How animal nutrition can contribute to sustainability
By Dr. Inge Heinzl, Editor, EW Nutrition
Nowadays, the whole world is talking about sustainability. Many efforts aim to maintain our world for future generations, creating a balance between our current needs and those of our children, grandchildren, and great-grandchildren. The right animal nutrition choices play a crucial role in achieving the challenging aim of sustainable animal production.
Animal nutrition solutions can support producers’ sustainability contributions, from animal welfare to antibiotic reduction
Sustainability – an old concept now set out in writing
The idea of sustainability is not new. Already the first humans lived sustainably, taking only as much as they needed and the environment could cope with, using all parts of the animals they killed. The German Hannss Carl von Carlowitz (1645-1714) coined the term sustainability in his oeuvre “Sylvicultura oeconomica” to counter a threatening raw material crisis. Wood was one of the most important raw materials. Besides heating, it was used for shipbuilding and mining. This was the reason that extensive areas in Europe were deforested and became deserted. Observing the impending disaster, von Carlowitz ” (1713) stated that only as many trees should be felled as can grow back through planned reforestation, sowing, and planting.
The Brundtland Report (1987), a document created by the World Commission on Environment and Development, is reckoned to be the starting signal for worldwide discussions about sustainability. In 2015, the result of a meeting of 193 members of the United Nations was the Agenda 2030 with 17 sustainable development goals for a “world we want” that should be achieved by 2030.
Sustainable Development Goals (SDG) of the Agenda 2030, fixed by the UN in 2015
How can the feed sector contribute to sustainability?
The animal nutrition industry’s sustainability efforts play into different SDGs, notably no. 2, zero hunger, no. 3, good health and well-being, no. 12, responsible consumption and production, no. 13, climate action, no. 14, life below water, and no. 15, life on land. In addition to the overarching goal of fostering higher animal welfare (cf. Keeling et al., 2019), the feed sector’s measures center on three areas:
Optimal use of feed resources, which includes optimizing feed conversion, preserving feed quality, and using alternative ingredients
Preserving the environment by reducing ammonia and methane emissions and energy requirements
Reducing antibiotics usage to maintain their efficacy for future generations
1. Make best use of available resources
One of the 17 points on the list of the United Nations is “responsible consumption and production”. For the feed industry, this means making the most out of available feed sources. Improvements in feed conversion, the maintenance of feed quality, and the use of alternative ingredients are all part of this.
Optimize FCR to utilize the available feed best
The feed conversion rate shows the amount of feed consumed in relation to the outputs produced, such as weight gain, eggs, or milk. The better or lower the feed conversion rate (FCR), the less feed you need to achieve your target, and the higher the yield. Products that improve feed conversion, therefore, can help to save resources.
Good feed conversion or an optimal utilization of nutrients depends on gut health. Only a healthy gut can digest the feed and absorb the nutrients adequately. Hence, products to improve feed conversion often do so by improving gut health.
Phytomolecules: proven to improve feed conversion
Herbs and their active components have been used in human and veterinary medicine for thousands of years to treat digestive tract diseases. Nowadays, products based on phytomolecules help improve feed conversion through their digestive, anti-inflammatory, and antimicrobial effects on the intestinal tract.
How do these three characteristics contribute to a better FCR?
Phytomolecules stimulate the secretion of digestive juices and the motility of the gut
Their antimicrobial effect supports a “healthy” balance in the microbiome, preventing damages of the gut wall by harmful microbes and, therefore, maintaining an optimal nutrient absorption
Their anti-inflammatory properties also contribute to good nutrient absorption and reduce endogenous nutrient loss
FCR improvements in broilers thanks to ACTIVO found in several studies
As phytomolecules are often volatile, EW Nutrition offers encapsulated phytomolecule-based products for the feed (ACTIVO product line). During episodes of elevated enteric challenge, e.g., weaning or following feed change, a liquid solution (ACTIVO LIQUID) can be applied via the waterline.
Enzymes help to make nutrients available
Some feed materials are hard to digest for certain animals. For example, pigs’ digestive systems do not have the enzymes required to break down non-starch polysaccharides (NSPs), such as cellulose, hemicellulose (ß-glucans and xylans), pectins or oligosaccharides. However, pig feed ingredients usually contain these substances.
Besides the non-usability of NSPs, the cage effect is a further problem. Cellulose and hemicellulose, water-insoluble NSPs, encage nutrients such as proteins or digestible carbohydrates. Encaged nutrients cannot be reached by the digestive enzymes and don’t become available to the animal.
Xylanases are available on the market to degrade structural substances in the feed and make them, as well as the nutrients they encaged, available for the organism.
Maintain the quality of your feed materials
Another possibility to save resources is the maintenance of feed quality. Bad weather conditions at harvest or incorrect storage can downgrade feed quality due to the development of molds and their mycotoxins or the oxidation of nutrients. Products mitigating the adverse effects of toxins, acidifiers that reduce microbial load, and antioxidants can help to keep your feed quality on a high level – or to re-establish it.
Mitigate the adverse effects of mycotoxins
Feed materials contaminated with mycotoxins harm animals in different manners and should not be used without further treatment. Mycotoxins are not visible – even if no molds are visible, mycotoxins might be present. Additionally, they are pH- and thermo-stable, meaning that mycotoxins produced in the raw materials on the field remain in the finished feed. As mycotoxins often do not cause apparent, specific symptoms but manifest in decreased performance, feed refusal or lower feed intake, and higher disease susceptibility, it is difficult to notice contamination.
Products such as SOLIS or MASTERSORB contain clay minerals (bentonite and montmorillonite) that adsorb the toxins. MASTERSORB GOLD and MASTERSORB FM also include toxin-adsorbing yeast cell walls and herbal substances to help protect the liver.
Feed spoilage through molds, yeasts, and mycotoxins wastes precious resources
Reduce microbes in the feed with acidifiers
Acidifiers based on organic acids counter harmful microbes in the feed in two ways. Most pathogenic bacteria are susceptible to low pH. The proliferation of, e.g., E. coli, Salmonella, and Clostridium perfringens is minimized at pH < 5 (cf. Fuller 1977). Acidic-tolerant beneficial bacteria such as Lactobacilli or Bifidobacterium, however, survive.
Other than antimicrobial activity, organic acids also cause a significant reduction in ammonia (Eriksen et al., 2014). This finding could be due to a reduction in the microbial deamination of amino acids, which would then be available for absorption, resulting in increased nitrogen digestibility and reduced ammonia excretion, as observed in monogastrics fed organic acids (Pearlin et al., 2020).
The acidifier product lines ACIDOMIX, FORMYCINE, and PRO-STABIL all help protect feed from contamination with pathogenic microorganisms.
Protect the feed’s nutrients from oxidation
The oxidation of nutrients in the feed decreases its nutritional value and, thereby, the value of the whole diet. Fat, proteins, fat-soluble vitamins, pigments, and other biologically active molecules, including sugars and phospholipids, can get oxidized. Metal ions and other pro-oxidative factors can affect the ingredients of the feed during mixing, storage, and feeding. The oxidation of fats and fat-soluble vitamins results in color changes or odors and – this is even more serious – in the production of harmful substances such as aldehydes and ketones. An oxidized feed can lead to oxidative stress in the animals, reduce their immunity, productivity, and livability.
To protect valuable ingredients, the timely addition of effective antioxidants such as STABILON is recommended.
Use alternatives to natural protein sources
Soybeans are an excellent source of protein in animal nutrition. During the last 50 years, soy production has increased from 27 million tons to 269 million tons, causing environmental degradation of forests and savannas (WWF, 2021). The use of alternative protein sources helps protect our environment.
Ruminants partly cover their protein requirements with the help of rumen bacteria. These bacteria can turn nitrogen from urea into bacterial protein, provided they receive enough energy available from carbohydrates. Thanks to its encapsulation, PROTE-N, a feed-grade urea-based nitrogen source, slowly releases nitrogen into the rumen, synchronized with the energy supply. PROTE-N affords producers a degree of independence from soybean protein without compromising nutritional quality.
Reducing soybeans in ruminant feeds helps to lower their environmental impact
2. Preserve the environment
Animal production generates gases such as ammonia and methane that negatively impact the environment. Measures to reduce these gases help to protect plants, animals, us, and our globe.
Reduce ammonia by improving protein digestion
Besides nitrogen oxides, ammonia is one of the primary sources of nitrogen pollution. Ammonia damages ecological systems through acidification and nutritional oversupply. Fast-growing plants that need high amounts of nitrogen or plants that tolerate low soil pH proliferate, whereas more susceptible plants disappear, decreasing biodiversity. According to Max-Planck-Gesellschaft (2017), reducing ammonia emissions by 50 % could prevent 250.000 deaths caused by fine dust worldwide per year.
Improved protein digestion in animals reduces their ammonia production. Decreasing the intestinal pH through using organic acid-based products such as ACIDOMIX or FORMYCINE is essential for the activation and correct functioning of the enzymes responsible for protein digestion.
Reduce methane, the second most abundant greenhouse gas
Together with CO2, N2O, and three fluorinated gases, methane belongs to the greenhouse gases listed in the Kyoto protocol. Being over 25 times more potent than carbon dioxide at trapping heat in the atmosphere, it dramatically affects the earth’s temperature and the climate system (United States Environmental Protection Agency). Methane is a final product of feed fermentation in the rumen and is produced by methanogenic bacteria. Ruminants can produce 250-500 L methane per day (Johnson & Johnson, 1995).
Reducing methane production in ruminants is a critical step towards climate protection. Herbal substances can change the microbiome, leading to improved protein and fiber degradation and reduced methane production (Ku-Vera et al., 2020). ACTIVO PREMIUM is a phytomolecules-based product for ruminants that helps reduce their methane emissions.
Energy savings
To preserve the environment, reducing energy needs is also an important topic. Using the surfactant SURF-ACE in the pelletizing process, feed mills can cut 10-15 % of their energy consumption or produce up to 10-15 % higher pellet output without increasing their energy consumption. When moisture is added together with the surfactant, the emulsion of the dietary fat and the added water leads to better general lubrication of the machinery and improved press throughput.
Feed mill efficiency is key to animal nutrition’s carbon footprint
3. Reduce antibiotic use in animal production to keep this tool effective
Point 3 on the UN’s Agenda 2030 is good health and well-being. For many years, antibiotics, a very effective weapon, have been used to fight bacterial diseases. However, the occurrence of resistance is increasing. One of the reasons is the inappropriate use of antibiotics. These substances are often used preventively or for viral diseases against which they are ineffective. Also, the use of antibiotics as growth promoters at low dosages in animal production strongly contributed to the development of antimicrobial resistance.
Limiting antibiotic use to therapeutic treatment is possible through good farm management and feed supplements that support animals’ gut health, immune systems, and respiratory health. For this purpose, solutions ranging from phytomolecules (ACTIVO products, GRIPPOZON) to egg immunoglobulins (GLOBIGEN products, PROTEGG), products mitigating the impact of toxins (MASTERSORB products, SOLIS), beta-glucans/MOS (BGMOS), and acidifiers (ACIDOMIX, FORMYCINE) are available.
The feed sector has the tools to achieve more sustainability!
The animal nutrition industry provides many products to support animal producers in coping with their main challenges, including the shift to more sustainable production practices. Solutions exist to save feed resources, better protect the environment, and keep antibiotic tools effective. As an additional reward, implementing sustainability solutions leads to healthy animals with high performance. Let’s all help to preserve this planet for our next generations!
References
Eriksen, J., Nørgaard, J. V., Poulsen, H. D., Poulsen, H. V., Jensen, B. B., & Petersen, S. O. (2014). Effects of Acidifying Pig diets on emissions of AMMONIA, methane, and sulfur FROM Slurry during storage. Journal of Environmental Quality, 43(6), 2086–2095. https://doi.org/10.2134/jeq2014.03.0108
Fuller, R. (1977). The importance of lactobacilli in maintaining normal microbial balance in the crop. British Poultry Science, 18(1), 85–94. https://doi.org/10.1080/00071667708416332
Johnson, K. A., & Johnson, D. E. (1995). Methane emissions from cattle. Journal of Animal Science, 73(8), 2483–2492. https://doi.org/10.2527/1995.7382483x
Keeling, Linda, Håkan Tunón, Gabriela Olmos Antillón, Charlotte Berg, Mike Jones, Leopoldo Stuardo, Janice Swanson, Anna Wallenbeck, Christoph Winckler, and Harry Blokhuis. “Animal Welfare and the United Nations Sustainable Development Goals.” Frontiers in Veterinary Science 6 (October 10, 2019). https://doi.org/10.3389/fvets.2019.00336.
Ku-Vera, J. C., Jiménez-Ocampo, R., Valencia-Salazar, S. S., Montoya-Flores, M. D., Molina-Botero, I. C., Arango, J., Gómez-Bravo, C. A., Aguilar-Pérez, C. F., & Solorio-Sánchez, F. J. (2020). Role of secondary plant metabolites on enteric methane mitigation in ruminants. Frontiers in Veterinary Science, 7. https://doi.org/10.3389/fvets.2020.00584
Pearlin, B. V., Muthuvel, S., Govidasamy, P., Villavan, M., Alagawany, M., Ragab Farag, M., Dhama, K., & Gopi, M. (2020). Role of acidifiers in livestock nutrition and health: A review. Journal of Animal Physiology and Animal Nutrition, 104(2), 558–569. https://doi.org/10.1111/jpn.13282
United Nations. (n.d.). How your company can advance each of THE SDGS: UN Global Compact. How Your Company Can Advance Each of the SDGs | UN Global Compact. https://www.unglobalcompact.org/sdgs/17-global-goals.
von Carlowitz, H. C. (1713). Sylvicvltvra oeconomica, oder, Hausswirthliche Nachricht und Naturmässige Anweisung zur Wilden BAŬM-ZŬCHT: Nebst gründlicher darstellung, wie Zu FÖRDERST durch Göttliches Benedeyen Dem allenthalben und insgemein einreissenden Grossen Holtz-mangel: Vermittelst Säe-pflantz- und Versetzung Vielerhand Bäume zu prospiciren …: Worbey zugleich eine Gründliche nachricht von den in Churfl. Sächss. Landen gefundenen Turff Dessen Naturliche beschaffenheit, Grossen NÜTZEN, Gebrauch und nutzlichen verkohlung, Aus Liebe Zu BEFÖRDERUNG des Algemeinen Bestens beschrieben. Verlegts Johann Friedrich Braun.
