Influence of nutrition and management on eggshell quality
Conference report
Many factors affect eggshell quality, such as nutrition, disease, genetics, environmental conditions, age of birds, stress, egg collection and handling, and packaging and transport. Eggshell quality, however, is primarily related to management and nutrition, not genetics or other factors. It is becoming a bigger issue as the length of the laying period has extended because, as hens get older, shell quality drops.
“The information in the genetics companies’ management guides is for direction and information only, as each egg producer’s production goals and conditions can vary”, says Vitor Arantes, Global Technical Services Manager and Global Nutritionist, Hy-Line International. He advises listening to your birds. For example, “diets should be aligned with the bird’s bodyweight development, rather than the age of birds and following feeding phases according to pre-planned timings for feed changes,” he noted.
Below are some of the nutritional factors impacting eggshell quality that producers should keep top of mind.
Early development and pre-starter diets
“Bodyweight at 6-12 weeks of age is key, but to achieve this goal, bodyweight up to 5 weeks of age is a MUST, stressed,” Dr. Arantes. “This critical period is an investment, so don’t be shy. Poor management in the first 5 weeks will delay production, increase mortality, and prevent the achievement of peak production targets. In turn, it will affect egg quality. Therefore, we must provide proper diets as soon as possible,” he said.
As shown below, chicks hatch with relatively underdeveloped internal organs and systems. During the first 5 weeks of age, the digestive tract and the immune system undergo much of their development. The development of the intestine is crucial for nutrient absorption and will determine a hen’s future production efficiency. Strong intestinal development will also strengthen the immune system and reduce the possibility of future enteric diseases and improve the response to vaccinations.
Multi-phasic body weight development during rearing and the start of lay
Pre-starter diets support the chicks’ transition from being fed by the yolk sac and are relatively high in energy, protein, and the vitamins and minerals required for growth and development. The chicks’ limited digestive capacity post-hatch demands easily digestible raw materials. A crumble containing high-quality, functional ingredients provides a good nutritional start in life. The use of feed additives, such as enzymes to improve digestibility, and synbiotics to aid in the early development of a microbial population and to prevent the intestinal colonization of pathogens, known as competitive exclusion, should be considered.
Teaching hens how to eat – preparing for the pre-peak phase
The objective is to develop sufficient feed intake capacity for the period start of lay, by feeding a developer diet from 10-16 weeks of age. This is a diluted diet with high levels of insoluble fiber to develop feed intake capacity (crop and gizzard).
“You can train pullets to eat by taking advantage of their natural feeding behavior,” commented Dr. Arantes “Because birds consume most of their feed before lights go off, the main feed distribution (60% of the daily ration) should be in the late afternoon, about 2-3 hours before ‘light off’. In the morning, birds will be hungry and finish the feed, including fine particles. Emptying feeders helps to prevent selective eating and will increase the uniformity of the flock. In the middle of the day, there should be no feed in feeders for 60-90 minutes,” he noted.
Don’t neglect the pre-lay phase
Start feeding a pre-lay diet when most pullets show reddening of the combs, which is a sign of sexual maturity. Feed for a maximum of 10–14 days before the point of lay. This is important to increase medullary bone calcium reserves. Large particle calcium should be introduced in this phase. Do not feed pre-lay later than the first egg as it contains insufficient calcium to support egg production.
There can be a negative impact on feed consumption from the sudden increase in dietary calcium levels from 1% to above 4% at the start of lay. Field experience indicates that the use of pre-lay diets helps as a smooth transition between the developer (low calcium and nutrient density) and the peaking diet. Correct feed formulation and matching diet density with consumption will minimize the impact of reduced calcification of bone over the laying cycle and extend the persistency of shell quality. It also helps to avoid the often-reduced appetite/daily feed intake during early production.
The following are suggested for pre-layer feed:
1.25 to 1.40% P
2.5% Ca (50% coarse limestone)
900-1,100g per hen total
Never before 15 weeks of age
Never after 2% hen day (HD) egg production
Understand your limestone
Calcium particle size is important for eggshell quality. Fine calcium carbonate particles pass through the gastrointestinal tract in 2-3 hours, whereas particles above 2mm are retained in the gizzard and will slowly solubilize, delaying the calcium assimilation. Eggshell formation takes 12 to 14 hours and occurs mainly during the night period. Providing a high amount of large calcium particle size before the night, when birds are sleeping, will help laying hens to produce a strong eggshell.
