylobacter jejuni also achieve higher viral shedding by decreasing viral-specific IgG and IgA production (Abaidullah et al., 2019)
Factors impairing intestinal immune defense
As the previous paragraph indicates, an imbalance of the intestinal microbiome called dysbiosis makes chickens more prone to diseases such as necrotic enteritis (Stanley et al., 2014). Several factors are disturbing the balance in the microbiome (Heinzl, 2020):
An abrupt change of feed
High contents of non-starch polysaccharides increase viscosity, decrease passage rate, lower the digestibility of other nutrients, and serve as nutrients for, e.g., Clostridium perfringens
High protein levels can also serve as a substrate for pathogens and cause a shift in the balance of the intestinal flora
Finely ground feed does not stimulate the gizzard muscles to do their work. pH increases, transit time decreases, and pathogenic microbes such as Salmonella, Campylobacter, and Clostridia proliferate.
Stress (heat or cold stress, re-assembling of groups, high stocking densities)
Mycotoxins
However, besides all these factors causing an overgrowth of commensal bacteria such as E. coli, ingested pathogens such as Marek’s or Newcastle Disease viruses can also cause this imbalance.
Immune defense in the gut – an interplay of different tools that must be protected
The first line of defense, the intestine, comprises different tools working together to fight pathogens and harmful substances. Besides the mucus layers and the specialized cells, the intestinal microbiome plays an essential role in immune defense by competing with pathogens for nutrients and binding sites, enhancing the secretion of anti-inflammatory substances, and stimulating the production of interferons, which fight the pathogens. However, several factors can impact the balance of the microbiome and cause dysbiosis. The best protection of this sensitive equilibrium can support the organism in defending against diseases and maintaining immunity and performance. Understanding the interplay between microbiota, immune function, and nutrition allows for effective strategies to enhance poultry health while reducing reliance on antibiotics. Future research will continue to provide insights into optimizing gut-immune interactions in poultry production.
References
Abaidullah, Muhammad, Shuwei Peng, Muhammad Kamran, Xu Song, and Zhongqiong Yin. “Current Findings on Gut Microbiota Mediated Immune Modulation against Viral Diseases in Chicken.” Viruses 11, no. 8 (July 25, 2019): 681. https://doi.org/10.3390/v11080681.
Baganz, Nicole L., and Randy D. Blakely. “A Dialogue between the Immune System and Brain, Spoken in the Language of Serotonin.” ACS Chemical Neuroscience 4, no. 1 (December 24, 2012): 48–63. https://doi.org/10.1021/cn300186b.
Berger, Miles, John A. Gray, and Bryan L. Roth. “The Expanded Biology of Serotonin.” Annual Review of Medicine 60, no. 1 (February 1, 2009): 355–66. https://doi.org/10.1146/annurev.med.60.042307.110802.
Broom, Leon J., and Michael H. Kogut. “The Role of the Gut Microbiome in Shaping the Immune System of Chickens.” Veterinary Immunology and Immunopathology 204 (October 2018): 44–51. https://doi.org/10.1016/j.vetimm.2018.10.002.
Burkhardt, Nina B, Daniel Elleder, Benjamin Schusser, Veronika Krchlíková, Thomas W Göbel, Sonja Härtle, and Bernd Kaspers. “The Discovery of Chicken Foxp3 Demands Redefinition of Avian Regulatory T Cells.” The Journal of Immunology 208, no. 5 (March 1, 2022): 1128–38. https://doi.org/10.4049/jimmunol.2000301.
Chabbi-Achengli, Yasmine, Amélie E. Coudert, Jacques Callebert, Valérie Geoffroy, Francine Côté, Corinne Collet, and Marie-Christine de Vernejoul. “Decreased Osteoclastogenesis in Serotonin-Deficient Mice.” Proceedings of the National Academy of Sciences 109, no. 7 (January 30, 2012): 2567–72. https://doi.org/10.1073/pnas.1117792109.
Clarke, G, S Grenham, P Scully, P Fitzgerald, R D Moloney, F Shanahan, T G Dinan, and J F Cryan. “The Microbiome-Gut-Brain Axis during Early Life Regulates the Hippocampal Serotonergic System in a Sex-Dependent Manner.” Molecular Psychiatry 18, no. 6 (June 2013): 666–73. https://doi.org/10.1038/mp.2012.77.
Diwakarla, S., L. J. Fothergill, J. Fakhry, B. Callaghan, and J. B. Furness. “Heterogeneity of Enterochromaffin Cells within the Gastrointestinal Tract.” Neurogastroenterology & Motility 29, no. 6 (May 9, 2017). https://doi.org/10.1111/nmo.13101.
Duangnumsawang, Yada, Jürgen Zentek, and Farshad Goodarzi Boroojeni. “Development and Functional Properties of Intestinal Mucus Layer in Poultry.” Frontiers in Immunology 12 (October 4, 2021). https://doi.org/10.3389/fimmu.2021.745849.
Heinzl, Inge. “Necrotic Enteritis: The Complete Overview.” EW Nutrition, August 8, 2023. https://ew-nutrition.com/necrotic-enteritis-complete-overview/.
Knecht, Leslie D., Gregory O’Connor, Rahul Mittal, Xue Z. Liu, Pirouz Daftarian, Sapna K. Deo, and Sylvia Daunert. “Serotonin Activates Bacterial Quorum Sensing and Enhances the Virulence of Pseudomonas Aeruginosa in the Host.” EBioMedicine 9 (July 2016): 161–69. https://doi.org/10.1016/j.ebiom.2016.05.037.
Kong, Shanshan, Yanhui H. Zhang, and Weiqiang Zhang. “Regulation of Intestinal Epithelial Cells Properties and Functions by Amino Acids.” BioMed Research International 2018 (2018): 1–10. https://doi.org/10.1155/2018/2819154.
Kwon, Yun Han, Huaqing Wang, Emmanuel Denou, Jean-Eric Ghia, Laura Rossi, Michelle E. Fontes, Steve P. Bernier, et al. “Modulation of Gut Microbiota Composition by Serotonin Signaling Influences Intestinal Immune Response and Susceptibility to Colitis.” Cellular and Molecular Gastroenterology and Hepatology 7, no. 4 (2019): 709–28. https://doi.org/10.1016/j.jcmgh.2019.01.004.
Linan-Rico, Andromeda, Fernando Ochoa-Cortes, Arthur Beyder, Suren Soghomonyan, Alix Zuleta-Alarcon, Vincenzo Coppola, and Fievos L. Christofi. “Mechanosensory Signaling in Enterochromaffin Cells and 5-HT Release: Potential Implications for Gut Inflammation.” Frontiers in Neuroscience 10 (December 19, 2016). https://doi.org/10.3389/fnins.2016.00564.
Liu, Yang, Xinjie Yu, Jianxin Zhao, Hao Zhang, Qixiao Zhai, and Wei Chen. “The Role of MUC2 Mucin in Intestinal Homeostasis and the Impact of Dietary Components on MUC2 Expression.” International Journal of Biological Macromolecules 164 (December 2020): 884–91. https://doi.org/10.1016/j.ijbiomac.2020.07.191.
Lyte, Mark. “Microbial Endocrinology in the Microbiome-Gut-Brain Axis: How Bacterial Production and Utilization of Neurochemicals Influence Behavior.” PLoS Pathogens 9, no. 11 (November 14, 2013). https://doi.org/10.1371/journal.ppat.1003726.
Marcobal, A., P. C. Kashyap, T. A. Nelson, P. A. Aronov, M. S. Donia, A. Spormann, M. A. Fischbach, and J. L. Sonnenburg. “A Metabolomic View of How the Human Gut Microbiota Impacts the Host Metabolome Using Humanized and Gnotobiotic Mice.” The ISME Journal 7, no. 10 (June 6, 2013): 1933–43. https://doi.org/10.1038/ismej.2013.89.