Antibiotics are a precious resource whose long-term efficacy must be protected – for human and animal health. This is a difficult challenge for healthcare providers (veterinarians and medical doctors), as well as farmers and integrators. In this series of articles, we will explore the general and specific measures and solutions through which animal production can contribute to the overall reduction of antibiotic use.
Responsible animal production contributes to maintaining antibiotic efficacy
Shortly after the discovery of penicillin in 1929, Alexander Fleming already pointed out the possibility of resistance during an interview with the New York Times. The first case of penicillin resistance was reported only one year after clinical trials began; within 20 years, 80% of Staphylococcus aureus isolates were resistant to penicillin (Lobanovska and Pilla, 2017).
Over the years, clients and patients have gotten used to receiving a pill to quickly fix their ailments. Often, antibiotics have been prescribed for illnesses they were not effective against, including viral challenges. This has unnecessarily accelerated the rate of resistance development. To reverse this trend, education is key. At the same time, the judicious use of antibiotics, meaning the correct antibiotic for the challenge plus proper administration and duration of use, is paramount for all medical professionals to help preserve the efficacy of these critical substances.
Antibiotic use in animal production must be reduced
For many years, animal producers have used antibiotics as growth promoters. The E.U. banned this type of use in 2006, and the United States followed in 2017. Evaluations have shown a decrease in antibiotic use in the U.S.: In 2014, according to the FDA, 17,000 tons of antibiotics were sold in the United States for livestock, representing 80 percent of all U.S. antibiotics sales. In 2019, a total of about 11,000 tons of antibiotics were sold for use in food-producing animals (FDA, 2020).
As the number of isolated multi-drug resistant bacteria increases and the discovery and approval of new antibiotics slows, it is imperative that the use of antibiotics in animal production, especially those that are critically important for humans, is reduced to a minimum. Hence, antibiotics should only be used to treat, control, or prevent diseases in case of imminent risk, but not for growth-promoting purposes.
Scanning electron micrograph of methicillin-resistant Staphylococcus aureus bacteria (yellow) and a dead human white blood cell (red). Credit: National Institute of Allergy and Infectious Diseases/NIH
Customers’ requests for antibiotic-free chicken push antibiotic reduction
Many birds are already raised without antibiotics in the US and elsewhere because of the demands of the market. Since 2016, chicken antibiotic sales decreased by 62% (Dall, 2020). Frequently, the goal of these antibiotic-free (ABF) production programs is to differentiate products in a highly competitive commodity market. The reduction of antibiotic use has been a secondary, generally unintended consequence.
Nevertheless, to meet customer demands for ABF products, antibiotics that are not important to human health but for production (e.g., ionophores) have also been eliminated. In many cases, this has negatively affected growth performance and bird health. As the requirements for production efficiency and welfare standards increase, transitioning from “conventional” to ABF production poses a challenge for everyone involved.
Antibiotic reduction through improved management
One must never trade animal welfare for reduced antibiotics use, but the need for them can be decreased through improved management practices. Flock health starts with genetics companies selecting birds that are resilient to disease and management challenges and continues all the way to the processing plant. All of the inputs and practices must be optimized in modern poultry production to maintain a high level of performance and animal welfare while reducing reliance on antibiotics.
Antibiotic-free requires diligent management
When antibiotics are not available, attention to detail becomes more decisive. All aspects of production are important, but the most critical stages are those that affect the downstream process. The pullets, breeders, and hatchery require the most meticulous care. Additionally, all production factors must meet the highest quality standards: feed, light, air quality, water quality, litter quality, biosecurity, vaccination, sanitation, nutrition and feeding.
Antibiotic reduction requires meticulous attention to detail to safeguard animal welfare.
Non-antibiotic feed additives support ABF programs
ABF production is all about sustainability. For agricultural operations to survive and thrive in the future, one has to move away from the old paradigm of “saving the way to success”. This is not impossible in ABF production, but misses out on the larger picture of long-term profitability, investment in innovation, and system change.
Non-antibiotic feed and water additives are essential resources to support sustainable management. To mention a few, probiotics, prebiotics, toxin binders, organic acids, and phytomolecules are all options for reducing the need for antibiotics based on different modes of action. Phytomolecules, for example, often have antimicrobial properties, some toxin binders can bind bacterial toxins, and pre- and probiotics support the gut flora. There are many kinds of solutions on the market; the key is to find the right ones for your issues.
Antibiotic stewardship: together for a healthier future
There is already a large body of literature demonstrating the benefits of alternative or complementary solutions. More importantly, there are already many people that successfully raise birds and other animals without antibiotics. Whenever possible, leverage your professional network and talk to trusted people with unique experiences. Working together, we can build a healthier future for people and animals.
The Antibiotic Reduction series
The series that debuts here consists of a set of articles offering professionals a practical overview of poultry production with reduced antibiotic use. The independent expert in charge, starting with the next article in the series, is Dr. TJ Gaydos, who holds a Master’s degree in Avian Medicine and is a diplomate of the American College of Poultry Veterinarians.
Dr. Gaydos works with integrated poultry companies and allied industries, focusing on bird health and antibiotic-free production performance. He has spent his veterinary career working to improve intestinal health, animal welfare, production efficiency, and reduce zoonotic diseases. He works extensively with intestinal health, probiotics and prebiotics, and other non-antimicrobial feed additives.
Topics covered under Dr. Gaydos’s guidance include biosecurity, nutrition, pullet management, hatchery sanitation, gut health, and more. Together they provide an extensive look at the producers’ pain points and potential strategies to maintain bird health while mitigating the need for antibiotics.
Lobanovska, Mariya, and Giulia Pilla . “Penicillin’s Discovery and Antibiotic Resistance: Lessons for the Future?” Yale Journal of Biology and Medicine. 90, no. 1 (March 29, 2017): 135–45.
U.S. Food and Drug Administration. “2020 Summary Report On Antimicrobials Sold or Distributed for Use in Food-Producing Animals” Food and Drug Administration, 2019. https://www.fda.gov/media/144427/download.
Norovirus outbreaks could be avoided with IgY technology
By Technical Team, EW Nutrition
In July, Public Health England (PHE), an executive agency of the Department of Health and Social Care in the United Kingdom, reported a rise of norovirus outbreaks in the country. Norovirus, a highly contagious virus similar to the coronavirus, is the main cause of viral food poisoning from shellfish. Symptoms include vomiting, diarrhea, cramps, as well as muscle aches and headaches.
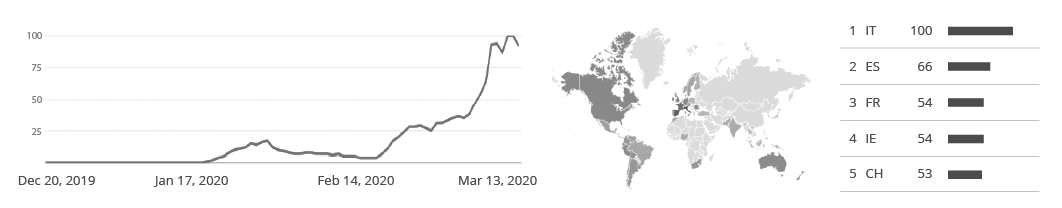
The PHE press release shows an increase in outbreaks during the last two months, returning to pre-pandemic levels. According to the organization, the number of outbreaks has nearly tripled when compared to the same time period in the last 5 years, affecting people of all age groups and settings in England. Closed places where the virus can spread quickly, especially childcare facilities and nursing homes, are the most affected, as shown below.
The issue is not restricted to England. According to the CDC (Centers for Disease Control and Prevention), about one out of every five cases of acute gastroenteritis that leads to diarrhea and vomiting is caused by norovirus, responsible for over 200,000 deaths and a global economic burden of more than $60 billion. The large costs come from healthcare costs and productivity losses and can be seen in low, middle, and high-income countries as shown below.
Global economic burden of norovirus gastroenteritis. Source: https://doi.org/10.1371/journal.pone.0151219.t003
Prevention is key
Noroviruses is easily transmitted through contact with infected individuals or contaminated surfaces. There are many ways to reduce the spread of the virus (e.g., washing the hand thoroughly with soap and water) but prevention is key.
The outbreaks often occur from contaminated oysters or other shellfish which are consumed raw, making foodborne transmission accountable for a considerable number of cases. The conventional cleaning and purifying methods currently used in the industry cannot reliably reduce the number of norovirus contained in its digestive organ, therefore it is of extreme necessity to look new solutions to improve safety in shellfish production. And this is exactly what EW Nutrition does.
Combatting the norovirus: the IgY solution
With our mission to mitigate the impact of antimicrobial resistance in mind, we developed a new technology to improve food safety in shellfish production. Our solution is based on a high value source of natural egg immunoglobulins (IgY), which will prevent the virus from infecting the oyster’s digestive organ.
This method consists in adding anti-norovirus IgY to the seawater during the depuration process, which is a postharvest treatment where the shellfish are placed in tanks of clean seawater to reduce contaminant levels and allow shellfish to cleanse or purge themselves by continuation of their normal filter-feeding and digestive processes.
Natural, effective, and safe
While depuration is a highly effective and very common commercial practice for removing different pathogens, several studies show that the depuration process alone is not enough to remove completely or lower the norovirus to a safe level. On the other hand, various trial results show that shellfish treated with EW Nutrition technology is completely free from or has very low amount of live norovirus, allowing a safe consumption of raw oysters and minimizing the risk of any other outbreaks.
For more information about our solution, you can reach out to Lucas Queiroz or to your local EW Nutrition contact.
The Quick Take: Mitigate spikes in Vitamin E costs
Vitamin E prices have spiked amid production issues and lack of availability. How can you mitigate the increased cost of vitamin E inclusion?
Vitamin E prices often see severe fluctuations caused by raw materials shortages, production or distribution issues, or regulations on some key production ingredients (such as m-cresol anti-dumping rules in China leading to a global price spike some months ago).
SANTOQUIN acts as a preservative for Vitamin E, allowing more of this vitamin to enter the tissue where it exerts its antioxidant effect. In addition, in the presence of selenium, another important cellular antioxidant mineral, SANTOQUIN can help protect or spare the Vitamin E needed for proper cell function.
The Food and Agriculture Organization of the United Nations, FAO, clearly confirms this mode of action: “Dietary deficiencies of vitamins A and E seem to be ameliorated in certain circumstances and ethoxyquin promotes higher levels of vitamin A storage in the liver. Repletion/deletion experiments show that in both monogastric and ruminant animals, a diet containing an anti-oxidant protects fat soluble vitamins throughout ingestion and metabolism. The important benefit of antioxidants most probably lies in their conservation of essential nutrients and their improved utilization by the animal. Altogether too often, it is the practice to use levels of vitamin E far above the animals’ nutrient requirement and the result is economically unfavorable. It has been shown in diets designed for chicken and turkey breeders that ethoxyquin has a vitamin E sparing effect.”
Beyond AGPs: Controlling necrotic enteritis through gut health optimization
Antibiotic growth promoters (AGPs) have routinely been used in intensive poultry production for improving birds’ performance. However, in recent years, reducing the use of antibiotics in animal production has become a top priority, due to concerns about the development of antibiotic-resistant bacteria and mounting consumer pressure. Multiple countries have introduced bans or severe restrictions on the non-therapeutic use of antibiotics, including in the US, where the Food and Drug Administration has implemented measures to curb the use of antibiotics since 2017.
However, the removal of AGPs poses challenges for poultry performance, including reduced feed efficiency, decreased daily weight gain, as well as higher mortality. Moreover, the withdrawal of AGPs in feed is widely recognized as one of the predisposing factors for necrotic enteritis (NE). NE is one of the most common and economically important poultry diseases, with an estimated global impact of US$ 5 to 6 billion per year. As a result of withdrawing AGPs, the usage of therapeutic antibiotics to treat NE has increased. To break out of this vicious cycle and to secure the efficiency of poultry production, alternatives are needed that combat NE where it starts: in the gut.
Necrotic enteritis: a complex disease
NE is caused by pathogenic strains of Clostridium perfringens (CP): ubiquitous, gram-positive, spore-forming anaerobic bacteria. The spores of CP can be found in poultry litter, feces, soil, dust, and contaminated feed. Low levels of different CP strains are naturally present in the intestines of healthy birds, kept in check by a balanced microbiome. However, when gut health is compromised, pathogenic strains can proliferate at the expense of unproblematic strains, resulting in clinical or sub-clinical NE.
Animals suffering from the clinical form show symptoms such as general depression, reluctance to move, and diarrhea, with mortality rates of up to 50%. Infected birds suffer from degenerated mucosa lesions in the small intestines. Even in its “mild”, subclinical form, which often goes unnoticed, the damage to the animals’ intestinal mucosa can result in permanently reduced performance and consequent economic losses for the producer.
Certain predisposing factors have been found to enable the proliferation of pathogenic strains in the gastrointestinal tract. Diet is a key example: the composition of the gut flora is directly linked to feed composition. High inclusion rates of cereals (barley, rye, oats, and wheat) that contain high levels of non-starch polysaccharides (NSPs), high levels of indigestible protein, and inclusion of proteins of animal origin (e.g. fishmeal) have been shown to predispose birds to NE.
A range of diseases (e.g. chicken infectious anemia, Gumboro, and Marek’s disease), but also other factors that have immunosuppressive effects, such as heat or cold stress, mycotoxins, feed changes, or high stocking density, render birds more susceptible to intestinal infections. The single most prominent predisposing factor for the occurrence of NE is the mucosal damage caused by coccidiosis.
Gut health is key to combating necrotic enteritis
To control NE, a holistic approach to optimizing the intestinal health of poultry is needed. It should take into account not only parameters such as diet, hygiene, and stress, but should also make use of innovative tools.
Phytomolecules, also known as secondary plant compounds, are essentially plants’ defense mechanisms against pathogens such as moulds, yeasts, and bacteria. Studies have demonstrated the antimicrobial effects of certain phytomolecules, including against antibiotic-resistant pathogens. Phytomolecules have also been found to boost the production of digestive enzymes, to suppress pro-inflammatory prostaglandins and have antioxidant properties. These features make them a potent tool for optimizing gut health, potentially to the point of replacing AGPs.
Can phytomolecules mitigate the impact of necrotic enteritis?
To study the impact of phytomolecules on the performance of broilers challenged with a NE-causing CP strain, a trial was conducted at a US-based research facility. In this 42-day study, 1050 male day-old Cobb 500 broiler chicks were divided into 3 groups, with 7 replicates of 50 chicks each.