The ratio of coarse to fine calcium particles will increase with bird age as below. Changing the particle size ensures that more calcium will be available at night from the diet instead of from the bone.
Calcium particle size recommendations
Particle size
Starter, Grower, Developer
Pre-Lay
Weeks
17-37
Weeks
38-48
Weeks
49-62
Weeks
63+
Fine (<2mm)
100%
50%
40%
35%
30%
25%
Coarse (2-4mm)
–
50%
60%
55%
70%
75%
The solubility of limestone may differ according to the source. Calcium with high solubility will not be stored for a long time in the gizzard, negating the particle size effect. Dietary calcium levels may need to be adjusted based on the solubility of your limestone. The in vitro solubility of your limestone source can easily be checked on the farm, with a simple technique using hydrochloric acid. The target is to recover 3-6% of the supplemented limestone.
Water
It’s impossible to have good eggshell quality if you don’t have good water intake and good quality water. For example, excessive salt levels in drinking water can cause persistent damage to shell quality.
Conclusion: invest in the rearing phase
Good nutrition and management practices are key to good shell quality. The rearing period is a key developmental time for future success during the laying period – it is an investment phase.
***
EW Nutrition’s Poultry Academy took place in Jakarta and Manila in early September 2023. Vitor Arantes, Global Technical Services Manager and Global Nutritionist, Hy-Line International, was a distinguished guest speaker in this event.
Gustavo Tesolin appointed Regional Director of EW Nutrition LATAM
Gustavo Tesolin
29 January 2024, Visbek – German-based company EW Nutrition, a global provider of functional animal nutrition solutions, has appointed Gustavo Carlos Tesolin as its Regional Director for Latin America.
An agricultural engineer by training, Gustavo Tesolin has forged an international career in the Animal Health business during the last 25 years. With different leadership roles in several major international organizations such as Novartis, Elanco, and Erber Group, Mr. Tesolin brings along important experience in Commercial Operations, P&L drive, and Strategy execution, with special emphasis on team development and geographic expansion.
“I am really excited to join a company as innovation-driven and science-focused as EW Nutrition,” Tesolin said. “The recent launch of VENTAR D, a novel phytogenic specifically designed and developed to improve animal husbandry results, will continue to strengthen our position in the region, together with our winning brands PRETECT D and ACTIVO” and added, “I am eager to take on the challenge and better acquaint the market with an excellent portfolio centered on Gut Health Management, Digestibility, and Feed Quality.”
Jan Vanbrabant, CEO of EW Nutrition, noted that Gustavo Tesolin is “the perfect combination of the right experience and the right attitude. We are happy to have found in him a proven leader with not just excellent market knowledge, but with the same values we share in EW Nutrition: a passion for innovation in the service of our customers, and relentless curiosity and energy to find the right solution.”
This appointment comes on the heels of several top-tier global hires over the last 18 months and reflects the company’s commitment to the Latin American market.
Tesolin will move to Mexico to coordinate EW Nutrition’s expansion in Latin American countries.
About EW Nutrition
EW Nutrition is a German-based international animal nutrition company that offers comprehensive solutions for animal gut health, toxin risk management, growth performance, and more.
Press contact
marketing@ew-nutrition.com
FEFAC: Quick Overview of 2023 EU Compound Feed Production
Total Production 2023: 144.3 million metric tons for farmed animals
Change from 2022: 2% decrease
Factors Influencing Decrease
Political and Market Pressures: Addressing crises and the shift towards sustainable feed.
Climate and Diseases: Effects of droughts, floods, Avian Influenza (AI), and African Swine Fever (ASF) on raw material supply and animal production.
National Policies: Initiatives for greenhouse gas and nitrate emission reduction.
Production Variability: Different trends across EU Member States, with notable decreases in countries like Germany, Ireland, Denmark, and Hungary, and slight increases in Austria, Bulgaria, Italy, and Romania.
Sector-Specific Trends
By Species
Pig Feed: Major decline of nearly 2.5 million tons. Key challenges included:
Loss of export markets, particularly in Asia
Negative media impact in Germany
Significant production drop in Denmark (-13.6%) and Spain (loss of 800,000 metric tons)
Italy’s ongoing struggle with ASF
Poultry Feed: Increase by 0.9 million tons, yet still 700,000 metric tons below 2021 levels. Challenges included declines in Hungary and Czechia due to reduced broiler production.
Cattle Feed: Decrease of 0.8 million tons from 2022.