Stanley, Dragana, Shu-Biao Wu, Nicholas Rodgers, Robert A. Swick, and Robert J. Moore. “Differential Responses of Cecal Microbiota to Fishmeal, Eimeria and Clostridium Perfringens in a Necrotic Enteritis Challenge Model in Chickens.” PLoS ONE 9, no. 8 (August 28, 2014). https://doi.org/10.1371/journal.pone.0104739.
Wallaeys, Charlotte, Natalia Garcia‐Gonzalez, and Claude Libert. “Paneth Cells as the Cornerstones of Intestinal and Organismal Health: A Primer.” EMBO Molecular Medicine 15, no. 2 (December 27, 2022). https://doi.org/10.15252/emmm.202216427.
Yano, Jessica M., Kristie Yu, Gregory P. Donaldson, Gauri G. Shastri, Phoebe Ann, Liang Ma, Cathryn R. Nagler, Rustem F. Ismagilov, Sarkis K. Mazmanian, and Elaine Y. Hsiao. “Indigenous Bacteria from the Gut Microbiota Regulate Host Serotonin Biosynthesis.” Cell 163, no. 1 (September 2015): 258. https://doi.org/10.1016/j.cell.2015.09.017.
A holistic approach to animal health and nutrition: From feed issues to intestinal permeability – A conversation in Berlin
Recently, The Poultry Site’s Sarah Mikesell interviewed Predrag Persak, EW Nutrition’s Regional Technical Manager for Northern Europe. The conversation covered topics as wide as sustainability and challenges in poultry production, and as narrow as intestinal permeability. Thanks to The Poultry Site for the great talk!
Sarah Mikesell, The Poultry Site: Hi, this is Sarah Mikesell with The Poultry Site, and today we are here with Predrag Peršak. He is the Regional Technical Manager for Northern Europe with EW Nutrition. Thanks for being with us today, Predrag.
Predrag Peršak, EW Nutrition: Nice to be here, Sarah. Thank you for inviting me.
SM: Very good. It’s nice to visit with you. And today, Predrag and I are in Berlin, Germany, at an exclusive event for the poultry industry called Producing for the Future, which is sponsored by EW Nutrition. You are one of our speakers today, Predrag, so I’m going to ask you just a few questions to let everybody know a little bit about your presentation.
You’ve described animal nutrition as “never boring and never finished.” What makes this field so dynamic and constantly evolving for you?
PP: I’ve been in animal nutrition for about 25 years. And in those 25 years, I would say that not even half a year passed without something extraordinary happening. From genetics to animal husbandry, especially here in Europe, we also have a lot of pressure from consumers and slaughterhouses to adapt production to the needs of the customers.
Sustainability, sourcing raw materials, and the variety of raw materials available in Europe – and the constant development of new ones – make life for an animal nutritionist very, very interesting. It’s also very challenging, and through these challenges you learn a lot.
So, applying what we learned 20 years ago is simply not enough anymore. For someone who wants to be challenged every day with new things, this is definitely the right industry to be in – especially now.
SM: Excellent. Can you explain your holistic approach to animal nutrition and how considering multiple factors benefits practical applications on farms?
PP: The concept of a holistic approach in animal nutrition is not new. But for me – being both a veterinarian and a nutritionist – it means having deeper insight into the animal itself, into all the metabolic processes, and also into the external influences: husbandry, genetics, diseases, and management. Looking at how all of these interact, we can only really solve problems by looking at the animal as a whole system.
The same applies to feed production. You cannot look at a feed mill as just one compartment. You have to look at sourcing raw materials, their quality, how they are processed – milling, pelleting, and other technologies – and then see how that feed performs on the farm.
So, a holistic approach can be applied both from the animal perspective and from the feed production perspective, across all steps and processes. This is something we use and promote daily in our work with customers.
SM: Very good. You’ve worked with unconventional protein and fiber sources. We’re hearing a lot more about that recently. What are those, and what potential do they bring to animal nutrition?
PP: When I talk about unconventional protein and fiber sources, we need to remember that the global feed production scene is very diverse. What applies in the U.S. or Brazil does not necessarily apply in Europe or the Far East.
Here in Europe, we try to use not by-products but co-products of food production. For example, different fractions of rapeseed or sunflower meal, which are widely produced in Europe but not often used by mainstream nutritionists due to certain limitations. By finding the right processing methods and combining them with technologies, we can make these unconventional materials usable in mainstream nutrition.
The same goes for fiber sources. Both fermentable and structural fibers are increasingly important for intestinal and digestive development, as well as for overall animal health. So, processing fibers in ways that maximize usability while minimizing negative effects is a big part of my work.
SM: From a cost standpoint for producers, are those lower-cost inputs, or just alternatives they need to look at?
PP: In Germany we have a perfect expression for this: “yes and no.” There is always pressure on price, especially in poultry, because food must be accessible to everyone. But at the same time, food must not harm the environment or human health, and we should use all resources not fit for humans but still usable for animals.
So, it’s not only about cost – about availability and sustainability. Working with just two, three, or five raw materials for a long time is not the way forward. The way forward is to think of everything that can be used properly, for the benefit of the animals, and ultimately to produce enough food for the world.
Also, using locally available products is important. Feed production is very diverse around the world—raw materials in Southeast Asia differ completely from those in Europe, Brazil, or the U.S. Using technologies to enable the use of locally produced by-products makes production not only sustainable, but also economically viable for local communities. That’s really the core of the feed industry: using what is produced locally.
SM: Interesting. Very cool. How does your interdisciplinary work across poultry, pigs, and ruminants give you unique insights that might be missed with a narrower focus?
PP: I come from a small feed mill in a small country, Croatia. There, you don’t have deep specialization by species or even by category, as you find in larger markets. Specialization has its advantages, but it can also limit creativity and “outside-the-box” thinking.
By working with ruminants, I learned about fermentation processes – knowledge that can be applied to pigs and even to poultry. For example, fermentation can reduce anti-nutritional factors, allowing higher inclusion levels of certain raw materials in poultry diets.
With pigs, fermentation of fibers – especially in piglets – is crucial, and some of that knowledge could be applied to turkeys, where we still face health issues.
So, working across species demands a lot – it leaves little time for other things – but it opens up unique perspectives and cross-species applications that benefit the entire livestock industry.
SM: I was talking with someone yesterday about mycotoxins – there’s a lot of research in pigs but less in poultry. That’s kind of what you’re talking about, right? Applying knowledge across species?
PP: Absolutely. We’re focused now on poultry, but we can learn from poultry too – not only about feeding but also about farm management, biosecurity, and more. These lessons can also apply to pigs or ruminants.
It’s all holistic – you cannot solve everything with nutrition alone. It’s always a package.
SM: You presented today about the importance of intestinal permeability. Why is it important, and how can understanding it impact animal health and performance outcomes?
PP: Intestinal permeability is one of the key features we use to describe gut health. Personally, I’m very practical. For 20 years we’ve talked about “gut health,” but the real question for veterinarians and nutritionists is: what do we actually do with that knowledge?
In my presentation, I explained intestinal permeability as a “point of no return” in gut health. When leaky gut develops, everything else can deteriorate – faster or slower – but it won’t return to normal without intervention.
By comparing how different stressors or pathogens impact intestinal permeability, we can better understand severity and decide where to focus. Nutritionists already pay attention to thousands of factors, but we need to identify the most impactful ones. That was my key message: focus on the most important drivers.
SM: And leaky gut has really become something the whole industry is talking about, right? I’ve even seen it in human health – my doctor has posters about it.
PP: Exactly. Across cows, pigs, and poultry, leaky gut is getting a lot of attention. It’s a physiological or pathophysiological feature that marks the point of no return.
We can talk about dysbiosis and all the causes, but once you reach leaky gut, you understand where intervention is needed. And it’s not just hype. For example, recently Nature published research showing certain types of human bone marrow conditions are linked to leaky gut and microbial influence on blood processes.