On the first day, all animals were vaccinated against coccidiosis through a live oocyst spray vaccination. The experimental diets met or exceeded the National Research Council requirements, and were fed as crumbles/pellets. On days 19, 20, and 21, all pens, except the negative control group, were challenged with a broth culture of C. perfringens. A field isolate of CP known to cause NE (originating from a commercial broiler operation) was utilized as the challenge organism. On day 21, three birds from each pen were selected, sacrificed, group weighed, and examined for the degree of present NE lesions.
The positive control group received no supplements. The trial group received a synergistic combination of two phytogenic products containing standardized amounts of selected, microencapsulated phytomolecules: an in-feed phytogenic premix (Activo®, EW Nutrition GmbH) and a liquid complementary feed supplied via the drinking water (Activo® Liquid, EW Nutrition GmbH). The products were given at inclusion rates corresponding to the manufacturer’s baseline antibiotic reduction program recommendations (Figure 1):
Figure 1: Trial design
The trial results indicate that the addition of phytomolecules helps to mitigate the impact of NE on broilers’ performance. The group receiving Activo® and Activo® Liquid showed a better feed conversion (Figure 2) compared to the positive control group (NE challenge, no supplement). Also, better lesion scores were noted for animals receiving phytomolecules (0.7 and 1) than for the positive control group (1.6).
The most significant effect was observed concerning mortality: the group receiving Activo® and Activo® Liquid showed a 50% lower mortality rate than the positive control group (Figure 3). These results clearly indicate that phytomolecules can play an important role in mitigating losses due to NE.
Figure 1: Adjusted FCR
Figure 2: Lesion scores and mortality
Tackling necrotic enteritis in a sustainable way
In an age of AGP-free poultry production, a concerted focus on fostering animals’ gut health is key to achieving optimal performance. This study strongly demonstrates that, thanks to their antimicrobial, digestive, anti-inflammatory and antioxidant properties, phytomolecules effectively support birds’ intestinal health when challenged with NE. The inclusion of Activo® and Activo® Liquid, two phytogenic products designed to synergistically support birds during critical periods, resulted in improved feed conversion, better lesion scores, and 50% lower mortality.
In combination with good dietary, hygiene, and management practices, phytomolecules are therefore a potent tool for reducing the use of antibiotics: including Activo® and Activo® Liquid in their animals’ diets allows poultry producers to reduce the incidence of NE, to mitigate its economic impact in case of outbreaks, and therefore to control NE in a sustainable way.
Tang, Karen L., Niamh P. Caffrey, Diego B. Nóbrega, Susan C. Cork, Paul E. Ronksley, Herman W. Barkema, Alicia J. Polachek, Heather Ganshorn, Nishan Sharma, James D. Kellner, and William A. Ghali. “Restricting the Use of Antibiotics in Food-producing Animals and Its Associations with Antibiotic Resistance in Food-producing Animals and Human Beings: A Systematic Review and Meta-analysis.” The Lancet Planetary Health 1, no. 8 (November 6, 2017): 316-27. doi:10.1016/s2542-5196(17)30141-9.
Van Immerseel, Filip, Julian I. Rood, Robert J. Moore, and Richard W. Titball. “Rethinking Our Understanding of the Pathogenesis of Necrotic Enteritis in Chickens.” Trends in Microbiology 17, no. 1 (2009): 32-36. doi:10.1016/j.tim.2008.09.005.
Understanding the dangers of mycotoxins for breeder hens
As the producers of hatching eggs and day-old chicks, breeding hens are the backbone of the poultry industry. Hence it is common practice to pay particular attention to this valuable asset’s feed, selecting raw materials of high nutritional quality and safety. However, in any feed formulated for animals in production and reproduction, studies show that it is almost inevitable to find a certain level of mycotoxin contamination.
Mycotoxins exert toxic effects mainly on the gastrointestinal tract, liver, and kidneys and can accumulate in some tissues but also in the eggs. Mycotoxin contamination in breeder hens rations does not always lead to visible symptoms, such as when trichothecenes cause oral lesions. However, it may influence productivity, egg quality, hatchery performance, as well as chick quality and immunity. Mycotoxin risk management is thus an essential part of managing breeder hens. Mycotoxins can negatively affect eggshell quality and, as a consequence, embryonic mortality.
Type of mycotoxin and exposure time determine effect on egg production
Mycotoxicosis in hens can cause reduced egg production, most likely because it causes a decrease in protein synthesis. A lower synthesis of albumin results from a degeneration of the liver tissue due to aflatoxin, ochratoxin, T2 and DON exposure. The liver then may look pale, friable and occasionally shows superficial hemorrhages.
The contamination levels at which these effects can be observed are as low as 100ppb in feed, for example, during a 21-day exposure to ochratoxin (Figure 1). With increasing levels of the toxin, production further decreases. A similar effect is observed when breeder hens are exposed to aflatoxins.
Figure 1 – Effect of mycotoxins on egg production, compared to non-contaminated control (=100 %)
Egg production, however, is not the only parameter that is affected when breeding hens are exposed to mycotoxins. Earlier on in the reproductive cycle, they already impact on embryonic mortality and hatchability. These effects are potentially more severe and may even occur without any noticeable change in the number of eggs produced.
Mycotoxins’ insidious consequences for eggshell quality and embryonic mortality
The eggshell is important to protect the progeny: thin and fragile shells can increase embryonic mortality, lower embryonic weight gain and decrease hatchability. Eggshell quality is a function of the hen’s calcium and vitamin D3 metabolism. The bioavailability of calcium and of vitamin D3 depends on intestinal integrity and on the production of enzymes and transporters that aid in feed metabolism. These processes can be adversely affected by aflatoxins, DON, T2, and Fumonisins.
The gastrointestinal tract is not the only site of mycotoxin action, however. Mycotoxins such as aflatoxins and ochratoxins have nephrotoxic effects, affecting calcium metabolism and increasing its excretion via the urine, while lowering its levels in blood serum.
Moreover, mycotoxins damage the liver, which plays a central role in egg production, being responsible for vitamin D3 metabolism and the synthesis of the lipids that make up the yolk. Moreover, the synthesis of transporters for lipids, calcium, and carotenoids ̶ important components of the egg ̶ also takes place in the liver. When liver function is impaired, the internal and external quality of the egg declines, which, in the end, affects the production of day-old chicks.
Figure 2 – Effects of mycotoxins on eggshell quality and embryonic mortality
Figure 2 summarises the possible ways in which mycotoxins can negatively affect eggshell quality and, as a consequence, increase embryonic mortality. If a hen’s intestinal integrity is compromised, the utilization of nutrients decreases. Liver and kidney damage leads to a diminished availability of calcium and other nutrients necessary for egg formation. The birds’ calcium (and phosphorus) levels in the plasma are then lower and may lead to a greater mobilization of calcium from the bones. However, this response cannot be maintained and the eggs get a thinner shell.
The thickness of the eggshell influences the egg’s moisture loss and exchange with the environment during the incubation period. An eggshell of optimal quality does not allow the loss of nutrients and prevents bacterial contamination. Thinner eggshells are less able to fulfill these functions, leading to higher embryo mortality.
Figure 3 – Effects of mycotoxins on embryonic mortality
Figure 3 shows the effect of different mycotoxins on embryonic mortality. Incremental levels of ochratoxin or aflatoxin heighten embryonic mortality in a range from 1.5 to 7.5 times the embryonic mortality of the control group. In some cases, embryos are affected even when the hens received feed contaminated with mycotoxin levels that are within the guidelines suggested by the EFSA.
For example, an exposure to 4900ppb of DON for ten weeks increases the number of embryos with abnormalities. The causes are not entirely clear, as only traces of DON can be found in the egg. However, we do know that this mycotoxin can affect the protein synthesis at the level of the hen’s liver and therefore compromise the deposition of nutrients into the egg.
Mycotoxins’ effects on the progeny may cause long-term damage
Ochratoxin and aflatoxin can be transferred into the egg, where they exert toxicity on the embryos. This does not necessarily result in mortality. However, the chicks can suffer from a compromised immune function due to two reasons: lower transmission of antibodies from the hen and lower viability of the chickens’ immune cells, accompanied by a lower relative weight of the bursa of Fabricio and the thymus.
When both aflatoxin and ochratoxin are present in the feed, the effect on these parameters is synergistic. As a consequence of mycotoxin contamination, the animals’ immune response is impaired, which makes them more susceptible to infection. The final result could be increased early chick mortality due to a higher incidence of bacterial and viral infections.
The transmission of other mycotoxins into the egg is minimal. While this means that a direct effect on the progeny is unlikely to occur, mycotoxin contamination still has a snowball effect: we have to consider the indirect effect of a lower deposition of nutrients on chick quality.
Prevention is key: mycotoxin risk management for breeder hens
The best approach to manage mycotoxin risk is to implement an integrated strategy that includes good crop and grain storing practices, regular raw material sampling and mycotoxin evaluation and analysis. Management tools (such as MasterRisk) can help to evaluate mycotoxin interactions and to choose the best strategy for dealing with specific mycotoxin challenges.
The results of mycotoxin analyses can be used to take decisions regarding the inclusion levels of raw materials and in choosing feed additives that counteract mycotoxins. Products based on plant extracts, yeast cell walls, and clay minerals can help to stabilize a digestive system challenged by mycotoxins. They support the barrier function in the intestine, preventing the passage of mycotoxins into the bloodstream.
Phytomolecules are another piece of the puzzle: thanks to their antimicrobial, anti-inflammatory and antioxidant properties, they support liver function. This is particularly important for long-living animals prone to accumulating mycotoxins in their body tissues.
For a long time the “deleterious effects” of mycotoxins on breeder hens and “their repercussions on progeny health status and performance have not received from a scientific point of view as much attention”(Calini and Sirri, 2007) as they ought to have. However, now that the dangers of mycotoxins for breeder hens’ welfare, health and performance are better understood, it is clear that mycotoxin risk evaluation and management is central to successful poultry production.
*This article first appeared in All About Feed on 31 October 2018
byInge Heinzl, Marisabel Caballero, Ajay Bhoyar, EW Nutrition
Eliminating necrotic enteritis from your operations starts from a good understanding of what it is, how to prevent it, and how to mitigate its effects on your poultry production.
Necrotic enteritis is a poultry disease caused by an overgrowth of Clostridium perfringens type A, and to a lesser extent type C, in the small intestine. The toxins produced by C. perfringens also damage the intestinal wall. In general, it occurs in broiler chickens of 2-6 weeks of age. In subclinical forms, it is characterized by impaired digestion. Clinical forms lead to severe problems and increased flock mortality in a very short time.
Necrotic enteritis is the cause of USD 6 billion annual losses in global poultry production and this controllable disease is on the rise. One reason is the voluntary or legally required reduction of antibiotics in animal production. This trend is driven by the increasing occurrence of antimicrobial resistance, as well as by consumer demand. Another reason is the reduction of ionophores which, besides their activity against coccidia, also show efficacy against clostridia. When anticoccidial live vaccines are used, the application of these ionophores is not possible and clostridia / necrotic enteritis increase (Williams, 2005).
While this is a widespread problem in all poultry, for broilers in particular, necrotic enteritis and coccidiosis are the most significant health problem.
Clinical and subclinical forms of NE
The clinical form
(c) Rob Moore
…is characterized by acute, dark diarrhea resulting in wet litter and suddenly increasing flock mortality of up to 1% per day after the first clinical signs appear (Ducatelle and Van Immerseel, 2010), sometimes summing up to mortality rates of 50% (Van der Sluis, 2013). The birds have ruffled feathers, lethargy, and inappetence.
Necropsy typically shows ballooned small intestines with a roughened mucosal surface, lesions, and brownish (diphtheritic) pseudo-membranes. There is a lot of watery brown, blood-tinged fluid and a foul odor during post-mortem examination. The liver is dark, swollen, and firm, and the gall bladder is distended (Hofacre et al., 2018).
In the case of peracute necrotic enteritis, birds may die without showing any preliminary signs.
The subclinical form
When birds suffer from the subclinical form, chronic damage to the intestinal mucosa and an increased quantity of mucus in the small intestine lead to impaired digestion and absorption of nutrients resulting in poor growth performance.
The deteriorated feed conversion and the resulting decreased performance become noticeable around day 35 of age. As feed contributes approximately 65-75% of the input cost to produce a broiler chicken, poor feed conversion increases production costs and significantly influences profitability. Often, due to a lack of clear symptoms, this subclinical disease remains untreated and permanently impacts the efficiency of production.
Pathogens
Responsible for necrotic enteritis are Gram-positive, anaerobic bacteria, specific strains of Clostridium perfringens type A and, to a lesser extent, type C (Keyburn et al., 2008).
Clostridia primarily occur in the soil where organic substances are degraded, in sewage, and the gastrointestinal tract of animals and humans. These bacteria produce spores, which are extremely resistant to environmental impact (heat, irradiation, exsiccation) as well as some disinfectants, and can survive for several years. Under suitable conditions, C. perfringens spores can even proliferate in feed or litter.
Clostridium perfringens is a natural inhabitant of the intestine of chickens. In healthy birds, it occurs in a mixture of diverse strains at 102-104 CFU/g of digesta (McDevitt et al., 2006). The disease starts when C. perfringens proliferates in the small intestine, usually due to a combination of factors such as high amount protein, low immunity, and an imbalance in the gut flora. Then, the number rises to 107-109 CFU/g of digesta (Dahiya et al., 2005).
NetB, a key virulence factor for NE
To establish in the host, Clostridium Spp. and other pathogens depend on virulence factors (see infobox). These virulence factors include, for example, “tools” for attachment, evasion or suppression of the host’s immune system, “tools” for getting nutrients, and “tools” for entry into intestinal cells. Over the years, the α-toxin produced by C. perfringens was assumed to be involved in the development of the disease and a key virulence factor. In 2008, Keyburn and coworkers found another key virulence factor by using a C. perfringens mutant unable to produce α-toxin, yet still causing necrotic enteritis.
Thus, another toxin was identified occurring only in chickens suffering from necrotic enteritis: C. perfringens necrotic enteritis B-like toxin (NetB). NetB is a pore-forming toxin. Pore-forming toxins are exotoxins usually produced by pathogenic bacteria, but may also be produced by other microorganisms. These toxins destroy the integrity of gut wall cell membranes. The leaking cell contents serve as nutrients for the bacteria. If immune cells are destroyed, an immune reaction might be partially impacted (Los et al., 2013).