“Green and animal welfare” policies affecting local production
Summary
The EU’s compound feed production in 2023 faced numerous challenges, leading to an overall decrease. The pig feed sector was most severely hit, while poultry feed showed some recovery. The influence of environmental, economic, and policy factors played a significant role in shaping these trends. Despite the price of feed cereals falling back to the levels seen before Russia’s invasion of Ukraine, these challenges will continue to be felt in 2024.
VISBEK (Germany), 1 September 2023 — EW Nutrition, a leading global provider of functional animal nutrition solutions, welcomes Jan Vanbrabant as its new Chief Executive Officer.
Jan has a PhD degree in microbiology and is an experienced manager in animal health and nutrition, having held leadership roles at DSM, Erber Group, Biomin and Kemin.
“We are very pleased that we have found a strong management lead in Jan, who embodies the philosophy of EW Nutrition”, says Jan Wesjohann, Managing Director of parent company EW Group. “EW Nutrition is an innovation-driven company, with intensive investment in R&D. Together with Jan we are looking to enter the next growth phase of EW Nutrition.”
“I am very excited to be joining the EW Nutrition team,” said Jan Vanbrabant. “EW Nutrition’s long-term focus has created an extremely competitive portfolio. EW Nutrition is uniquely positioned to support its customers in mastering the challenges of the changing animal health and nutrition environment.”
Former CEO Michael Gerrits is heading into retirement after six years leading EW Nutrition. “I want to thank Michael Gerrits for his essential stewardship in bringing the company to the next level,” said Jan Wesjohann.
About EW Nutrition
EW Nutrition is a global animal nutrition company that offers integrators, feed companies, and veterinarians comprehensive solutions for animal gut health and performance, feed quality, digestibility, and more. It is focused on promoting sustainable growth through reduced FCR, natural support against challenges, reduced need for antibiotics, and planet-friendly protein production.
By Lea Poppe, Regional Technical Manager, EW Nutrition
Diarrhea due to infestation with cryptosporidia is one of the most pressing problems in calf rearing. These protozoa, along with rotaviruses, are now considered the most common pathogens in infectious calf diarrhea. Due to their high resistance and thus limited possible control and prevention measures, they have now overtaken other pathogens such as coronaviruses, salmonellae, and E. coli.
Cryptosporidia show complex development
Cryptosporidia are single-celled intestinal parasites. In calves, Cryptosporidium parvum and Cryptosporidium bovis are most commonly found. C. bovis is normally considered nonpathogenic. Accordingly, the disease known as cryptosporidiosis is caused by C. parvum. The rapid tests for determining the diarrheal pathogens, which are increasingly widespread, are usually unsuitable for distinguishing between the individual strains, which can lead to false positive results.
Resistant in the environment, active in the animal
In the environment, cryptosporidia are distributed as oocysts. The oocysts are only about 5 µm in size and have a very resistant shell. They can remain infectious for up to 6 months in high humidity and moderate temperatures. Drought and extreme temperatures (below -18°C and above 65°C) cause the oocysts to die.
After oral ingestion, the oocysts are reactivated by conditions in the gastrointestinal tract (low pH and body temperature): As sporozoites, the parasites attach to the posterior small intestine, causing diarrhea symptomatology. There, they surround themselves with a special protective membrane, and the complex life cycle continues. Only a few days after infection, reproductive forms are detectable in the calf’s intestine, and excretion of infectious oocysts in the feces begins.
Figure 1 (Olias et al., 2018): Life cycle of cryptosporidia: ingested oocysts release four sporozoites that invade host enterocytes (intestinal epithelial cells). There, they develop into trophozoites before asexual and sexual reproduction ensues, and thin- and thick-walled oocysts are formed. Thick-walled oocysts are excreted through the intestine. Thin-walled oocysts may break apart, and the sporozoites may infect other enterocytes, resulting in relapse or prolonged diarrhea. Infestation of the cells leads to their destruction, resulting in villi atrophy or fusion.
Oocysts bring the disease to the animal
Cryptosporidiosis is transmitted either by direct contact of calves with feces from infected animals or indirectly by ingesting contaminated feed, bedding, or water. Each gram of feces excreted by calves showing symptoms may contain up to 100 million oocysts. According to experimental studies, as few as 17 orally ingested oocysts are sufficient to trigger infection. In addition, some multiplication forms can infect other intestinal cells directly within the intestine and thus further advance the disease by autoinfection.