So, this is not a passing trend. It’s fundamental. And once we solve one issue, another door opens. That’s why this industry is never boring.
SM: Very good. Well, thank you for all the information today, Predrag.
PP: Thank you, Sarah. It was a pleasure to talk with you.
Mycotoxins as contributors to antibiotic resistance?
By Dr. Inge Heinzl, Editor EW Nutrition and
Marie Gallissot, Global Manager Feed Quality Solutions EW Nutrition
Antibiotic resistance is a growing global health concern, making infections more complicated to treat and increasing the risk of disease spread, severe illness, and death. While overuse and misuse of antibiotics are the primary causes, recent research has uncovered another unexpected contributor: mycotoxins. Among these, deoxynivalenol (DON), a toxin commonly found in contaminated grains, has been shown to significantly alter gut microbiota and promote antibiotic resistance. This article examines how DON impacts gut bacteria, influences antibiotic resistance, and highlights why this issue warrants urgent attention.
Mycotoxins – originators of antimicrobial resistance?
Actually, it would be logical…
Alexander Fleming discovered Penicillin when he returned after the summer holidays and saw that a mold had grown on the agar plate he had prepared. Around the mold, Staphylococcus was unable to proliferate. The reason was a substance produced by the mold – penicillin, which, like other toxins produced by molds, is a mycotoxin. In his article about the origin of antibiotics and mycotoxins, Shier (2011) stated that antibiotics and mycotoxins share considerable similarities in structure, metabolic roles, and biosynthesis.
A short excursus to antimicrobial resistance
In general, the primary mechanisms of resistance involve the prevention or limitation of the antimicrobial substance’s uptake, modifying the drug target, inactivating the drug, or facilitating its discharge with efflux pumps.
There are two types of resistance: natural resistance, which is further divided into intrinsic and induced resistance, and acquired resistance.
Intrinsic resistance is a “characteristic” of a bacterial species and is not dependent on antibiotic exposure. An example is the reduced permeability of the outer membrane of gram-negative bacteria, which prevents certain antibiotics from entering.
Induced resistance, however, needs to be initiated by antibiotics. Here, multidrug-efflux pumps can be mentioned.
The third one, acquired resistance, refers to the process by which bacteria acquire genetic material, the resistance genes, from other bacteria that are resistant. The mechanisms include vertical transfer to daughter cells and horizontal transfer, such as the transfer from dead bacteria to living ones, by viruses, or the transfer of plasmids (Reygaert, 2018).
Figure 1: Different possibilities of transfer of resistance genes
Deoxynivalenol (DON) promotes resistance in gut microbiota
A Chinese group of researchers (Deng et al., 2025) examined for the first time the influence of DON on the intestinal microbiota of chickens. One of the most alarming findings is DON’s ability to enhance antibiotic resistance. It contributes to this issue in several ways:
Encouraging resistant bacteria – By disrupting microbial balance, DON provides a survival advantage to bacteria that carry resistance genes.
Activating resistance genes – Studies suggest that DON can increase the expression of genes that help bacteria withstand antibiotics.
Enhancing gene transfer – Bacteria can share resistance genes through horizontal gene transfer. DON appears to promote this process, making antibiotic-resistant strains spread more rapidly.
Weakening antibiotic effectiveness – DON-induced changes in the gut environment can reduce the effectiveness of antibiotics, making treatments less successful.
A further indication that mycotoxins can enhance resistance is the significant overlap in the geographical distribution of antimicrobial-resistant bacteria and genes with that of mycotoxins, as noted by Deng et al.
Which protection mechanisms do bacteria have against mycotoxins?
In the case of mycotoxins, bacteria employ similar molecular mechanisms to those used against antibiotics. In an in vitro experiment, Hassan et al. (2019) challenged Devosiamutans, a gram-negative bacterium, with DON in the growth medium. DON inhibits protein synthesis, induces oxidative stress, and compromises cell membrane integrity in eucaryotic cells. Hassan et al. asserted three adaptive mechanisms as the response to the challenge:
Activation of cellular membrane proteins (adenosine 5’-triphosphate-binding cassette -ABC- transporters) responsible for the unidirectional transport of substrates, either outward or inward. These ABC transporters can work as drug efflux pumps.
Production of DON-specific deactivation enzymes, thereby engaging a toxin-specific pyrroloquinoline quinone-dependent detoxification pathway. This enables the bacterial isolate to transform DON to a non-toxic stereoisomer.
Upregulation of auxiliary coping proteins, such as porins (transmembrane proteins involved in metabolite exchange), glutathione S-transferases, and phosphotransferases, both of which are likely involved in the detoxification of xenobiotics.
Public health implications and preventive measures
Given the widespread presence of DON in food and animal feed, its potential role in antibiotic resistance poses a serious threat. The combination of increased bacterial resistance and weakened antibiotic efficacy could lead to more difficult-to-treat infections. This is particularly concerning in hospital settings, where antibiotic-resistant infections already cause high mortality rates.
To address the issue, several strategies can be implemented:
Reducing DON contamination: Implementing improved agricultural practices, such as crop rotation, the use of fungal-resistant crop varieties, and maintaining proper storage conditions, can help limit fungal growth and DON production.
Monitoring food and feed supply – Strict regulations and testing for DON contamination in grains and animal feed are essential to minimize human and animal exposure.
Maintaining gut health: A healthy diet rich in fiber, probiotics, and gut health-supporting feed supplements, such as Ventar D or products from the Activo line, may help counteract some of the adverse effects of DON on gut microbiota.
Developing new treatments: Research into alternative therapies and new antibiotics is crucial to combat the rise of antibiotic resistance.
Antimicrobial resistance: Be aware of the mycotoxins!
The connection between mycotoxins, such as DON, and antibiotic resistance underscores the need for a broader perspective on public health and food safety and once again brings the “One Health Concept” into focus. While antibiotic overuse remains the primary driver of resistance, environmental factors, such as exposure to mycotoxins, should not be overlooked. By increasing awareness, enhancing food safety regulations, and investing in research, we can take steps to mitigate this emerging threat and safeguard the effectiveness of antibiotics for future generations.
References:
Deng, Fengru, Chuying Yao, Linyu Ke, Meichan Chen, Mi Huang, Jikai Wen, Qingmei Chen, Jun Jiang, and Yiqun Deng. “Emerging Threat to Antibiotic Resistance: Impact of Mycotoxin Deoxynivalenol on Gut Microbiota and Clonal Expansion of Extensively Drug-Resistant Enterococci.” Environment International 197 (March 2025): 109353.
https://doi.org/10.1016/j.envint.2025.109353.
Hassan, Yousef I., Jian Wei He, Dion Lepp, and Ting Zhou. “Understanding the Bacterial Response to Mycotoxins: The Transcriptomic Analysis of Deoxynivalenol-Induced Changes in Devosia Mutans 17-2-E-8.” Frontiers in Pharmacology 10 (November 14, 2019).
https://doi.org/10.3389/fphar.2019.01098.
Reygaert, Wanda C. “An Overview of the Antimicrobial Resistance Mechanisms of Bacteria.” AIMS Microbiology 4, no. 3 (2018): 482–501.
https://doi.org/10.3934/microbiol.2018.3.482.
Shier, W. Thomas. “On the Origin of Antibiotics and Mycotoxins.” Toxin Reviews 30, no. 1 (January 28, 2011): 6–30.
https://doi.org/10.3109/15569543.2011.550862.
Smith, William P., Benjamin R. Wucher, Carey D. Nadell, and Kevin R. Foster. “Bacterial Defences: Mechanisms, Evolution and Antimicrobial Resistance.” Nature Reviews Microbiology 21, no. 8 (April 24, 2023): 519–34.
https://doi.org/10.1038/s41579-023-00877-3.