Additionally, pathogenic strains of C. perfringens produce bacteriocins – the most important being Perfrin (Timbermont et al., 2014) – to inhibit the proliferation of harmless Clostridium Spp. strains and to replace the normal intestinal flora of chickens (Riaz et al., 2017).
Examples of virulence factors
1. Adhesins
Enable the pathogen to adhere or attach within the target host site, e.g. via fimbria. Pili enable the exchange of RNA or DNA between pathogens.
2. Invasion factors
Facilitate the penetration and the distribution of the pathogens in the host tissue (invasion and spreading enzymes). For example: hyaluronidase attacking the hyaluronic acid of the connective tissue or flagella enabling the pathogens to actively move.
3. Toxins
Damage the function of the host cells or destroy them (e.g. endotoxins – lipopolysaccharides, exotoxins)
4. Strategies of evasion
Enable the pathogen to bypass the strategies of defense of the immune system (e.g. antiphagocytosis factors provide protection against an attack by phagocytes; specific antibodies are inactivated by enzymes).
A chicken with optimal gut health may be less susceptible to NE. Additional predisposing factors are necessary to allocate nutrients and make the gut environment suitable for the proliferation of these pathogens, enabling them to cause disease (Van Immerseel et al., 2008; Williams, 2005).
Predisposing factors
Feed: composition and particle size
The role of feed in the development of necrotic enteritis should not be underestimated. This is where substances creating an intestinal environment favorable for C. perfringens come into play.
Mycotoxin contamination
Mycotoxins harm gut integrity and create ideal conditions for the proliferation of Clostridium perfringens.
Mycotoxins do not have a direct effect on C. perfringens proliferation, toxin production, or NetB transcription. However, mycotoxins disrupt gut health integrity, creating a favorable environment for the pathogen. For example:
DON provides good conditions for proliferation of C. perfringens by disrupting the intestinal barrier and damaging the epithelium. The possibly resulting permeability of the epithelium and a decreased absorption of dietary proteins can lead to a higher amount of proteins in the small intestine. These proteins may serve as nutrients for the pathogen (Antonissen et al., 2014).
DON and other mycotoxins decrease the number of lactic acid producing bacteria indicating a shift in the microbial balance (Antonissen et al., 2016.).
Eimeria ssp.
An intact intestinal epithelium is the best defense against potential pathogens such as C. perfringens. Here, Coccidiosis comes into play. Moore (2016) showed that by damaging the gut epithelium, Eimeria species give C. perfringens access to the intestinal basal domains of the mucosal epithelium. Then, the first phase of the pathological process takes place and from there, C. perfringens invades the lamina propria. Damage to the epithelium follows (Olkowski et al., 2008). The plasma proteins leaking to the gut and the mucus produced are rich nutrient sources (Van Immerseel et al., 2004; Collier et al., 2008). A further impact of Coccidiosis is shifting the microbial balance in the gut by decreasing the number of e.g., Candidatus savagella which activates the innate immune defense.
Eimeria induce leakage of plasma proteins by killing epithelial cells
They enhance mucus production in the intestine
1+2 lead to an increase in available nutrients and create an environment favorable for the proliferation of C. perfringens.
Not only Eimeria Spp., also other pathogens (e.g. Salmonella Spp., Ascarid larvae, viruses) and agents, such as mycotoxins damaging the intestinal mucosa can pave the way for a C. perfringens infection.
Predisposing factors like wet litter, the moisture of which is essential for the sporulation of Eimeria Spp. oocysts, must also be considered as promoting factors for necrotic enteritis (Williams, 2005).
Immunosuppressive factors
Besides the already explained influencers feed, mycotoxins and coccidia, also other predisposing factors must be mentioned. In general, any factor which induces stress in the animals disrupts the balance of the intestinal flora. The resulting suppression of the immune system contributes to the risk of necrotic enteritis (Tsiouris, 2016). These factors include:
Bacteria: Shivaramaiah and coworkers (2011) investigated a neonatal Salmonella typhimurium infection as a predisposing factor for NE. The early infection causes significant damage to the gut (Porter et al., 1998) Additionally, Hassan et al. (1994) showed that the challenge with Salmonella typhimurium negatively impacted the development of lymphocytes which might also promote a colonization of Clostridium perfringens.
Viruses: Infectious Bursal Disease is known to increase the severity of infections with salmonella, staphylococci, but also clostridia. Another clostridia-promoting viral disease is Marek’s Disease.
Stress: The intestinal tract is particularly sensitive to any type of stress. This stress can be caused by e.g. too high temperatures, high stocking densities, an abrupt change of feed.
Treatment
In acute cases, the farmer should consult a veterinarian and treat his birds.
It must be mentioned that, as the treatment takes place via feed or water, only birds which still consume water or feed may be treated.
Antibiotics
Antibiotics targeting Gram-positive bacteria are commonly used for the treatment of acute NE. The antibiotic choice shall be addressed by a veterinarian, taking into account mode of action and the presence of resistance genes in the farm/flock.
The prophylactic use of antibiotics is not recommened and many countries have already banned it in order to reduce antimicrobial resistance (AMR).
Antimicrobial Resistance (AMR)
Some bacteria are less sensitive to certain antibiotics due to genetic mutations. They are able to:
stimulate the production of enzymes, which break down or modify the antibiotics and inactivate them (1).
eliminate entrances for antibiotics or promote the development of pumps, which discharge the antibiotic before taking effect (2).
change or eliminate molecules to which the antibiotic would bind (targets for the antibiotics).
This means that, when the corresponding antibiotics are used, bacteria resistant against these antibiotics survive. Due to the fact that their competitors have been eliminated they are able to reproduce better. Additionally, this resistance may be transferred by means of “resistance genes”
to daughter cells
via their intake from dead bacteria (3)
through horizontal gene transfer (4)
through viruses (5)
Every application of antibiotics promotes the development of resistance (Robert Koch Institute, 2019). A short-term use, better biosecurity, or an application at low dosage give the bacteria a better chance to adapt.
Bacteriophages
Experimental use of phage treatments have shown to be effective in reducing disease progression and symptoms of necrotic enteritis (Miller et al., 2010). By oral application of a bacteriophage cocktail, Miller and coworkers could reduce mortality by 92% in C. perfringens challenged broilers compared to the untreated control.
Mode of action: the endolysins, highly evolved enzymes produced by bacteriophages, are able to digest the bacterial cell wall for phage progeny release (Fischetti, 2010). However, phages are still not approved by the EFSA.
Prevention
Preventing a disease is always better – and more cost-effective – that its treatment.
How, then, should it be done?
Preventing the conditions that favor the proliferation of Clostridium perfringens and strengthening the host’s immune response lowers the probability of disease.
Besides eliminating the predisposing factors, the main targets are:
There is evidence that most Clostridium strains isolated from birds suffering from necrotic enteritis could induce the disease experimentally, while strains isolated from healthy birds cannot. This confirms that only specific strains are problematic (Ducatelle and Van Immerseel, 2010).
It is therefore of the highest importance to avoid introducing these pathogenic strains to the farm.
Separate clothing, boots, and hand washing/disinfecting facilities in each poultry house
More than 14 days of down time between flocks
Specific measures against coccidiosis
Vaccination
According to parasitologists, 7 to 9 Eimeria species are found in chickens, and they do not cross-protect against each other. An effective vaccination must contain sporulated oocysts of the most critical pathogenic Eimeria species (E. acervulina, E. maxima, E. tenella, E. necatrix, and E. brunetti). The more species contained in the vaccine, the better. However, if not applied the correct way, vaccines can be ineffective or cause reactions in the birds that might lead to NE (Mitchell, 2017).
Anticoccidials
Alternate use of chemicals (synthetic compounds) and ionophores (polyether antibiotics) with different modes of action is important to avoid development of resistance.
Ionophores have a specific mode of action and kill oocysts before they are able to infect birds. Being very small, ionophore molecules can be taken up and diffused into the outer membrane of the sporozoite. There, it decreases the concentration gradient leading to an accumulation of water within the sporozoite causing its bursting.
Diet
Minimizing non-starch polysaccharides (NSPs) in cereals
To prevent a “feeding” of Clostridium perfringens, high content of water-soluble but indigestible NSPs such as wheat, wheat by-products, and barley should be avoided or at least minimized. Additionally, xylanases should be included in the feed formulation to reduce the deleterious effects of NSPs and improve feed energy utilization. Instead of these cereals, maize could be included in the diet. It is considered a perfect ingredient in broiler diets due to its high energy content and high nutrient availability.
Formulating low protein diets/diets with highly digestible amino acids
Feeding low-protein diets supplemented with crystalline amino acids might be beneficial to reduce the risk of necrotic enteritis (Dahiya et al., 2007). To improve protein digestibility and therefore reduce the proliferation of C. perfringens, proteases may be added to the feed.
Avoiding/Minimizing poor quality fats / animal fats in the diet
These fats tend to increase the count of Clostridium perfringens; thus, they should be replaced by higher quality and/or vegetable fats, respectively.
Feed form
In terms of feed form, Engberg et al. (2002) found that birds fed pellets showed a reduced number of Clostridium perfringens in the caeca and the rectum than mash-fed birds. Branton and co-workers (1987) reported a lower mortality by feeding roller-milled (coarsely ground) than hammer-milled feed.
Additives
Additives can be used either to prevent the proliferation of Clostridium perfringens or to change the environmental conditions in a way that proliferation of C. perfringens is prevented.
Probiotics
These live microbial supplements can be used to help to establish, maintain or re-establish the intestinal microflora.
Mode of action:
compete with pathogenic bacteria for substrates and attachment sites
produce antimicrobial substances inhibiting the growth of pathogenic bacteria (Gillor et al., 2008)
bind and neutralize enterotoxins (Mathipa and Thantsha, 2017)
promote immune function of the host (Yang et al., 2012)
Prebiotics
These feed ingredients serve as substrates to promote beneficial bacteria in the intestine.
Mode of action:
D-mannose or fructose, starches non-digestible by birds, selectively stimulate the growth and the activity of the “good” gut flora
Fructooligosaccharides decrease C.perfringens and E. coli in the gut and increase the diversity of Lactobacillus Spp. (Kim et al., 2011)
Galactooligosaccharides, in combination with a B. lactis based probiotic, have been reported to selectively promote the proliferation of Bifidobacterium ssp. (Jung et al., 2008).
Organic acids
Organic acids are often used in animal diets to improve intestinal health.
Mode of action:
decreased pH promotes beneficial bacteria
caprylic acid suppresses C. perfringens, but also Salmonella Spp. by inhibiting their utilization of glucose (Skrivanova et al., 2006)
lauric, citric, oleic and linoleic acid as well as medium-chain fatty acids (C8-C14) impede the growth of C. perfringens
Phytomolecules
Phytomolecules, also known as secondary plant compounds, have been used against pathogens for centuries. In general, two subgroups of these substances are known as effective against Clostridium perfringens:
Tannins
Many studies have shown the efficacy of tannins against different pathogens such as helminths, Eimeria, viruses, and bacteria
Extracts from the chestnut and quebracho trees are effective not only against C. perfringens, but also its toxins (Elizando et al., 2010)
Activity of tannins against Eimeria (Cejas et al., 2011) and Salmonella Sp., two predisposing factors for NE.
Essential Oils
Their hydrophobic characteristic enables them to interact with the lipids of the membrane of C. perfringens.
They can incorporate into the bacterial membrane and disrupt its integrity.
This increases the permeability of the cell membrane for ions and other small molecules such as ATP, leading to the decrease of the electrochemical gradient above the cell membrane and the loss of the cell’s energy equivalents.
Besides their direct effect on Clostridium Spp., a lot of phytomolecules improve gut health and help to prevent a proliferation of Clostridium ssp. and therefore necrotic enteritis.
Mycotoxin/bacterial toxin binders
These binders have two modes of action:
Binding mycotoxins, damage of the intestinal epithelium can be reduced or even prevented, so that the preconditions for Clostridium proliferation are not generated.
Binding toxins produced by Clostridium perfringens can reduce the occurrence or severity of lesions:
Alpha-toxin (phospholipase C) hydrolyses membrane phospholipids and damages erythrocytes, leucocytes, myocytes, and endothelial cells and causes their lysis (Songer, 1996). This leads to necrosis and tissue damage.
Binding NetB toxin, the key virulence factor, could reduce the severity of necrotic enteritis.
Conclusion
The ever-growing trend of reduced antibiotic and ionophore use is contributing to an increased incidence of necrotic enteritis in poultry production.
The subclinical form of necrotic enteritis generally goes unnoticed, resulting in poor feed efficiency and is a major cause of financial losses to poultry producers.
Maintaining optimum gut health is key to preventing the occurrence of necrotic enteritis. In the era of antibiotic-free poultry production, alternatives acting against this pathogenic bacterium and also against its predisposing factors must be considered to control this devastating disease.
References
Annett, C.B., J. R. Viste, M. Chirino-Trejo, H. L. Classen, D. M. Middleton, and E. Simko. “Necrotic enteritis: effect of barley, wheat and corn diets on proliferation of Clostridium perfringens type A.” Avian Pathology 31 (2002): 599– 602. https://doi.org/10.1080/0307945021000024544
Antonissen G, F. Van Immerseel, F. Pasmans, R. Ducatelle, F. Haesebrouck, L. Timbermont, M. Verlinden, G.P.J. Janssens, V. Eeckhaut, M. Eeckhout, S. De Saeger, S. Hessenberger, A. Martel, and S. Croubels. “The mycotoxin deoxynivalenol predisposes for the development of Clostridium perfringens-Induced necrotic enteritis in broiler chickens. PLoS ONE 9 no. 9 (2014): e108775. https://doi.org/10.1371/journal.pone.0108775
Antonissen, G., V. Eeckhaut, K. Van Driessche, L. Onrust , F. Haesebrouck, R. Ducatelle, R.J. Moore, and F. Van Immerseel. “Microbial Shifts Associated With Necrotic enteritis.” Avian Pathol. 45 no. 3 (2016): 308-312. https://doi.org/10.1080/03079457.2016.1152625
Branton, S.L., F.N. Reece, and W.M. Hagler. “Influence of a wheat diet on mortality of broiler chickens associated with necrotic enteritis.” Poultry Sci. 66 (1987): 1326-1330. https://doi.org/10.3382/ps.0661326
Cejas, E., S. Pinto, F. Prosdócimo, M. Batalle, H. Barrios, G. Tellez, and M. De Franceschi. “Evaluation of quebracho red wood (Schinopsis lorentzii) polyphenols vegetable extract for the reduction of coccidiosis in broiler chicks.” International Journal of Poultry Science 10 no. 5 (2011): 344–349. https://doi.org/10.3923/ijps.2011.344.349
Collier, C.T., C.L. Hofacre, A.M. Payne, D.B. Anderson, P. Kaiser, R.I. Mackie, and H.R. Gaskins. “Coccidia-induced mucogenesis promotes the onset of necrotic enteritis by supporting Clostridium perfringens growth.” Veterinary Immunology and Immunopathology 122 (2008):104–115.