Cryptosporidiosis caused by cryptosporidia often presents with typical diarrhea symptoms and occurs primarily in calves up to 3 weeks of age. Older calves may also be infected with cryptosporidia but usually show no symptoms. Pathogen excretion and, thus, the spread of disease within the herd is nevertheless likely due to the minimal infectious dose.
Damage to the intestinal wall leads to retardation of growth
Attachment of cryptosporidia to the intestinal wall is associated with an inflammatory reaction, regression and fusion of the intestinal villi, and damage to the microvilli. As a result, nutrient absorption in the small intestine is impaired, and more undigested nutrients enter the colon. The microflora starts a fermentation process with lactose and starch, leading to increased lactate levels in the blood and, thus, hyperacidity in the calf. Faintness, unwillingness to drink, recumbency, and growth disorders are the consequences.
Diarrhea often occurs late or not at all and, accordingly, is not considered the main symptom of cryptosporidiosis. When diarrhea occurs, it lasts about 1-2 weeks. The feces are typically watery, greenish-yellow, and are often described as foul-smelling. Due to diarrhea, there is a loss of electrolytes and dehydration.
Studies show: Cryptosporidia are the most prevalent diarrheal pathogens
Several studies in different regions, which examined calf diarrhea and its triggers in more detail, came to a similar conclusion: Cryptosporidia are one of the most common causes of calf diarrhea. In addition, mixed infections often occur.
DL = days of life WL = weeks of life n.m. = not mentioned EP = enteropathogen
Cryptosporidia reduces profit
Infection with cryptosporidia and sometimes subsequent diarrhea entails treatment of the animals and generates costs (veterinarian, medication, electrolyte drinks). In addition, poorer feed conversion, lower growth, and animal losses result in lower production efficiency.
A Scottish study shows 34 kg less gain in the first six months of life compared to healthy calves in beef calves that experienced severe cryptosporidiosis in the first three weeks of life. Similar results are described in lambs, also a susceptible species to cryptosporidia. These studies suggest a long-term negative effect of cryptosporidia on growth performance and production efficiency.
Here’s how you can support your calves against cryptosporidia
High resistance of the pathogens to environmental influences, a very low necessary infection dose combined with an elevated excretion of infectious oocysts, and the possibility of autoinfection make cryptosporidia tough opponents. This is also reflected in their worldwide distribution.
What is the treatment?
Suitable drugs for the treatment of cryptosporidiosis are currently unavailable on the market. The only medicine that can be used in case of cryptosporidiosis infestation may only be administered to calves that have had diarrhea symptoms for 24 hours or less. Accordingly, this agent is usually used only for prevention. Scientific studies on its effectiveness are contradictory; some suggest that it merely delays the onset of the disease. In addition, it is not always easy to use due to the exact dosage that must be followed. Doubling the dose (sometimes happening already due to incorrectly observed intervals between doses) can lead to a toxic overdose.
Accordingly, only the symptoms of the disease – diarrhea with its accompanying symptoms – can be treated. Electrolyte and water losses must be continuously compensated with the help of a high-quality electrolyte drink. The buffer substances contained also reduce the hyperacidity of the blood caused by faulty fermentation in the intestines. For successful treatment, the electrolyte drink should be given in addition to the milk drink. Under no circumstances should the feeding of milk or milk replacer be discontinued because the sick calf urgently needs energy and nutrients. Opinions to the contrary are outdated.
As always: prevention is better than treatment
To make it more difficult for cryptosporidiosis to spread from the outset, it is worth looking at the risk factors. These include direct contact with other calves and general herd size. Furthermore, organic farms seem to have more problems with cryptosporidia. Weather also influences calves born during warmer and, at the same time, wetter weather periods (temperature-humidity index) often get sick.
Due to the limited possibilities for treatment, prevention is of greater importance. For other diarrheal pathogens such as rotavirus, coronavirus, and E. coli, it has become established practice to vaccinate dams to achieve better passive immunization of the calf. However, commercial vaccination against cryptosporidia is not currently available, making dam vaccination as unavailable as calf vaccination.
Accordingly, optimal colostrum management is the first way to protect the calf from cryptosporidia infection. This also confirms the general discussion on the Failure of Passive Transfer: various studies suggest that calves with poor immunoglobulin supply suffer from diarrhea more frequently than calves with good supply, although a concrete link to cryptosporidia itself cannot always be established with certainty.