The big challenge: Keeping sows healthy and productive – Part 2 Nutritional interventions – Phytomolecules
Dr. Inge Heinzl – Editor of EW Nutrition, and Dr. Merideth Parke – Global Application Manager for Swine, EW Nutrition
The first of the two articles focused on general aspects to be observed to achieve a particular stock of healthy and well-performing sows, as well as high productivity on the farm. In addition to general measures, feed supplements can be used to further support the sows. Phytomolecules with characteristics supporting gut and overall health are effective for this purpose.
Phytomolecules – how can they help?
Phytogenics, also known as phytomolecules, are plant-derived, natural bioactive compounds that promote livestock health and well-being, as well as improve growth performance and production efficiency. Phytomolecules encompass a diverse range of compounds, including terpenes, phenols, glycosides, saccharides, aldehydes, esters, and alcohols.
The literature describes some of their effects, including stimulation of digestive secretions, immune stimulation and anti-inflammatory activity, intestinal microflora modulation, and antioxidant effects (Durmic and Blanche, 2012; Ehrlinger, 2007; Zhao et al., 2023), as well as estrogenic and hyperprolactinemic properties (Farmer, 2018) and effects on colostrum and milk porcine sensory profiles (Val-Laillet et al., 2018). They represent exciting antibiotic alternatives in swine production (Omonijo et al., 2018).
1. Phytomolecules modulate intestinal microbiota
Phytomolecules are microbiome modulators through different mechanisms. They can directly impact pathogenic bacteria by damaging the cell membrane, cell wall, or cytoplasm, interrupting the anion exchange, resulting in changes to cellular pH, and inhibiting the cell’s energy production system. Additionally, phytomolecules interfere with the virulence capacity of pathogenic bacteria through the indirect quorum quenching mechanism. (Rutherford and Bassler, 2012).
The favorable consequence of this differential microbial modulation is maintaining gut microbiome diversity, shifting it to a bacterial population with reduced pathogenic and increased beneficial microbes.
Proof of Ventar D’s pathogen-inhibiting effect
An in vitro study evaluated the effect of Ventar D on pathogenic Clostridium perfringens and beneficial Lactobacillus spp.
Process
To test the effect of Ventar D on four different beneficial Lactobacillus spp., and pathogenic Clostridium perfringens, the phytogenic formulation (Ventar D) was added to the respective culture medium in the following concentrations: 0 µg/mL – control, 500 µg/mL (only C. perfr.), 750 µg/mL, 1000 mg/mL (only C. perfr.), and 1250 µg/mL.
After cultivating the bacteria in the culture medium, the colony-forming units (CFU) were counted.
Results and discussion
The study demonstrated a dose-dependent decrease in the Clostridium perfringens population. At the lowest tested concentration (500 µg/mL), Ventar D’s antimicrobial effect was already detectable; at 750 µg/mL, scarce colonies were observed; and at 1000 µg/mL, C. perfringens could no longer grow.
Figure 1: Effect of Ventar D on Clostridium perfringens
In contrast, even at higher concentrations of Ventar D, the beneficial L. agilis S73 and L. agilis S1 populations were only mildly affected, and L. casei and L. plantarum were unaffected.
Figure 2: Effect of Ventar D on Lactobacillus spp.
These findings confirm the differential antimicrobial activity of Ventar D’s formulation, specifically a bactericidal effect on pathogenic C. perfringens populations and a mild to no inhibition of beneficial Lactobacillus spp.
2. Phytomolecules improve intestinal integrity
The gut barrier is semipermeable and is responsible for immune sensing and regulating the movement of nutrients and undesirable microbes and substances.
The “gatekeepers” are tight junctions (TJ), adherent junctions (AJ), and desmosomes situated between the intestinal enteric cells (IEC). The tight junctions regulate the transport of small molecules and ions. The adherent junctions and desmosomes maintain the integrity of the intestinal barrier by keeping the IECs together through cell-adhesion bonds.
Oxidative stress resulting from factors such as heat stress or fat oxidation in the feed, as well as dysbacteriosis caused by changes in diet, out-of-feed events, poor dietary formulation, or bacterial contamination, can compromise the integrity of these critical adhesions and junctions between enterocytes.
The support of these tight junctions prevents bacteria and toxins from passing into the organism. Besides reducing disease occurrence, it also reduces the activation of the immune system and inflammatory processes. Ingested nutrients can be used for growth and need not be spent for the defense of the organism.
Proof of Ventar D’s gut barrier-stabilizing effect
An experiment was conducted to determine the level of tight junction gene expression biomarkers closely related to gut integrity.
Process
The experiment was conducted in broilers. They were fed 100 g of Ventar D/ t of feed, and the gene expression of Claudin and Occludin was measured (the higher the gene expression, the higher the level of gut barrier damage).
Figure 3: Effect of Ventar D on gut barrier function
Results
The lower levels of both gut tight junction gene expression biomarkers, Claudin and Occludin, in Ventar D-supplemented birds support a lower level of damage and a more robust gut barrier function (Figure 3).
3. Phytomolecules act as antioxidants
As mentioned, oxidative stress can disrupt gut barrier function and negatively impact the health of sows and piglets. Therefore, it is vital to scavenge reactive oxygen species (ROS) to reduce the damage these free radicals can cause to enterocytes and tight junctions.
Proof of Ventar D’s antioxidant effect in vitro
In this case, an in vitro trial was conducted to show Ventar D’s antioxidant effects.
Process
Ventar D’s antioxidant activity was tested in vitro using the ORAC (Oxygen Radical Absorbent Capacity) test. The ORAC test measures the antioxidant activity of a compound compared to that of the Vitamin E analog Trolox.
Result
The components in Ventar D demonstrated its capacity as an antioxidant, with a more substantial effect than the Vitamin E analog Trolox (see Figure 4).
Figure 4: Antioxidant capacity of Ventar D compared to Vit. E analogue (AUC – Area under curve)
4. Phytomolecules decrease inflammation
In intensive production, animals face daily inflammation associated with various stressors, including gut incidents and intestinal dysbiosis, social hierarchy-associated fighting resulting in musculoskeletal or skin injuries, farrowing and lactation trauma to reproductive organs, and diseases affecting any system in the pig.
Animals with high-performance expectations, such as gestating, farrowing, and lactating sows, are particularly susceptible to high nutrient diversion, which can lead to inflammation and activation of the immune system. To mitigate the excessive continuation of inflammatory processes, phytomolecules with anti-inflammatory effects can be utilized.
Proof of Ventar D’s anti-inflammatory effect in vitro
The anti-inflammatory effect of Ventar D was shown in an in vitro trial conducted in the Netherlands.
Process
For the trial, cells from mice (Murine macrophages, RAW264.7) were stressed with lipopolysaccharides (LPS, Endotoxin) from E. coli O111:B4 (0.25 µg/ml) to provoke an immune response. To evaluate the effects of Ventar D, two different concentrations (50 and 200 ppm) were tested, and the levels of NF-κB, IL-6, and IL-10 were determined. IL-6 and IL-10 could be measured directly using specific ELISA kits, whereas, in the case of NF-κB activity, an enzyme induced by NF-κB (secreted embryonic alkaline phosphatase – SEAP) was used for measurement. The trial design was as follows (Figure 5):
Figure 5: Trial design
Results
The trial results showed a dose-dependent reduction of NF-κB activity in LPS-stimulated mouse cells, with 11% and 54% reductions at 50 and 200 ppm Ventar D, respectively. The pro-inflammatory cytokine IL-6 was downregulated, and the anti-inflammatory cytokine IL-10 was upregulated by 84% and 20%, respectively, resulting in a decrease in the IL-6 to IL-10 ratio. This ratio is essential in balancing the pro- and anti-inflammatory outcomes of cellular signaling.