Dahiya, J.P., D. Hoehler, A.G. Van Kessel, and M.D. Drew. “Effect of different dietary methionine sources on intestinal microbial populations in broiler chickens.” Poultry Science 86 (2007):2358–2366
Dahiya, J.P., D. Hoehler, D.C. Wilkie, A.G. van Kessel, and M.D. Drew. “Dietary glycine concentration affects intestinal Clostridium perfringens and Lactobacilli populations in broiler chickens.” Poultry Science 84 no.12 (2005):1875-85. https://doi.org/10.1093/ps/84.12.1875
Diaz Carrasco, J.M., L.M. Redondo, E.A. Redondo, J.E. Dominguez, A.P. Chacana, and M.E. Fernandez Miyakawa. “Use of plant extracts as an effective manner to control Clostridium perfringens induced necrotic enteritis in poultry.” BioMed Research International (2016): Article ID 3278359. https://dx.doi.org/10.1155/2016/3278359
Ducatelle, R. and F. van Immerseel. “Necrotic enteritis: emerging problem in broilers.” WATTAgNet.com – Poultry Health and Disease (April 9, 2010).
Elizondo, A.M., E.C. Mercado, B.C. Rabinovitz, and M.E. Fernandez-Miyakawa. “Effect of tannins on the in vitro growth of Clostridium perfringens.” Veterinary Microbiology 145 no. 3-4 (2010): 308–314. https://doi.org/10.1016/j.vetmic.2010.04.003
Engberg, R.M., M.S. Hedemann, and B.B. Jensen. “The influence of grinding and pelleting of feed on the microbial composition and activity in the digestive tract of broiler chickens.” · British Poultry Science 43 no. 4 (2002):569-579. https://doi.org/10.1080/0007166022000004480
Fischetti, V.A. “Bacteriophage endolysins: A novel anti-infective to control Gram-positive pathogens.” J Med Microbiol. 300 no. 6 (2010): 357–362. https://doi.org/10.1016/j.ijmm.2010.04.002
Gillor, O., A. Etzion and M.A. Riley. “The dual role of bacteriocins as anti- and probiotics.” Appl Microbiol Biotechnol. 81 no. 4 (2008): 591–606. https://doi.org/10.1007/s00253-008-1726-5
Hassan, J. O., and R. Curtiss III. “Virulent Salmonella typhimurium induced lymphocyte depletion and immunosuppression in chickens.” Infect. Immun. 62 (1994):2027–2036 https://doi.org/10.1128/IAI.62.5.2027-2036.1994
Hofacre, C.L., J.A. Smith, and G.F. Mathis. “Invited Review. An optimist’s view on limiting necrotic enteritis and maintaining broiler gut health and performance in today’s marketing, food safety, and regulatory climate.” Poultry Science 97 (2018):1929–1933. https://dx.doi.org/10.3382/ps/pey082
Jung, S.J., R. Houde, B. Baurhoo, X. Zhao, and B. H. Lee. “Effects of galacto-oligosaccharides and a bifidobacteria lactis-based probiotic strain on the growth performance and fecal microflora of broiler chickens.” Poultry Science 87 (2008):1694–1699. https://doi.org/10.3382/ps.2007-00489
Kaldhusdal and Skjerve. “Association between cereal contents in the diet and incidence of necrotic enteritis in broiler chickens in Norway.” Preventive Veterinary Medicine 28 (1996):1-16. https://doi.org/10.1016/0167-5877(96)01021-5
Keyburn, A. L., S. A. Sheedy, M. E. Ford, M. M. Williamson, M. M. Awad, J. I. Rood, and R. J. Moore. “Alpha-toxin of Clostridium perfringens is not an essential virulence factor in necrotic enteritis in chickens.” Infect. Immun. 74 (2006): 6496–6500. https://doi.org/10.1128/IAI.00806-06
Keyburn, A.L., J.D. Boyce, P. Vaz, T.L. Bannam, M.E. Ford, D. Parker, A. Di Rubbo, J.I. Rood, and R.J. Moore. “NetB, a new toxin that is associated with avian necrotic enteritis caused by Clostridium perfringens.” PLoS Pathog 4 no. 2, e26 (2008): 0001-0011. https://doi.org/10.1371/journal.ppat.0040026
Kim, G.-B., Y. M. Seo , C. H. Kim , and I. K. Paik. “Effect of dietary prebiotic supplementation on the performance, intestinal microflora, and immune response of broilers.” Poultry Science 90 (2011):75–82. https://doi.org/10.3382/ps.2010-00732
Knap, I., B. Lund, A. B. Kehlet, C. Hofacre, and G. Mathis. “Bacillus licheniformis prevents necrotic enteritis in broiler chickens.” Avian Diseases 54 no. 2 (2010):931-935. https://doi.org/10.1637/9106-101509-ResNote.1
Knarreborg, A., M.A. Simon, R.M. Engberg, B.B. Jensen, and G.W. Tannock. “Effects of Dietary Fat Source and Subtherapeutic Levels of Antibioticon the Bacterial Community in the Ileum of Broiler Chickensat Various Ages.” Applied and Environmental Microbiology 68 no. 12 (2002): 5918-5924. https://doi.org/0.1128/AEM.68.12.5918–5924.2002
Kocher, A. and M. Choct. “Improving broiler chicken performance. The efficacy of organic acids, prebiotics and enzymes in controlling necrotic enteritis.” Australian Government-Rural Industries Research and Development Corporation. Publ. no. 08/149 (2008).
Kondo, F. “In vitro lecithinase activity and sensitivity to 22 antimicrobial agents of Clostridium perfringens isolated from necrotic enteritis of broiler chickens.” Research in veterinary Science 45 (1988): 337-340. https://doi.org/10.1016/S0034-5288(18)30961-5
Kubena, L.F., J.A. Byrd, C.R. Young, and D.E. Corrier. “Effects of tannic acid on cecal volatile fatty acids and susceptibility to Salmonella typhimurium colonization in broiler chicks.” Poultry Science 80, no. 9 (2001): 1293–1298. https://doi.org/10.1093/ps/80.9.1293
Los, F.C.O., T.M. Randis, R.V. Aroian, and A.J. Ratner. “Role of pore-forming toxins in bacterial infectious diseases.” Microbiology and Molecular Biology Reviews 77 (2013): 173-207 https://doi.org/10.1128/MMBR.00052-12
M’Sadeq S.A., Shubiao Wu, Robert A. Swick, Mingan Choct. “Towards the control of necrotic enteritis in broiler chickens with in-feed antibiotics phasing-out worldwide.” Animal Nutrition 1 (2015): 1-11. https://dx.doi.org/10.1016/j.aninu.2015.02.004
Mathipa, M.G. and M.S. Thantsha. “Probiotic engineering: towards development of robust probiotic strains with enhanced functional properties and for targeted control of enteric pathogens.” Gut Pathog. 9 no. 28 (2017). https://doi.org/10.1186/s13099-017-0178-9
McDevitt, R.M., J.D. Brooker, T. Acamovic, and N.H.C. Sparks. “Necrotic enteritis, a continuing challenge for the poultry industry.” World’s Poultry Science Journal 62; World’s Poultry Science Association (June 2006). https://doi.org/10.1079/WPS200593
Miller, R.W., J. Skinner, A. Sulakvelidze, G.F. Mathis, and C.L. Hofacre. “Bacteriophage therapy for control of necrotic enteritis of broiler chickens experimentally infected with Clostridium perfringens.” Avian Diseases 54 no. 1 (2010): 33-40. https://doi.org/10.1637/8953-060509-Reg.1
Mitsch, P., K. Zitterl-Eglseer, B. Köhler, C. Gabler, R. Losa, and I. Zimpernik. “The Effect of Two Different Blends of Essential Oil Components on the Proliferation of Clostridium perfringens in the Intestines of Broiler Chickens.” Poultry Science 83 (2004):669–675. https://doi.org/10.1093/ps/83.4.669
Olkowski, A.A., C. Wojnarowicz, M. Chirino-Trejo, B. Laarveld, and G. Sawicki. “Sub-clinical necrotic enteritis in broiler chickens: Novel etiological consideration based on ultra-structural and molecular changes in the intestinal tissue.” Veterinary Science 85 (2008): 543–553. https://doi.org/10.1016/j.rvsc.2008.02.007
Pan, D. and Z. Yu. “Intestinal microbiome of poultry and its interaction with host and diet.” Gut Microbes 5 no. 1 (2014): 108–119. https://dx.doi.org/10.4161/gmic.26945
Rougière, N. and B. Carré. “Comparison of gastrointestinal transit times between chickens from D + and D- genetic lines selected for divergent digestion efficiency.” Animal 4 no. 11 (2010): 1861-1872. https://doi.org/10.1017/S1751731110001266
Santos, F.B.O., B.W. Sheldon, A.A. Santos Jr., and P.R. Ferket. ”Influence of housing system, grain type, and particle size on Salmonella colonization and shedding of broilers fed triticale or corn-soybean meal diets.” Poultry Science 87 (2008): 405-420. https://dx.doi.org/10.3382/ps.2006-00417
Schiavone, A. , K. Guo, S. Tassone, L .Gasco, E. Hernandez, R. Denti, and I. Zoccarato. “Effects of a Natural Extract of Chestnut Wood on Digestibility, Performance Traits, and Nitrogen Balance of Broiler Chicks.” Poult Sci. 87 no. 3 (2008): 521-527. https://doi.org/10.3382/ps.2007-00113
Shivaramaiah, S., R. E. Wolfenden, J. R. Barta, M. J. Morgan, A. D. Wolfenden, B. M. Hargis, and G. Téllez. „The role of an early Salmonella typhimurium infection as a predisposing factor for necrotic enteritis in a laboratory challenge model.” Avian Diseases 55 (2011): 319-323. https://doi.org/10.1637/9604-112910-ResNote.1
Singh, Y., V. Ravindran, T.J. Wester, A.L. Molan, and G. Ravindran. “ Influence of feeding coarse corn on performance, nutrient utilization, digestive tract measurements, carcass characteristics, and cecal microflora counts of broilers.” Poultry Science 93 (2014): 607–616. https://dx.doi.org/10.3382/ps.2013-03542
Skrivanova, E., M. Marounek, V. Benda, and P. Brezina. “Susceptibility of Escherichia coli, Salmonella sp. and Clostridium perfringens to organic acids and monolaurin.” Veterinarni Medicina 51 no. 3 (2006): 81–88. https://doi.org/10.17221/5524-VETMED
Stanley D., Wu S.-B., Rodgers N., Swick R.A., and Moore R.J. “Differential Responses of Cecal Microbiota to Fishmeal, Eimeria and Clostridium perfringens in a Necrotic Enteritis Challenge Model in Chickens.” PLoS ONE 9 no. 8 (2014): e104739. https://doi.org/10.1371/journal.pone.0104739
Tan, L., D. Rong, Y. Yang, and B. Zhang. “Effect of Oxidized Soybean Oils on Oxidative Status and Intestinal Barrier Function in Broiler Chickens.” Brazilian Journal of Poultry Science 18 no. 2 (2018): 333-342. http://dx.doi.org/10.1590/1806-9061-2017-0610
Tan, L., D. Rong, Y. Yang, and B. Zhang. “The Effect of Oxidized Fish Oils on Growth Performance, Oxidative Status, and Intestinal Barrier Function in Broiler Chickens.” J. Appl. Poult. Res. 28 (2019): 31-41. http://dx.doi.org/10.3382/japr/pfy013
Timbermont L., A. Lanckriet, J. Dewulf, N. Nollet, K. Schwarzer, F. Haesebrouck, R. Ducatelle, and F. Van Immerseel. “Control of Clostridium perfringens-induced necrotic enteritis in broilers by target-released butyric acid, fatty acids and essential oils.” Avian Pathol. 39 no. 2 (2010): 117-21. https://doi.org/10.1080/03079451003610586
Tsiouris, V. “Poultry management: a useful tool for the control of necrotic enteritis in poultry.” Avian Pathol. 45 no. 3 (2016):323-325. https://doi.org/10.1080/03079457.2016.1154502
Van der Most, P.J., B. de Jong, H.K. Parmentier and S. Verhulst. “Trade-off between growth and immune function: a meta-analysis of selection experiments.” Functional Ecology 25 (2011): 74-80. https://doi.org/0.1111/j.1365-2435.2010.01800.x
Van der Sluis, W. “Clostridial enteritis is an often underestimated problem.” Worlds Poult. Sci. J. 16 (2000):42–43.
Van Immerseel, F., J. De Buck, F. Pasmans, G. Huyghebaert, F. Haesebrouck, and R. Ducatelle. “Clostridium perfringens in poultry: an emerging threat of animal and public health.” Avian Pathology 33 (2004): 537-549. https://doi.org/10.1080/03079450400013162
Van Immerseel, F., J.I. Rood, R.J. Moore, and R.W. Titball. “Rethinking our understanding of the pathogenesis of necrotic enteritis in chickens.” Trends in Microbiology 17 no. 1 (2008):32-36. https://doi.org/10.1016/j.tim.2008.09.005
Wade, B., A.L. Keyburn, T. Seemann, J.I. Rood, and R.J. Moore. “Binding of Clostridium perfringens to collagen correlates with the ability to cause necrotic enteritis in chickens.” Veterinary Microbiology 180 no. 3–4 (2015): 299-303. https://doi.org/10.1016/j.vetmic.2015.09.019
Williams, R.B. “Intercurrent coccidiosis and necrotic enteritis of chickens: rational, integrated disease management by maintenance of gut integrity.” Avian Pathology 34 no. 3 (2005):159-180. https://doi.org/10.1080/03079450500112195
Yang , C.M., G.T. Cao, P.R. Ferket, T.T. Liu, L. Zhou, L. Zhang, Y.P. Xiao, and A. G. Chen. “ Effects of probiotic, Clostridium butyricum, on growth performance, immune function, and cecal microflora in broiler chickens.” Poultry Science 91 (2012): 2121–2129. https://dx.doi.org/10.3382/ps.2011-02131
Are endotoxins behind your low livestock productivity?