Figure 6: Activity of NFκB
Figure 7: IL-6/IL-10 ratio
5. Phytomolecules improve production performance and efficiency
The intensive production systems of today encompass many factors that create stress in the animals. Phytomolecules exhibiting the positive characteristics mentioned in points 1 to 4 result in better performance in animals.
In pigs in suboptimal conditions, the antimicrobial effect of phytomolecules is the most important. However, in pigs held under optimal conditions and with extraordinary growth, the antioxidant and anti-inflammatory effects are most relevant. Anabolic processes, driven by strong growth, increase oxidative stress, while non-infectious inflammations burden the immune system.
Proof of Ventar D’s performance-promoting effect in pigs
To evaluate growth-promoting effects in pigs, a study was conducted on a commercial farm in the United States.
Process
A total of 532 approx. 24-day-old weaned piglets were housed in 28 pens, each containing 19 non-castrated males or gilts. Piglets were blocked by body weight and fed a three-phase feeding program (Table 1). Phase 1 and 2 diets were pellets, and phase 3 was mash. Diets were based on corn and soybeans, and a concentrate including soy protein concentrate, whey permeate, and fish meal was added in phases 1 and 2, at a ratio of 50% of the total feed in phase 1 and 25% in phase 2. No feed medication was used in this trial.
Table 1: Feeding scheme and product application
Trial groups
Feeding Phase 1 (day1 – day 14)
Feeding Phase 2 (day 15 – day 24)
Feeding Phase 3
Control
No additive
No additive
No additive
Ventar D
Ventar D 200 g/MT
Ventar D 200 g/MT
Ventar D 200 g/MT
Results
Adding Ventar D increased final body weight and improved FCR (see Figures 8 to 10). Furthermore, the addition of Ventar D to the feed reduced mortality.
Figures 8-10: Performance of piglets fed Ventar D in comparison to a negative control
Phytomolecules can help to keep sows healthy and productive
Intensive animal production places a significant strain on animal organisms. High stocking density often accompanies high pathogenic pressure and stress, and high growth performance can lead to increased oxidative stress and inflammation. It isn’t easy to keep all challenges under control. However, phytomolecules can be a solution as their modes of action cover different relevant topics.
References
Durmic, Z., and D. Blache. “Bioactive Plants and Plant Products: Effects on Animal Function, Health and Welfare.” Animal Feed Science and Technology 176, no. 1–4 (September 2012): 150–62. https://doi.org/10.1016/j.anifeedsci.2012.07.018.
Farmer, Chantal. “Nutritional Impact on Mammary Development in Pigs: A Review.” Journal of Animal Science 96, no. 9 (June 15, 2018): 3748–56. https://doi.org/10.1093/jas/sky243.
Omonijo, Faith A., Liju Ni, Joshua Gong, Qi Wang, Ludovic Lahaye, and Chengbo Yang. “Essential Oils as Alternatives to Antibiotics in Swine Production.” Animal Nutrition 4, no. 2 (June 2018): 126–36. https://doi.org/10.1016/j.aninu.2017.09.001.
Rutherford, S. T., and B. L. Bassler. “Bacterial Quorum Sensing: Its Role in Virulence and Possibilities for Its Control.” Cold Spring Harbor Perspectives in Medicine 2, no. 11 (November 1, 2012). https://doi.org/10.1101/cshperspect.a012427.
Val-Laillet, David, J Stephen Elmore, David Baines, Peter Naylor, and Robert Naylor. “Long-Term Exposure to Sensory Feed Additives during the Gestational and Postnatal Periods Affects Sows’ Colostrum and Milk Sensory Profiles, Piglets’ Growth, and Feed Intake1.” Journal of Animal Science, June 29, 2018. https://doi.org/10.1093/jas/sky171.
Zhao, Bi-Chen, Tian-Hao Wang, Jian Chen, Bai-Hao Qiu, Ya-Ru Xu, Qing Zhang, Jian-Jie Li, Chun-Jiang Wang, Qiu-Feng Nie, and Jin-Long Li. “Effects of Dietary Supplementation with a Carvacrol–Cinnamaldehyde–Thymol Blend on Growth Performance and Intestinal Health of Nursery Pigs.” Porcine Health Management 9, no. 24 (May 23, 2023). https://doi.org/10.1186/s40813-023-00317-x.
Enhancing Poultry Gut Health with Novel Xylanase: A Sustainable Path to Reduced Antimicrobial Use
By Ajay Bhoyar, Senior Global Technical Manager, EW Nutrition
Gut health is pivotal to profitable poultry production, as the gastrointestinal tract (GIT) enables nutrient digestion and absorption while acting as a defense against pathogens. A healthy gut improves feed conversion, boosts immune resilience, and reduces reliance on antimicrobials—critical in the fight against antimicrobial resistance (AMR). With AMR posing significant threats to public health and animal agriculture, strategies like biosecurity, sustainable management, and effective dietary interventions are gaining traction. Feed enzymes have emerged as essential tools for managing feed costs, mitigating anti-nutritional factors, and improving nutrient utilization. Among these, feed enzymes like xylanase stand out. By breaking down xylan, a major component of non-starch polysaccharides (NSPs) in plant-based feed ingredients, xylanase reduces gut viscosity, enhances nutrient utilization, and supports optimal gut health and productivity. This article explores the innovative application of novel GH10 xylanases, such as Axxess XY, as a sustainable solution for improving feed efficiency and gut health in poultry production.
Xylanase in Poultry Nutrition
Xylanase plays a pivotal role in enhancing nutrient availability by addressing the limitations of endogenous enzyme synthesis in poultry. Xylanase enzymes belong to the carbohydrase class, catalyzing the breakdown of xylan, a major NSP in plant-based feed ingredients. They hydrolyze xylan into simple sugars like arabino-xylo-oligosaccharides (AXOs) and xylo-oligosaccharides (XOs), reducing the encapsulation of nutrients and digesta viscosity. These actions improve overall nutrient digestibility and bird performance.
Fig.1: Arabinoxylans – anti-nutrient mode of action in chicken
The primary benefit of feed xylanase lies in its ability to reduce digesta viscosity. By partially hydrolyzing NSPs in the upper digestive tract, xylanase ensures better nutrient absorption in the small intestine. Studies (Matthiesen et al., 2021; Choct & Annison, 1992) confirm that reduced viscosity enhances feed digestibility, leading to improved performance in poultry. Further, to realize the optimum benefits, it is crucial that xylanase efficiently degrades both soluble and insoluble arabinoxylans. The insoluble arabinoxylans are part of the cell wall structure of plant cells, resulting in a cage effect, entrapping nutrients like starch and protein. Effectively breaking down insoluble arabinoxylans ensures that the nutrients trapped in plant cell walls are released for growth and production.
Mechanisms Supporting Gut Health
Viscosity Reduction
High NSP content increases digesta viscosity and slows digestion and nutrient absorption. Soluble arabinoxylan is not digested in the small intestine of broilers. It produces a viscous chime, leading to the proliferation of pathogenic bacteria, intestinal inflammation, impairment of barrier function in the intestine, and severe intestinal lesions (Teirlynck et al., 2009). Xylanase mitigates this by breaking down xylans, a major component of NSPs in common feed ingredients. This results in a better flow of digesta and reduced energy losses.
Microbial Metabolites
Xylo-oligosaccharides (XOS) can also be produced in the intestine of monogastric animals to some extent when exogenous enzymes, such as xylanase, are added to the feed (Baker et al., 2021).
The XOS generated by xylanase action on arabinoxylans can act as prebiotics, fostering beneficial bacteria like Lactobacillus and Bifidobacterium, which can outcompete harmful species. XOS can positively impact the gut microbiota, enhance short-chain fatty acid (SCFA) production, stimulate immune activity in the gastrointestinal tract, and improve energy utilization.
Fig. 2. Axxess XY improved beneficial microbes and reduced the clostridial population in broilers.