Impaired health status of the animals in stressful situations or an aggravation of the disease after antibiotic treatment? The culprit might be endotoxins.
What are endotoxins?
Origin
Endotoxins, together with exotoxins, are bacterial toxins. In contrast to exotoxins, which are actively secreted by living bacteria, endotoxins (name “endotoxin” greek; endo = inside; toxin = poison) are components of the outer cell membrane of gram-negative bacteria such as Escherichia coli, Salmonella, Shigella, and cyanobacteria (blue-green algae). They are only released in case of
bacterial death due to effective host defense mechanism or activities of certain antibiotics
Biochemically, endotoxins are lipopolysaccharides (LPS). They are composed of a relatively uniform lipid fraction (Lipid A) and a species-specific polysaccharides chain. Their toxicity is mainly due to the lipid A; the polysaccharide part modifies their activity. Unlike the bacteria, their endotoxins are very heat stable and resist sterilization. The names endotoxin and lipopolysaccharides are used synonymously with “endotoxin” emphasizing on the occurrence and biological activity and “lipopolysaccharide” on the chemical structure (Hurley, 1995).
General structure of Gram-negative lipopolysaccharides (according to Erridge et al., 2002)
Impact
Endotoxins belong to the so-called pyrogen-agents (they provoke fever), activating several immunocompetent cells’ signaling pathways. Early contact with endotoxins leads to activation and maturation of the acquired immune system. Braun-Fahrländer and co-workers (2002) found that children exposed to endotoxins had fewer problems with hay fever, atopic asthma, and atopic sensitization. This might be an explanation that in human populations, after the elevation of the hygiene standards, an increase of allergies could be observed.
Different animal species show different sensibilities to endotoxin infusions, e.g. (healthy) dogs, rats, mice, hens tolerate concentrations ≥1mg / kg body weight, whereas (healthy) ruminants, pigs, horses react very sensible already at concentrations <5μg / kg body weight (Olson et al., 1995 cited in Wilken, 2003).
Reasons for increased exposure of the organism to endotoxins
Endotoxins usually occur in the gut, as the microflora also contains gram-negative bacteria. The precondition for endotoxins to be harmful is their presence in the bloodstream. In the bloodstream, low levels of endotoxins can still be handled by the immune defense, higher levels can get critical. An increase of endotoxins in the organism results from higher input and/or lower clearance or detoxification rate.
Higher input of endotoxins into the organism
The “normal” small amounts of endotoxins arising in the gut due to regular bacterial activity and translocated to the organism have no negative impact as long as the liver performs its clearance function. Also, the endotoxins stored in the adipose tissue are not problematic. However, some factors can lead to a release of the endotoxins or translocation of endotoxins into the organism:
1. Stress
Stress situations such as parturition, surgeries, injuries can lead to ischemia in the intestinal tract and translocation of endotoxins into the organism (Krüger, 1997). Other stress situations in animal production, such as high temperatures and high stocking densities, contribute to higher endotoxin levels in the bloodstream. Stress leads to a higher metabolic demand for water, sodium, and energy-rich substances. For a higher availability of these substances, the intestinal barrier’s permeability is increased, possibly leading to a higher translocation of bacteria and their toxins into the bloodstream.
Examples:
Higher levels of endotoxins in pigs in an experimental study suffering from stress due to loading and transport, elevated temperatures (Seidler (1998) cited in Wilken (2003)).
Marathon runners (Brock-Utne et al., 1988) and racing horses (Baker et al., 1988) also showed higher endotoxin concentrations in the blood proportional to the running stress; thus, trained horses showed lower concentrations than untrained.
2. Lipolysis for energy mobilization
If endotoxins, due to continuous stress, consistently get into the bloodstream, they can be stored in the adipose tissue. The SR-B1 (Scavenger receptor B1, a membrane receptor belonging to the group of pattern recognition receptors) binds to lipids and the lipopolysaccharides, probably promoting the incorporation of LPS in chylomicrons. Transferred from the chylomicrons to other lipoproteins, the LPS finally arrives in the adipose tissue (Hersoug et al., 2016). The mobilization of energy by lipolysis e.g., during the beginning of lactation, for example, leads to a re-input of endotoxins into the bloodstream.
3. Damage of the gut barrier
In normal conditions, due to bacterial activity, endotoxins are present in the gut. Damage of the gut barrier allows translocation of these endotoxins (and bacteria) into the bloodstream.
4. Destruction of Gram-negative bacteria
Another “source” for endotoxins is the destruction of the bacteria. This can be done on the one hand by the organism’s immune system or by treatment with bactericidal substances targeting gram- bacteria (Kastner, 2002). To prevent an increased release of endotoxins, in the case of Gram-negative bacteria, a treatment with bacteriostatic substances only inhibiting the growth and not destroying the bacteria, or with bactericidal in combination with LPS-binding agents, would be a better alternative (Brandenburg, 2014).
5. Proliferation of gram-negative bacteria
As gram-negative bacteria also release small amounts of endotoxins when they grow, everything promoting their proliferation also leads to an increase of endotoxins:
Imbalanced feeding
High yielder cows e.g., are fed diets rich in starch, fat, and protein. Increased feeding of fat leads to a higher concentration of endotoxins in the organism, as the same “transporter” (scavenger receptor class B type 1, SR-BI) can be used (Hersoug et al., 2016) for the absorption of fat as well as for the absorption of endotoxins.
In a study with humans as representors of the monogastric species, Deopurkar and co-workers gave three different drinks (glucose – 100% carbohydrate, orange juice – 92% carbohydrate, and cream – 100% fat) to healthy participants. Only the cream drink increased the level of lipopolysaccharides in the plasma.
Infectious diseases
Infectious diseases like mastitis, metritis, and other infections caused by gram-bacteria such as E. coli, Salmonella, etc. can be regarded as sources of endotoxin release.
Decreased detoxification or degradation
Main responsible organ: the liver
Task: detoxification and degradation of translocated endotoxin. The liver produces substances such as lipopolysaccharide binding proteins (LBP) which are necessary for binding and neutralizing lipopolysaccharide structures.
During the post-partum period, the organism is in a catabolic phase, and lipolysis is remarkably increased for energy generation due to milk production. Increased lipolysis leads, as mentioned before, to a release of endotoxins out of the adipose tissue but also fatty degeneration of the liver. A fatty degenerated liver cannot bring the same performance in endotoxin clearance than a normal liver (Andersen, 2003; Andersen et al., 1996; Harte et al., 2010; Wilken, 2003). In a study conducted by Andersen and co-workers (1996), they couldn’t achieve complete clearance of endotoxins in cows with fatty livers. The occurrence of hepatic lipidoses increases after parturition (Reid and Roberts, 1993; Wilken, 2003).
Also, other diseases of the liver influence endotoxin clearance in the liver. Hanslin and co-workers (2019) found an impaired endotoxin elimination in pigs with pre-existing systemic inflammatory response syndrome.
Relation between lipid metabolism and endotoxin metabolism (according to Fürll, 2000, cited in Wilken, 2003)
Issues caused by endotoxins
Endotoxins, on the one hand, can positively stimulate the immune system when occurring in small amounts (Sampath, 2018). According to McAleer and Vella (2008), lipopolysaccharides are used as natural adjuvants to strengthen immune reaction in case of vaccination by influencing CD4 T cell responses. On the other hand, they are involved in the development of severe issues like MMA-Complex (Pig Progress) or a septic shock (Sampath, 2018).
MMA Complex in sows
MMA in sows is a multi-factorial disease appearing shortly after farrowing (12 hours to three days), which is caused by different factors (pathogens such as E. coli, Klebsiella spps., Staph. spps. and Mycoplasma spps., but also stress, diet). MMA is also known as puerperal syndrome, puerperal septicemia, milk fever, or toxemia. The last name suggests that one of the factors intervening in the disease is bacterial endotoxins. During the perinatal phase, massive catabolism of fat takes place to support lactation. The sows often suffer from obstipation leading to higher permeability of the intestinal wall, with bacteria, respectively endotoxins being transferred into the bloodstream. Another “source” of endotoxins can be the udder, as the prevalence of gram-negative bacteria in the mammary glands is remarkable (Morkoc et al., 1983).
The endotoxins can lead to an endocrine dysfunction: ↑ Cortisol, ↓ PGF2α, ↓Prolactin, ↓ Oxytocin. MMA stands for:
– Mastitis, a bacterial infection of the udder.
Mastitis can be provoked from two sides: on the one hand, endotoxemia leads to an elevation of cytokines (IL1, 6, TNFα). Lower Ca- and K-levels cause teat sphincter to be less functional, facilitating the entry of environmental pathogens into the udder and resulting in mastitis. On the other hand, due to farrowing stress, Cortisol concentrations get higher. The resulting immunosuppression enables E. coli to proliferate in the udder.
– Metritis, an infection of the uterus with vulvar discharges:
It leads to reduced contractions and, therefore, to prolonged and/or complicated farrowing or dead piglets. Metritis can be promoted by stress causing a decrease in oxytocin and prostaglandin F2α secretion. Morkoc and co-workers (1983) didn’t find a relation between metritis and endotoxins.
– Agalactia, a reduction or total loss of milk production:
In many cases, agalactia is not detected until the nursing litter shows signs of hunger and/or weight loss. At worst, the mortality rate in piglets increases. Probably, milk deficiency is caused by lower levels of the hormones involved in lactation. Prolactin levels e.g., may be dramatically reduced by small volumes of endotoxin (Smith and Wagner, 1984). The levels of oxytocin are often half those in normal sows (Pig Progress, 2020).
Endotoxin shock
A septic shock can be the response to the release of a high amount of endotoxins.
In the case of an infection with gram-negative bacteria, the animals are treated with (often bactericidal) antibiotics. Also, the immune system is eliminating the bacteria. Due to bacterial death, endotoxins are massively released. When not bound, they activate the immune system including macrophages, monocytes, and endothelial cells. Consequently, high amounts of cellular mediators like TNFα, Interleukin 1 (IL-1), IL-6, and leukotrienes are released. High levels of pro-inflammatory cytokines activate the complement and coagulation cascade. In some animals, then the production of prostaglandins and leukotrienes is stimulated, implicating high fever, decreased blood pressure, generation of thrombi in the blood, collapse, damaging several organs, and lethal (endotoxic) shock.
Endotoxic shock only occurs to a few susceptible animals, although the whole herd may have been immune-stimulated. A more severe problem is the decrease in the normally strong piglets’ performance, deviating resources from production to the immune system because of the endotoxemia.
Amplified diarrhea
Lipopolysaccharides lead to an augmented release of prostaglandins, which influence gastrointestinal functions. Together with leukotrienes and pro-inflammatory mediators within the mucosa, they reduce intestinal absorption (Munck et al., 1988; Chiossone et al., 1990) but also initiate a pro-secretory state in the intestine. Liang and co-workers (2005) observed a dose-dependent accumulation of abundant fluid in the small intestine resulting in increased diarrheagenic activity and a decreased gastrointestinal motility in rats.
Conclusion
Acting against Gram- bacteria can result in an even more severe issue – endotoxemia. Endotoxins, besides having a direct negative impact on the organism, also contribute to some diseases. Supporting gut health by different approaches, including the binding of toxins, helps to keep animals healthy.
By Inge Heinzl, EW Nutrition
References
Andersen, P.H. “Bovine endotoxicosis – some aspects of relevance to production diseases. A review.” Acta vet. scand. Suppl. 98 (2003): 141-155. DOI: 10.1186/1751-0147-44-S1-P57
Andersen, P.H., N. Jarløv, M. Hesselholt, and L. Bæk. “Studies on in vivo Endotoxin Plasma Disappearance Times in Cattle.” Zentralblatt für Veterinärmedizin. Reihe A 43 no. 2(1996): 93-101. DOI: 10.1111/j.1439-0442.1996.tb00432.x
Baker, B., S.L. Gaffin, M. Wells, B.C. Wessels and J.G. Brock-Utne. “Endotoxaemia in racehorses following exertion.” Journal of the South African Veterinary Association June (1988): 63-66. https://journals.co.za/docserver/fulltext/savet/59/2/1341.pdf?expires=1598542211&id=id&accname=guest&checksum=E50C766D318776E09CA41DA912F14CAD
Beutler, B. and T. Rietschel. “Innate immune sensing and its roots: The story of endotoxin.” Nature Reviews / Immunology 3(2003): 169-176. DOI: 10.1038/nri1004
Braun-Fahrländer, C., J. Riedler, U. Herz, W. Eder, M. Waser, L. Grize, S. Maisch, D. Carr, F. Gerlach, A. Bufe, R.P. Lauener, R. Schierl, H. Renz, D. Nowak and E. von Mutius. „Environmental exposure to endotoxin and its relation to asthma in school-age children. ”The New England Journal of Medicine 347 (2002): 869-877. DOI: 10.1056/NEJMoa020057.
Chiossone, D. C., P.L. Simon, P.L. Smith. “Interleukin-1: effects on rabbit ileal mucosal ion transport in vitro.” European Journal of Pharmacology 180 no. 2-3 (1990): 217–228. DOI: 10.1016/0014-2999(90)90305-P.
Deopurkar R., H. Ghanim, J. Friedman, et al. “Differential effects of cream, glucose, and orange juice on inflammation, endotoxin, and the expression of Toll-like receptor-4 and suppressor of cytokine signaling-3.” Diabetes care 33 no. 5 (2010):991–997.