Barrier Function
By lowering inflammation and irritation in the intestine, xylanase helps maintain gut integrity, reducing the risk of pathogen translocation from the intestinal lumen. In a broiler study, xylanase decreases epithelial apoptosis index, up-regulates tight junction gene expression, and inhibits mucin synthesis in the small intestine, likewise alleviating the intestinal mucosal barrier impairment from Clostridium perfringens challenge (Liu et al., 2012).
Practical Considerations for Xylanase Use
Enzyme Stability
Enzymes are proteins that tend to lose their catalytic activity at high temperatures. When exposed to excessive heat, an enzyme’s protein structure can irreversibly unfold, disrupting its active site and causing loss of function. Therefore, ensuring enzyme stability during feed processing is critical for maintaining its activity in the intestine. Intrinsically heat-stable enzymes have an inherent ability to withstand higher temperatures without the need for a protective coating and are immediately available for action upon ingestion.
Feed Composition
Xylanase efficacy is influenced by diet composition, particularly the NSP content and the presence of xylanase inhibitors in common feedstuffs. It is important to choose a xylanase that can resist the activity of xylanase inhibitors and is effective against both soluble and insoluble arabinoxylans.
The recommended energy matrix value for the xylanase enzyme should be used while formulating the feeds to create energy-deficient diets to reap the full benefits of xylanase use.
Optimal Dosage
Proper dosing is essential to maximizing the benefits of feed enzymes while avoiding unnecessary costs. It is important to follow manufacturers’ recommendations and avoid underdosing an enzyme.
GH10 Xylanases: The Superior Choice for Animal Nutrition
Most feed xylanases are classified into glycoside hydrolase families 10 (GH10) and 11 (GH11) based on their substrate specificity, catalytic action, and structural features.
Why GH10 Xylanases Are More Effective
Broader Substrate Specificity:
Unlike GH11 xylanases, GH10 xylanases can effectively hydrolyze both soluble and insoluble xylan substrates. This broader activity ensures an efficient breakdown of xylans in a wide range of feed ingredients.
Higher Catalytic Efficiency:
GH10 enzymes cleave xylan at substituted regions, yielding shorter xylo-oligosaccharides that can positively impact gut health and maximize nutrient availability.
Thermostability:
Feed processing often involves high temperatures during pelleting. Axxess XY, a GH10 family xylanase, demonstrates remarkable thermostability, maintaining over 85% activity even at 95°C for extended conditioning times. This resilience ensures consistent enzyme performance during feed manufacturing and digestion.
Fig.3: Optimum recovery of Axxess XY at elevated conditioning time and temperatures
Novel Applications of Axxess XY: A GH10 Xylanase
Axxess XY exemplifies the advantages of GH10 xylanases in poultry nutrition. Its ability to efficiently act on both soluble and insoluble arabinoxylans makes it a versatile feed enzyme. The enzyme’s high thermostability ensures efficient enzyme activity in the gut and subsequent optimum nutrient utilization under challenging processing conditions, promoting gut health and maximizing performance.
Key Benefits of Axxess XY
Enhanced Nutrient Utilization:
By unlocking nutrients trapped in NSPs, Axxess XY promotes better feed conversion ratios (FCRs).
Improved Gut Health:
Reducing the digest’s viscosity reduces gut health challenges and predisposition to gut infections. Further, the short-chain oligosaccharides released by Axxess XY support beneficial gut microbiota, improving digestive health.
Economic Efficiency:
Enabling the optimum use of high-fiber, cost-effective, locally available feed ingredients without compromising performance makes Axxess XY an asset for profitability.
In a recently conducted 42-day trial at a commercial farm, Axxess XY maintained the average body weight of broilers with a 100 kcal/kg reduction in metabolizable energy while significantly reducing feed cost/kg body weight. The diets were based on corn, DDGS, and soybean meal.
Figures 4 and 5: Body weight and cost of feed in broilers fed a diet reduced by 100 kcal/kg in metabolizable energy compared to a standard diet without Axxess XY
Conclusion
Xylanase exemplifies how feed enzymes can transcend their traditional role in feed cost reduction to support enhanced gut health. Xylanase supports reduced antimicrobial use in poultry production by improving nutrient utilization, reducing digesta viscosity, and fostering healthy microbiota. Its integration into comprehensive gut health management strategies offers a sustainable pathway to combat AMR and ensure the long-term viability of poultry farming. By targeting NSPs, these enzymes enhance nutrient digestibility, reduce feed costs, and support sustainable production practices.
GH10 xylanases, particularly Axxess XY, stand out for their superior substrate specificity, catalytic efficiency, and thermostability. By incorporating Axxess XY into feed formulations, poultry producers can unlock the full nutritional potential of feed ingredients, ensuring optimal performance and profitability. As the poultry industry continues to evolve, adopting advanced enzyme technologies like Axxess XY represents a strategic step toward sustainable and efficient animal nutrition.
References:
Baker, J.T.; Duarte, M.E.; Holanda, D.M.; Kim, S.W. Friend or Foe? Impacts of Dietary Xylans, Xylooligosaccharides, and Xylanases on Intestinal Health and Growth Performance of Monogastric Animals. Animals 2021, 11, 609.
Choct, M., and G. Annison. “Anti‐nutritive Effect of Wheat Pentosans in Broiler Chickens: Roles of Viscosity and Gut Microflora.” British Poultry Science 33, no. 4 (September 1992): 821–34. https://doi.org/10.1080/00071669208417524.
Liu D, Guo S, Guo Y. Xylanase supplementation to a wheat-based diet alleviated the intestinal mucosal barrier impairment of broiler chickens challenged by Clostridium perfringens. Avian Pathol. 2012;41(3):291-8.
Matthiesen, Connie F., Dan Pettersson, Adam Smith, Ninfa R. Pedersen, and Adam. C. Storm. “Exogenous Xylanase Improves Broiler Production Efficiency by Increasing Proximal Small Intestine Digestion of Crude Protein and Starch in Wheat-Based Diets of Various Viscosities.” Animal Feed Science and Technology 272 (February 2021): 114739. https://doi.org/10.1016/j.anifeedsci.2020.114739.
Teirlynck, E.; Haesebrouck, F.; Pasmans, F.; Dewulf, J.; Ducatelle, R.; van Immerseel, F. The cereal type in feed influences Salmonella enteritidis colonization in broilers. Poult. Sci. 2009, 88, 2108–2112.
Sustainability will push more by-products into pig feed – Keep track of mycotoxins!
Mycotoxin Team EW Nutrition
Most grains used in feed are susceptible to mycotoxin contamination, causing severe economic losses all along feed value chains. As skyrocketing raw material prices force producers to include a higher proportion of economical cereal by-products in the feed, the risks of mycotoxin contamination likely increase. This article reviews why mycotoxins cause the damage they do – and how effective toxin-mitigating solutions prevent this damage.
Mycotoxin contamination of cereal by-products requires solutions
Cereal by-products may become more important feed ingredients as grain prices increase. However, from a sustainability point of view and considering population growth, using cereal by-products in animal feed makes much sense. Distiller’s dried grains with solubles (DDGS) are a good example of how by-products from food processing industries can become high-quality animal feed.
Figure 1: By-products are a crucial protein source (data from FEFAC Feed&Food 2021 report)
Still, research on what happens to mycotoxins during food processing shows that mycotoxins are concentrated into fractions that are commonly used as animal feed (cf. Pinotti et al., 2016; Caballero and Heinzl, 2022). To safeguard animal health and performance when feeding lower-quality cereals, monitoring mycotoxin risks through regular testing and using toxin-mitigating solutions is essentialy.
Problematic effects of mycotoxins on the intestinal epithelium
Mostmycotoxinsare absorbedin the proximal partof the gastrointestinal tract. Thisabsorption can behigh, as in the case of aflatoxins(ca. 90%), but also very limited, as in the case of fumonisins(< 1%); moreover, it depends on the species.Notably, a significant portion ofunabsorbedtoxinsremains within thelumenof the gastrointestinal tract.