Erridge, C., E. Bennett-Guerrero, and I.R. Poxton. “Structure and function of lipopolysaccharides.” Microbes and Infection 4 no. 8 (2002): 837-851. DOI: 10.1016/s1286-4579(02)01604-0
Fritsche, D. “Endotoxinpromovierte bakterielle Translokationen und Besiedelung von Uterus und Euter beim Hochleistungsrind im peripartalen Zeitraum.“ Dissertation. Leipzig, Univ., Veterinärmed. Fak. (1998)
Hanslin, K., J. Sjölin, P. Skorup, F. Wilske, R. Frithiof, A. Larsson, M. Castegren, E. Tano, and M. Lipcsey. “The impact of the systemic inflammatory response on hepatic bacterial elimination in experimental abdominal sepsis.” Intensive Care Medicine Experimental 7 (2019): art. 52. https://doi.org/10.1186/s40635-019-0266-x
Harte, A.L., N.F. da Silva, S.J. Creely, K.C. McGee, T. Billyard, E.M. Youssef-Elabd, G. Tripathi, E. Ashour, M.S. Abdalla, H.M. Sharada, A.I. Amin, A.D. Burt, S. Kumar, C.P. Day and P.G. McTernan. “Research Elevated endotoxin levels in non-alcoholic fatty liver disease.” Journal of Inflammation 7 (2010): 15-24. DOI: 10.1186/1476-9255-7-15
Hersoug, L.-G., P. Møller, and S. Loft. “Gut microbiota-derived lipopolysaccharide uptake and trafficking to adipose tissue: implications for inflammation and obesity.” Obesity Reviews 17 (2016): 297–312. DOI: 10.1111/obr.12370
Hurley, J. C. “Endotoxemia: Methods of detection and clinical correlates.” Clin. Microbiol. Rev. 8 (1995): 268–292. DOI: 10.1128/CMR.8.2.268
Kastner, A. “Untersuchungen zum Fettstoffwechsel und Endotoxin-Metabolismus bei Milchkühen vor dem Auftreten der Dislocatio abomasi.“ Inaug. Diss. Universität Leipzig, Veterinärmed. Fak. (2002). https://d-nb.info/967451647/34
Krüger M. “Escherichia coli: Problemkeim in der Nutztierhaltung.“ Darmflora in Symbiose und Pathogenität. Ökologische, physiologische und therapeutische Aspekte von Escherichia coli. 3. Interdisziplinäres Symposium. Alfred-Nissle-Gesellschaft (Ed.). Ansbach, 28.-29. Nov. (1997): 109-115.
Liang, Y.-C., H.-J. Liu, S.-H. Chen, C.-C. Chen, L.-S. Chou, and L. H. Tsai. “ Effect of lipopolysaccharide on diarrhea and gastrointestinal transit in mice: Roles of nitric oxide and prostaglandin E2.” World J Gastroenterol. 11 no. 3 (2005): 357–361. DOI: 10.3748/wjg.v11.i3.357
McAleer, J.P. and Vella, A.T. “Understanding how lipopolysaccharide impacts CD4 T cell immunity.” Crit. Rev. Immunol. 28 no. 4 (2008): 281-299. DOI:10.1615/CRITREVIMMUNOL.V28.I4.20
Morkok, A., L. Backstrom, L. Lund, A.R.Smith. “Bacterial endotoxin in blood of dysgalactic sows in relation to microbial status of uterus, milk, and intestine.” JAVMA 183 (1983): 786-789. PMID: 6629987
Munck, L.K., A. Mertz-Nielsen, H. Westh, K. Buxhave, E. Beubler, J. Rask-Madsen. “Prostaglandin E2 is a mediator of 5-hydroxytryptamine induced water and electrolyte secretion in the human jejunum.” Gut 29 no. 10 (1988): 1337-1341
Sampath, V.P. “Bacterial endotoxin-lipopolysaccharide; structure, function and its role in immunity in vertebrates and invertebrates.” Agriculture and Natural Resources 52 no. 2 (2018): 115-120. https://doi.org/10.1016/j.anres.2018.08.002
Seidler, T. “Freies Endotoxin in der Blutzirkulation von Schlachtschweinen: eine Ursache für bakterielle Translokationen?“ Diss. Universität Leipzig, Veterinärmed. Fak. (1998).
Smith, B.B. and W.C. Wagner. “Suppression of prolactin in pigs by Escherichia coli endotoxin.“ Science 224 no. 4649 (1984): 605-607
Wilken, H. “Endotoxin-Status und antioxidative Kapazität sowie ausgewählte Stoffwechselparameter bei gesunden Milch- und Mutterkühen.“ Inaugural Diss. Universität Leipzig (2003).
8 ways COVID-19 might impact your business
by Ilinca Anghelescu, EW Nutrition
By now there is no doubt the economic impact of COVID-19 will be massive. The question is, how exactly will the pandemic impact your business – and what can you do to mitigate or prevent what’s coming?
Since January, the international community has been aware of the seriousness and ease of contagion of COVID-19. Despite that, internet searches for “coronavirus” only exploded over the past couple of weeks. Worldwide, as a population, we were more interested in Harry Styles, home loan rates and Gal Gadot than in the impending crisis.
In other words, we individually, as well as markets, were slow to understand and accept the long-term implications of the pandemic.
Google searches for “Coronavirus” since December and which countries did it most
Now that the seriousness of the pandemic has hit home, there are known losses in some industries: airlines worldwide are expected to lose $29BN, with some smaller players being forced out of business by reduced demand. Other low-margin industries, like restaurants or the travel industry, are likewise already sustaining major losses.
Figure 1 – Visualization of COVID-19 impact on markets / Restaurant reservations
And while we are seeing the world slowly understanding and adapting to a new reality, you, like everyone else, have to prepare for the impact of COVID-19 on your business. Quick note: though it may appear so at first, not all the ways the pandemic affects businesses are negative!
Labor shortage
As workers are affected by the pandemic, many will either choose to stay home or will be forced to. In some countries, self-isolation measures are elective. In other regions that are more severely affected, the government may require workers in non-essential industries to not break isolation measures. This may affect your company at all levels, from processing feed or feeding animals to delivering goods across quarantined regions.
What you can do: – Identify proximity workers you can rely on
– Preemptively create crisis scenarios for a reduced workforce
– Create a waiting list of short-term labor resources
Supply shortages
Lockdowns in China, Italy or Spain already provide examples of what happens when regions go into full isolation. Consider how massive shortages in the antibiotic supply from China or shipment delays across the world, for instance, are already affecting the animal production industry.
What you can do: – Overstock now
– Contact alternative suppliers to create an improved supply chain
– Check expiration dates for your existing supply and consume early dates first
– Choose alternatives for products with an uncertain supply chain
Demand shortages
Depending on the industry and the market, you may be faced with reduced demand. Simply consider the fact that reduced demand for restaurants will lower, in turn, demand for supplies for the restaurant: less meat, butter, milk consumed in restaurants is less meat ordered.
What you can do: – Prepare for basic production only
– Prepare to stock raw materials long-term if possible
– Discuss with suppliers to cut or minimize deliveries
Government policies
The Food and Drug Administration, the United States’ highest authority in food and medicine safety, announced it would suspend inspections of foreign food manufacturers. The impact of this decision could be felt in the quality of foreign feed or raw materials quality. Other governments are already – or might soon be – limiting imports, restricting non-essential activities, offering financial packages for at-risk businesses,
What you can do: – Review government policy updates on a daily basis and tailor production and operations to ensure compliance
– Give early feedback to government measures
– Apply early for relief measures, even if not severely impacted yet
Lower biosecurity standards
Even now, biosecurity is implemented more in theory than in practice. Routinely there are small infringements – and we can expect their numbers to grow massively in times of crisis. People are less likely to go through the motions if personnel is reduced, supervision is less strict, and the financial pressure of the pandemic is high. This will trigger severe risks for the animal and feed production industries, as well as for product packaging. Workers who hide symptoms to be able to sustain wages; workers attempting to speed up work because of reduced personnel; reduced or looser inspections and monitoring – all these could perpetrate risks to your operations and to the population at large.
What you can do: – Tighten biosecurity measures and controls
– Supplement lower government monitoring with additional on-location measures
– If you operate with a reduced workforce, periodically check how downscaling affects biosecurity implementation
Immediate economic downturn
At this point, almost all industries have a global component: your raw materials may be imported; the ingredients in the antibiotics or vaccines you use may come from anywhere around the world; your packaging may be produced in China; your software solutions may come from the Indian subcontinent; your quality controls may be managed by a consultancy from a distant European country – and so on. However much we may try to avoid it, there may be immediate repercussions on your business. Either because your goods may be inaccessible for part of the world, because of lower demand on the consumer side, or because of diminished production capacities, you may feel the impact of the pandemic sooner rather than later.
What you can do: – Cut costs for non-essentials
– If you are in feed production, consider stocking on toxin binders, search for alternative suppliers, and assess your supply levels
– If you are in livestock production, employ solutions for animal health and welfare to lower disease risk
– Apply for government bailout early
– Assess your export strategy and prepare to zoom in on domestic
– Assess long-term payroll capacities during diminished business demand
Changing consumption trends
It turns out that, after all, the impact could be positive for some industries. The meat industry seems to be doing relatively well, despite the challenges. While in China, severely affected by ASF on the animal side and now by COVID-19 on the human side, meat production was dramatically affected, in other regions demand for – and supply of – animal protein is stable. Consider the new opportunities for frozen or prepackaged food products: as less fresh meat is consumed in restaurants or bought because of infrequent store visits, consumption of these meat products and by-products is not expected to go down – in fact, it may well increase.
The market might, however, first have to be taught to embrace these prepackaged or frozen products.
What you can do: – Prepare for less fresh meat demand by upping prepackaged meat production
– Teach your end-users about the benefits of frozen products, from meat to egg whites, for instance
Negative impact for others, positive impact for you
While the negative effects are real, there are ways you can balance the COVID-19 impact by taking advantage of some of the positives. Consider that, to give just one example, the energy market is likely going to take a hit. This, in turn, may lead to lower fuel costs for farmers.
Reduced travel means more savings for your company, and while working from home (WFH) may lead in some cases to somewhat reduced productivity, taking an early stand and instructing your team on how to structure WFH days will help preserve productivity while cutting down on energy, fuel and other travel costs, cleaning, in-office equipment depreciation, and other such expenses.
What you can do: – Check your balance sheets regularly
– Transfer savings from quick benefits into investments into long-term strategy
– Most importantly – never panic!
Ongoing research into treating COVID-19 already shows great promise. While we do not yet know how long these unusual circumstances will last, you can make provisions for the near future and think long-term of how to protect your businesses from this pandemic or any future such challenges.
Egg antibody technology for nursery pig application
Pigs at birth having insufficient immunity are simply not able to cope with the stress situations they face early in life. They of course become susceptible to the many pathogens common in the farrowing house. The resulting negative effects are added medical costs for treating the pigs and often an increased mortality. Strengthening the immune system by applying egg antibodies (IgY) during the first days of piglet’s life is a proven viable option.
Immunity in pigs
Humans and animals are protected against diseases by specific antibodies (AB). Newborns receive the antibodies maternally (passive immunity) and they produce them after contact with pathogens (active immunity). But unlike humans, who receive maternal AB within the womb, sows possess a multi-layered placenta which prevents the transfer of AB during gestation. Therefore, an early intake of AB from colostrum is essential. This intake should begin immediately after birth as absorption decreases with every hour. But, the maternal antibodies are only a “starter immune kit”. The young pigs immediately must begin to develop their own “active immunity”.
Figure 1: Immune status of the young pig (Sieverding, 2000)
Figure 1 shows gaps of low immunity shortly after birth and about six weeks after, as the level of passive immunity begins to drop and the active immunity starts to build up. The strength of the passive immune protection depends on quantity and quality of the colostrum consumed by the nursery pig. The quality is determined by the pathogens the sows have been confronted with during their life. Young gilts and sows with only short adaptation time into the herd often do not have the farm-specific antibodies needed to pass to their nursing pigs.
How can egg antibodies serve as a tool ?
Young pigs are challenged by different pathogens (see figure 2). From studies made by the German internist Felix Klemperer (Klemperer, 1893) we know that hens which come in contact with pathogens (in his studies with tetanus bacillus) produce antibodies against these pathogens. The antibodies are transferred to the egg yolk and are intended for being a starter protection kit for the chicks.
Technology allows us today to produce a highly valuable product based on egg powder. It contains significant amounts of natural egg immunoglobulins (IgY – immunoglobulins from the yolk). These egg antibodies mainly act in the gut. There they recognize and tie up pathogens and in this way render them ineffective.
Figure 2: Commonly occurring pathogens causing diarrhea in pigs as they age
Not all egg powders are equal
Early work done by Kellner et al. (1994) showed the effectiveness of egg powder containing relevant antibodies against diarrhea causing pathogens in nursery pigs. In the trial they evaluated three groups receiving egg powder with relevant antibodies, egg powder from regular eggs or no additive (negative control).
Results:
(Figure 3: Effects of egg powder with relevant antibodies and egg powder from regular eggs in comparison to a negative control):
The group that received egg powder containing relevant antibodies completely recovered from diarrhea on day 4.
In the group fed normal egg powder on day 4 still 9 % suffered from severe diarrhea.
In the control more than 70 % showed either severe or light diarrhea.
The results show that the effectiveness of egg powder depends on its content of antibodies.
Reducing mortality by oral administration of egg antibodies
The effectiveness of egg antibodies in pigs was demonstrated also in other studies (Erhard et al., 1996, Yokoyama et al., 1992, Nguyen et al., 2005, Yokoyama et al., 1997). One trial conducted in Germany showed promising results concerning reduction of mortality in the farrowing unit. For the trial 96 sows and their litters were divided evenly into three groups (32 sows each) and the pigs were treated as follows:
Group
Number of pigs
Treatment
Negative Control
530
no treatment
Group EP – 1+3
494
egg powder-based product Globigen Pig Doser, 4 ml on day 1, 2 ml on day 3
Group EP – 1, 2, 3
527
egg powder-based product Globigen Pig Doser, 4 ml on day 1, 2 ml on day 2 and 3
*EP = Egg powder-based product
Results: Figure 4 shows regardless of the frequency of oral application dosage given to pigs both were very supportive and significantly reduced mortality compared to the control. This resulted in a higher number of weaned pigs than in the control.
Figure 4: Mortality and resulting number of pigs weaned per sow and year
Conclusion
Using egg antibodies in pig nutrition is an effective tool to reduce mortality in young pigs. They can be applied individually by doser (newly weaned pigs) or via powder in the feed. Both practices have proven effectively in commercial operations.
Kellner, J., Erhard, M.H., Renner, M. and Lösch, U.: Therapeutischer Einsatz von spezifischen Eiantikörpern bei Saugferkeldurchfall – ein Feldversuch. Tierärztliche Umschau 49, Nr.1; 31-34 (1994).
Klemperer, F.: Ueber natürliche Immunität und ihre Verwerthung für die Immunisierungstherapie. Arch. f. Exp. Pathol. Pharmakol. 31, 356-382 (1893)
Nguyen, V. S, Bui, H. N. P. and Nguyen, T. P. N.: Hyperimmunized chicken egg protein improves performance of piglets. Asian Pork Magazine, 18-19; April/May (2005).