Importantly, studies based on realistic mycotoxin challenges (e.g., Burel et al., 2013) show that the mycotoxin levels necessary to trigger damaging processes are lower than the levels reported as safe by EFSA, the Food Safety Agency of the European Union. The ultimate consequences range from diminished nutrient absorption to inflammatory responses and pathogenic disorders in the animal (Figure 2).
Figure 2: Mycotoxins’ impact on the GIT and consequences for monogastric animals
1. Alteration of the intestinal barrier‘s morphology and functionality
Several studies indicate that mycotoxins such as aflatoxin B1, DON, fumonisin B1, ochratoxin A, and T2, can increase the permeability of the intestinal epithelium of poultry and swine (e.g., Pinton & Oswald, 2014). This is primarily a consequence of the inhibition of protein synthesis.
As a result, there is an increase in the passage of antigens into the bloodstream (e.g., bacteria, viruses, and toxins). This increases the animal’s susceptibility to infectious enteric diseases. Moreover, the damage that mycotoxins cause to the intestinal barrier entails that they are also being absorbed at a higher rate.
2. Impaired immune function in the intestine
The intestine is a very active immune site, where several immuno-regulatory mechanisms simultaneously defend the body from harmful agents. Immune cells are affected by mycotoxins through the initiation of apoptosis, the inhibition or stimulation of cytokines, and the induction of oxidative stress.
It is known that fumonisin B1 may induce changes in the balance of sphingolipids at the cellular level, including for gastrointestinal cells. This facilitates the adhesion of pathogenic bacteria, increases in their populations, and prolongs infections, as has been shown in the case of E. coli. The colonization of the intestine of food-producing animals by pathogenic strains of E. coli and Salmonella also poses a risk to human health.
4. Interaction with bacterial toxins
When mycotoxins induce changes in the intestinal microbiota, this can increase the endotoxin concentration in the intestinal lumen. Endotoxins promote the release of several cytokines that induce an enhanced immune response, causing inflammation, thus reducing feed consumption and animal performance, damage to vital organs, sepsis, and death of the animals in some cases.
The synergy between mycotoxins and endotoxins can result in an overstimulation of the immune system. The interaction between endotoxins and estrogenic agents such as zearalenone, for example, generates chronic inflammation and autoimmune disorders because immune cells have estrogen receptors, which are stimulated by the mycotoxin.
Increased mycotoxin risks through by-products? Invest in mitigation solutions
To prevent the detrimental consequences of mycotoxins on animal health and performance, proactive solutions are needed that support the intestinal epithelium’s digestive and immune functionality and help maintain a balanced microbiome in the GIT. This becomes even more important as the current market conditions will likely engender a long-term shift towards including more cereal by-products in animal diets.
Trial data shows that EW Nutrition’s toxin-mitigating solution SOLIS MAX 2.0 provides adequate protection against feedborne mycotoxins. The synergistic combination of ingredients in SOLIS MAX 2.0 prevents mycotoxins from damaging the animals’ gastrointestinal tract and entering the bloodstream and additionally acts as antioxidant and liver-protecting:
Figure 3: Moa of Solis Max 2.0
In-vitro study shows strong mitigation effects of SOLIS MAX 2.0 against a wide range of mycotoxins
Animal feed is often contaminated with two or more mycotoxins, making it essential for an anti-mycotoxin agent to be effective against a wide range of different mycotoxins. A trial with SOLIS MAX 2.0 was conducted at an independent laboratory in Spain with an inclusion level of the product of 0.10% (equivalent to 1 kg per ton of feed). A phosphate buffer solution at pH 7 was prepared to simulate intestinal conditions in which a portion of the mycotoxins may be released from the binder (desorption). The following mycotoxins were evaluated in the test (see Table 1):
Table 1: Mycotoxin challenges
Each mycotoxin was tested separately by adding a challenge to buffer solutions, incubating for one hour at 41°C, to establish the baseline (table). At the same time, a solution with the toxin challenge and Solis Max 2.0 was prepared, incubated, and analyzed for the residual mycotoxin to find the binding efficacy. All analyses were carried out using high-performance liquid chromatography (HPLC) with standard detectors.
Figure 4: SOLIS MAX 2.0 (1 kg/t of feed) adsorption capacity against different mycotoxins (%)
The results (Figure 4) demonstrate that SOLIS MAX 2.0 is a highly effective solution against the most common mycotoxins in raw materials and animal feed.
Mycotoxin risk management for better animal feed
A healthy gastrointestinal tract is crucial to animals’ overall health: it ensures that nutrients are optimally absorbed, provides adequate protection against pathogens through its immune function, and is key to maintaining a well-balanced microflora. Even at levels considered safe by the European Union, mycotoxins can compromise different intestinal functions, resulting in lower productivity and susceptibility to disease.
The globalized feed trade, which spreads mycotoxins beyond their geographical origin, climate change, and raw material market pressures additionally escalate the problem. On top of rigorous testing, producers should mitigate unavoidable mycotoxin exposures by using solutions such as SOLIS MAX 2.0 – for stronger animal health, welfare, and productivity.
References
Antonissen, Gunther, An Martel, Frank Pasmans, Richard Ducatelle, Elin Verbrugghe, Virginie Vandenbroucke, Shaoji Li, Freddy Haesebrouck, Filip Van Immerseel, and Siska Croubels. “The Impact of Fusarium Mycotoxins on Human and Animal Host Susceptibility to Infectious Diseases.” Toxins 6, no. 2 (January 28, 2014): 430–52. https://doi.org/10.3390/toxins6020430.
Burel, Christine, Mael Tanguy, Philippe Guerre, Eric Boilletot, Roland Cariolet, Marilyne Queguiner, Gilbert Postollec, et al. “Effect of Low Dose of Fumonisins on Pig Health: Immune Status, Intestinal Microbiota and Sensitivity to Salmonella.” Toxins 5, no. 4 (April 23, 2013): 841–64. https://doi.org/10.3390/toxins5040841.
Burton, Emily J., Dawn V. Scholey, and Peter E. Williams. “Use of Cereal Crops for Food and Fuel – Characterization of a Novel Bioethanol Coproduct for Use in Meat Poultry Diets.” Food and Energy Security 2, no. 3 (September 19, 2013): 197–206. https://doi.org/10.1002/fes3.30.
Ghareeb, Khaled, Wageha A. Awad, Josef Böhm, and Qendrim Zebeli. “Impacts of the Feed Contaminant Deoxynivalenol on the Intestine of Monogastric Animals: Poultry and Swine.” Journal of Applied Toxicology 35, no. 4 (October 28, 2014): 327–37. https://doi.org/10.1002/jat.3083.
Mani, V., T. E. Weber, L. H. Baumgard, and N. K. Gabler. “Growth and Development Symposium: Endotoxin, Inflammation, and Intestinal Function in livestock1,2.” Journal of Animal Science 90, no. 5 (May 1, 2012): 1452–65. https://doi.org/10.2527/jas.2011-4627.
Obremski, K. “The Effect of in Vivo Exposure to Zearalenone on Cytokine Secretion by Th1 and Th2 Lymphocytes in Porcine Peyer’s Patches after in Vitro Stimulation with LPS.” Polish Journal of Veterinary Sciences 17, no. 4 (2014): 625–32. https://doi.org/10.2478/pjvs-2014-0093.
Oswald, I. P., C. Desautels, J. Laffitte, S. Fournout, S. Y. Peres, M. Odin, P. Le Bars, J. Le Bars, and J. M. Fairbrother. “Mycotoxin Fumonisin B1 Increases Intestinal Colonization by Pathogenic Escherichia Coli in Pigs.” Applied and Environmental Microbiology 69, no. 10 (2003): 5870–74. https://doi.org/10.1128/aem.69.10.5870-5874.2003.
Pinotti, Luciano, Matteo Ottoboni, Carlotta Giromini, Vittorio Dell’Orto, and Federica Cheli. “Mycotoxin Contamination in the EU Feed Supply Chain: A Focus on Cereal Byproducts.” Toxins 8, no. 2 (February 15, 2016): 45. https://doi.org/10.3390/toxins8020045.
Pinton, Philippe, and Isabelle Oswald. “Effect of Deoxynivalenol and Other Type B Trichothecenes on the Intestine: A Review.” Toxins 6, no. 5 (May 21, 2014): 1615–43. https://doi.org/10.3390/toxins6051615.
Mycotoxins pose a threat to the horse’s digestive system
Author: Judith Schmidt, Product Manager On Farm Solutions
Alarm in the gut! Horses have a susceptible digestive system that can quickly become unbalanced. Intestinal disorders in horses are usually associated with colic. Many factors can be responsible for intestinal issues. Have you ever thought about mycotoxins? What can horse owners do to support their horse´s gut health?
The equine stomach is not robust at all. Depending on their age and use, more than half of all horses suffer from stomach pain. Their digestive system is very sensitive and very different from that of other mammals: Horses cannot vomit and often suffer from severe abdominal pain, diarrhea, or cramps if they overeat or ingest spoiled feed.
The horse´s digestive system is complex and sensitive
The horse´s stomach has a relatively small capacity of around twelve to fifteen liters. Depending on the feed’s consistency and composition, it remains in the stomach for around one to five hours before it is pressed through the stomach outlet (pylorus) into the small intestine. The horse´s entire intestine is about ten times its body length.
Figure 1: The horse’s digestive tract
The horse´s gastrointestinal tract is a complex network, reacting extremely sensitively to changes and, therefore, highly susceptible to disorders. It essentially consists of the head intestine (lips, oral cavity, teeth, and esophagus), stomach (blind pouch, fundus, and stomach outlet), small intestine (duodenum, jejunum, and ileum), and large intestine (caecum, colon and rectum). Each section plays a crucial role in the digestive process; any disruption can lead to health issues. Understanding this structure is key to maintaining a horse’s digestive health.
Digestive disorders can have various reasons
Intestinal problems in horses can stem from diverse causes, often a complex interplay of multiple factors. By understanding these causes more deeply, horse owners can be better equipped to prevent and manage these issues. In the following, we delve into several of these causes.
1. Too long time between the feedings
Usually, a feeding break should be at most four to six hours, as, in nature, a horse is busy eating for at least 18 hours a day. In contrast to humans, who produce stomach acid only after food intake, the horse’s stomach produces gastric acid around the clock. The continuous intake of roughage, intensive chewing, and high saliva production (a horse produces 5 to 10 L of saliva per day) is, therefore, essential to protect the stomach mucosa by neutralizing excess gastric acid.
A too-long time between feedings and, therefore, no saliva production leads to an accumulation of gastric acid in the stomach. Four hours without roughage can already cause inflammation of the mucosa and probably ulcers.
2. Excessive amounts of concentrated feed
Excessive amounts of concentrates such as wheat or rye, conditioned by less chewing activity, increase gastric acid and histamine production, and the stomach lining can be attacked. Also, in this case, the development of stomach ulcers is possible.
Furthermore, the possibly resulting hyperacidity of the organism can lead to malfunctions of the organs, the skin, and the hooves.
3. Stress
Stress can also lead to a higher production of gastric acid and, therefore, to gastric ulcers. The horse is a flight animal. When it is under stress, it prepares for the impending escape, and the muscles are preferably supplied with blood, resulting in a lower blood flow to the mucous membranes. Furthermore, the rising cortisone level reduces the hydrochloric acid-suppressing prostaglandin E. As a result, more stomach acid is produced, irritating the gastric mucosa.
Stress can be triggered, e.g., by transportation, competitions, training, a change of house, a new rider, unsuitable equipment, or poor posture.
4. Dental diseases
The teeth are essential for digestion. When feed is chewed, it is broken down and mixed with saliva. Chipped teeth cannot chew well, and the feed is not sufficiently salivated or crushed, which has a detrimental effect on digestion.
For this reason, an expert vet should check the horse´s teeth at least once a year.
5. Administration of painkillers/medication
As with humans, long-term medication administration can promote the formation of stomach ulcers. For this reason, it is essential to ensure that horses are fed a gentle diet on the stomach, especially when using oral pain therapy, and to add stomach protection if necessary.
6. Endotoxins
If pathogens such as E. coli or clostridia proliferate extremely or are killed by an antibiotic, endotoxins can be released. These toxins can cause transformation or inflammation of the gut mucosa. In drastic cases, whole areas of the mucosa can die off.
7. Mycotoxins – the hidden danger in horse feed
Mycotoxins in plants and horse feed are a common but often unnoticed danger to horses’ health. Mycotoxins are natural, secondary metabolites of molds that have a toxic effect on humans and animals and can trigger mycotoxicosis. Contaminated feed can severely affect the horse’s health and, in the worst case, lead to death.
Over 90 % of the world´s feed production is estimated to be contaminated with at least one mycotoxin (see also Global Mycotoxin Report 2023, EW Nutrition. The intake of mycotoxins via hay, grain, silage, or compound feed can hardly be avoided. Mycotoxins are an increasing problem for all horse owners. Scientific studies show that the mycotoxins DON and ZEA are most frequently found in horse feed and, therefore, are also frequently detected in sports horses’ urine and blood samples.
Due to the highly toxic metabolic products, feed contaminated with molds can lead to severe liver and kidney diseases in horses, affect fertility, trigger colic, and promote digestive issues (diarrhea and watery stools).
Mycotoxins Horses
Figure 2: Mycotoxins and their impact on horses
How to protect the horse from mycotoxins?
The first measure against the ingestion of mycotoxins is prevention. Correct pasture management and adequate barn and feed hygiene can contribute to preventing the ingestion of toxins.
However, despite the best prophylactic measures, it is impossible to prevent mycotoxin contamination of feed completely. As mycotoxins are not visible, analyzing the feed regarding mycotoxin contamination is recommended.
To protect your horse from mycotoxins, EW Nutrition developed MasterRisk, a tool for evaluating the risk of mycotoxins. Additionally, EW Nutrition has developed a complementary feed specifically for your horse´s needs in the form of granules. The sophisticated formulation of “Toxi-Pearls” is designed to bind mycotoxins and mitigate the adverse effects of mycotoxin contamination.
The pearls contain a mixture of mycotoxin binder, brewer’s yeast, and herbs:
The contained mycotoxin binder effectively controls the most important feed myco- and endotoxins. It additionally supports the liver and immune system and strengthens the intestinal barrier.
Brewer´s yeast supports the natural strength of the gastrointestinal tract. Due to its high natural content of beta-glucans and mannan-oligosaccharides (MOS), unique surface structure, and the associated high adsorption power, brewer´s yeast has a prebiotic effect on the intestinal microbiome.
The additional unique herbal mixture consists of the typical gastrointestinal herbs oregano, rosemary, aniseed, fennel, and cinnamon. The processed beetroot is a true all-rounder. Literature shows that it has an antioxidant effect and strengthens the immune system. It also promotes bile secretion and, therefore, supports fat digestion.
Conclusion
The horse’s digestive tract is highly sensitive and must be supported by all means. Besides failures in management, such as too long breaks between feedings or too high amounts of feed concentrate, mycotoxins present a high risk in horse nutrition. To prevent horses from intestinal issues, feed and stress management, dental care, and medication in the case of disease must be optimized. Particular attention should be paid to possible mycotoxin contamination. Effective toxin risk management, which consists of analysis, risk evaluation, and adequate toxin risk-managing products, should be implemented.






")