Yokoyama, H., Peralta, R.C., Diaz, R., Sendo, S., Ikemori, Y. and Kodama, Y.: Passive protective effect of chicken egg yolk immunoglobulins against experimental enterotoxigenic Escherichia coli infection in neonatal piglets. Infection and Immunity, 998-1007; Mar. (1992)



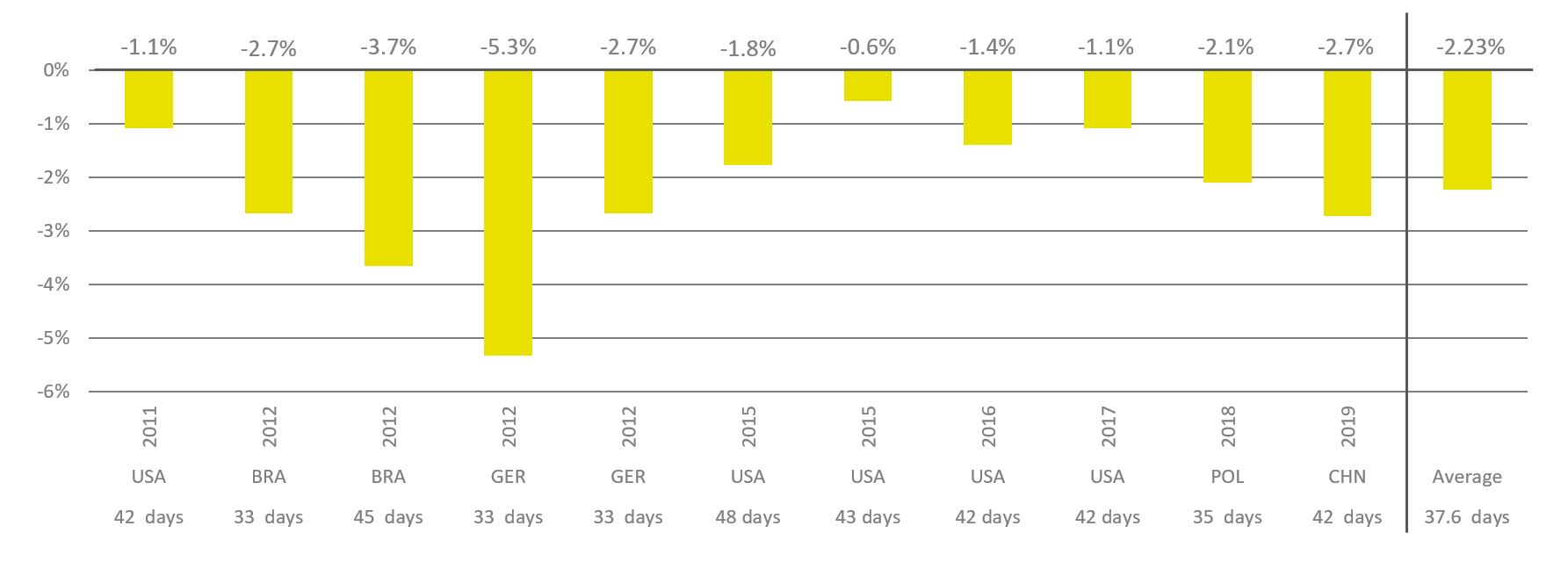
 Feed spoilage through molds, yeasts, and mycotoxins wastes precious resources
Feed spoilage through molds, yeasts, and mycotoxins wastes precious resources  Reducing soybeans in ruminant feeds helps to lower their environmental impact
Reducing soybeans in ruminant feeds helps to lower their environmental impact Feed mill efficiency is key to animal nutrition’s carbon footprint
Feed mill efficiency is key to animal nutrition’s carbon footprint

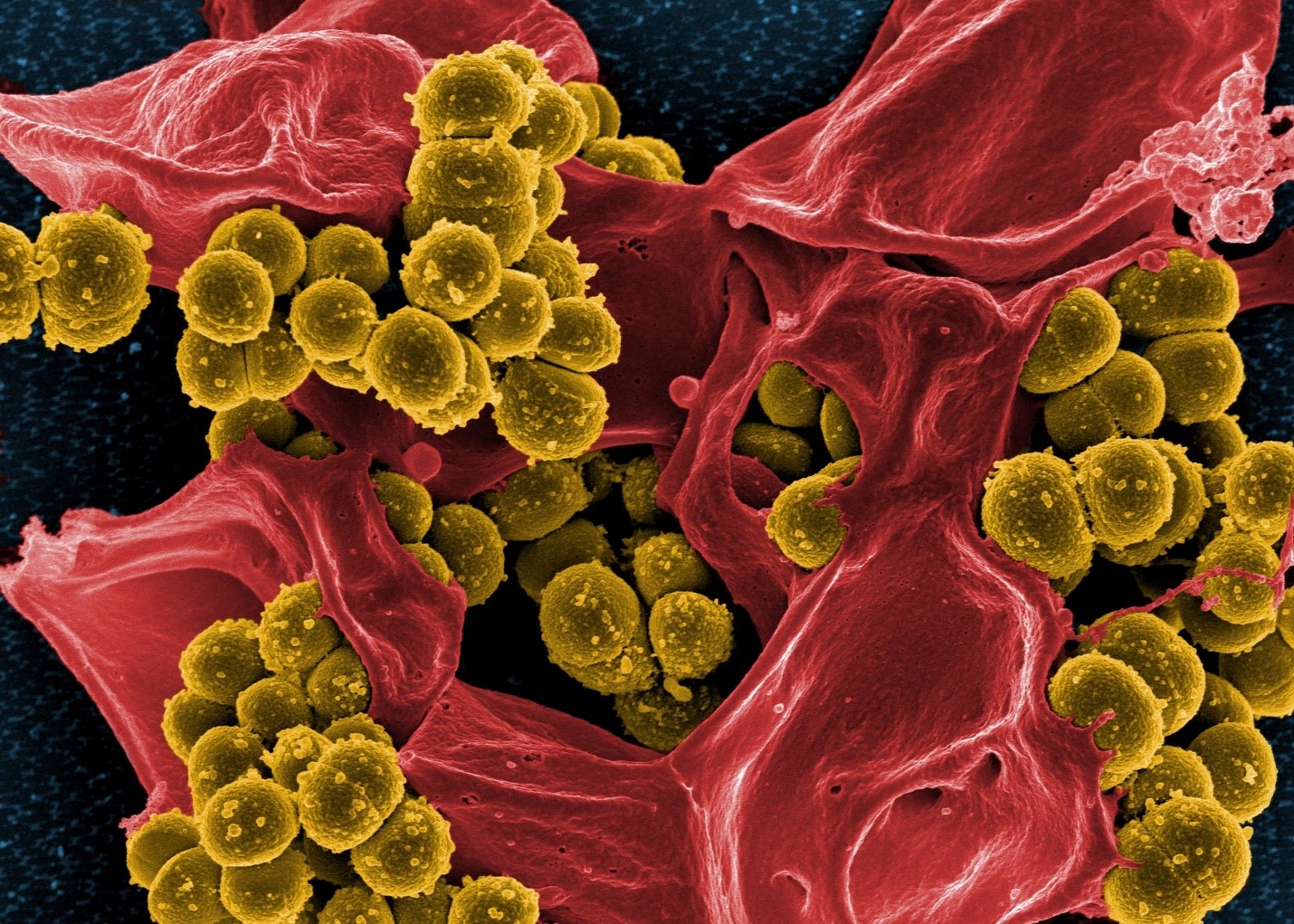



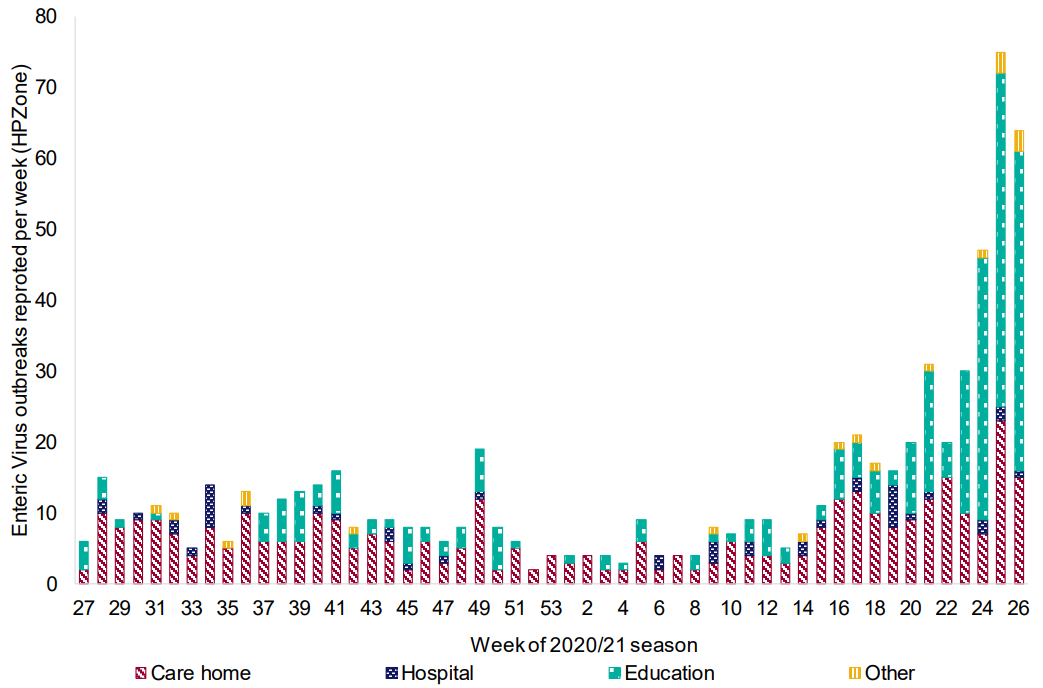
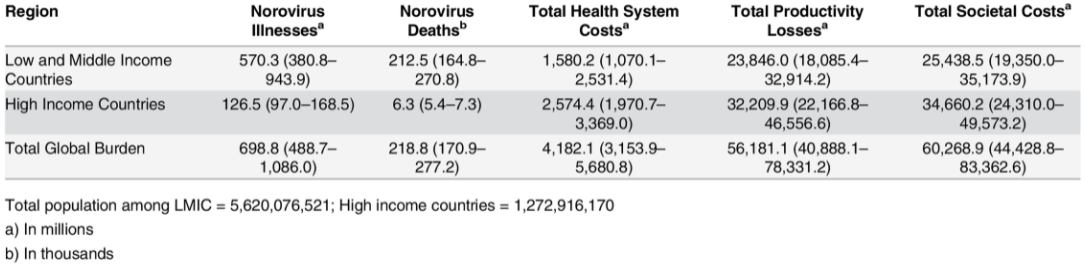



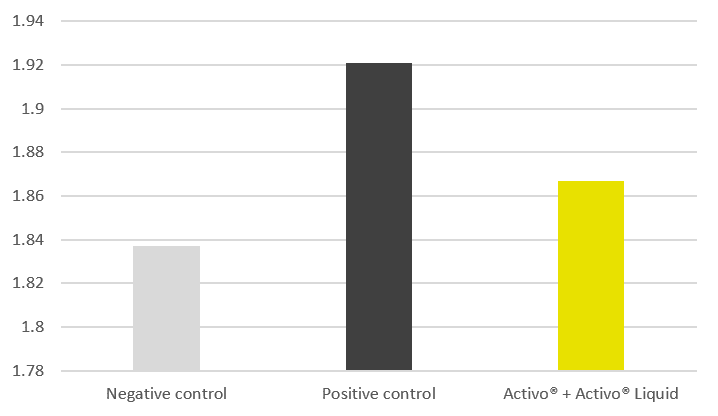
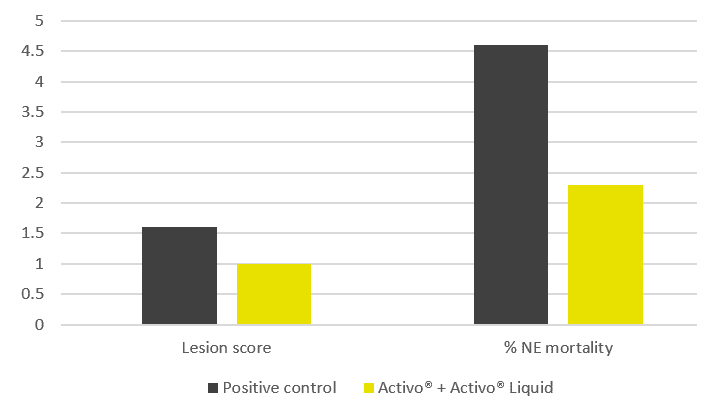



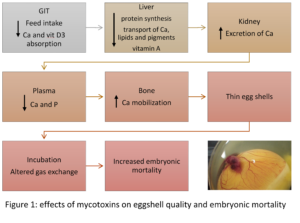


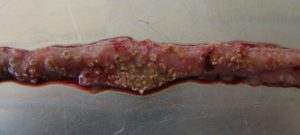




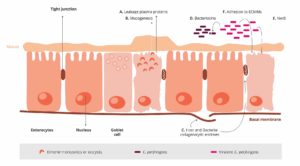
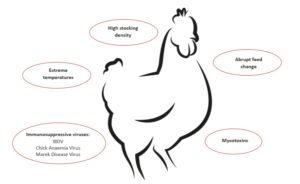
 To prevent a “feeding” of Clostridium perfringens, high content of water-soluble but indigestible NSPs such as wheat, wheat by-products, and barley should be avoided or at least minimized. Additionally, xylanases should be included in the feed formulation to reduce the deleterious effects of NSPs and improve feed energy utilization. Instead of these cereals, maize could be included in the diet. It is considered a perfect ingredient in broiler diets due to its high energy content and high nutrient availability.
To prevent a “feeding” of Clostridium perfringens, high content of water-soluble but indigestible NSPs such as wheat, wheat by-products, and barley should be avoided or at least minimized. Additionally, xylanases should be included in the feed formulation to reduce the deleterious effects of NSPs and improve feed energy utilization. Instead of these cereals, maize could be included in the diet. It is considered a perfect ingredient in broiler diets due to its high energy content and high nutrient availability.