Furthermore, it is essential to break the chain of infection within farms. In addition to the separate housing of the calves, it is necessary to ensure consistent hygiene. One should take advantage of the pathogen’s weakness as well as its sensitivity to high temperatures and ensure that the water temperature is sufficiently high when cleaning the calf pens and calving area. When disinfecting afterward, it is crucial to consider the spectrum of activity of the agent used, as not all are effective against cryptosporidia.
Egg immunoglobulins support animals against cryptosporidia
Egg immunoglobulins were initially designed to help chicks get started. In this process, hens form antibodies against pathogens they are confronted with. As studies have shown, this also works with cryptosporidia. Cama and Sterling (1991) tested their produced antibodies in the neonatal mouse model and achieved a significant (P≤0.001) reduction in parasites there. Kobayashi et al. (2004) registered decreased binding of sporozoites to the intestinal cell model and their decreased viability in addition to oocyst reduction.
In the IRIG Research Institute (2009, unpublished), feeding egg powder with immunoglobulins against cryptosporidia (10 g/day) to 15 calves reduced oocyst excretion. Before administration, calves excreted an average of 106.42 oocysts/g of feces. After administration of egg powder, only two calves still showed 103.21 oocysts/g feces, and the other 13 of the 15 calves showed no oocyst excretion. All these results are confirmed by positive customer feedback on IgY-based feed supplements.
Egg immunoglobulins and optimal colostrum management as a key solution
Since there are no effective drugs against cryptosporidia, animals must be prophylactically protected against this disease as much as possible. In addition to optimal colostrum management, which means feeding high-quality colostrum (IgG≥50g/L) to the calf as soon as possible after birth, we have products with egg immunoglobulins available to support the calf as a prophylactic against cryptosporidia infestation and thus prevent significant performance losses, especially during rearing.
References
Brainard, J., Hooper, L., McFariane, S., Hammer, C. C., Hunter, P. R., & Tyler, K. (2020). Systemic review of modifiable risk factors shows little evidential support for most current practices in Cryptosporidium management in bovine calves. Parasitology research 119, 3572-3584.
Cama, V. A., and C. R. Sterling. “Hyperimmune Hens as a Novel Source of Anti-Cryptosporidium Antibodies Suitable for Passive Immune Transfer.” University of Arizona. Wiley-Blackwell, January 1, 1991. https://experts.arizona.edu/en/publications/hyperimmune-hens-as-a-novel-source-of-anti-cryptosporidium-antibo.
Kobayashi, C, H Yokoyama, S Nguyen, Y Kodama, T Kimata, and M Izeki. “Effect of Egg Yolk Antibody on Experimental Infection in Mice.” Vaccine 23, no. 2 (2004): 232–35. https://doi.org/10.1016/j.vaccine.2004.05.034.
Lamp, D. O. (25. Januar 2020). Rinder aktuell: Kälberdurchfall durch Kryptosporidien – Hartnäckig und weitverbreitet. BAUERNBLATT, S. 52-53.
Losand, B., Falkenberg, U., Krömker, V., Konow, M., & Flor, J. (2. März 2021). Kälberaufzucht in MV – Alles im grünen Bereich? 30. Milchrindtag Mecklemburg-Vorpommern.
Luginbühl, A., K. Reitt, A. Metzler, M. Kollbrunner, L. Corboz, and P. Deplazes. “Feldstudie Zu Prävalenz Und Diagnostik Von Durchfallerregern Beim Neonaten Kalb Im Einzugsgebiet Einer Schweizerischen Nutztierpraxis.” Schweizer Archiv für Tierheilkunde 147, no. 6 (2005): 245–52. https://doi.org/10.1024/0036-7281.147.6.245.
Olias, P., Dettwiler, I., Hemphill, A., Deplazes, P., Steiner, A., & Meylan, M. (2018). Die Bedeutung der Cryptosporidiose für die Kälbergesundheit in der Schweiz. Schweiz Arch Tierheilkd, Band 160, Heft 6, Juni 2018, 363-374.
Santín, M., Trout, J. M., Xiao, L., Zhou, L., Greiner, E., & Fayer, R. (2004). Prevalence and age-related variation of Cryptosporidium species and genotypes in dairy calves. Veterinary Parasitology 122, 103-117.
Shaw, H. J., Innes, E. A., Marrison, L. J., Katzer, F., & Wells, B. (2020). Long-term production effects of clinical cryptosporidiosis in neonatal calves. International Journal for Parasitology 50, 371-376.
Silverlås, C., H. Bosaeus-Reineck, K. Näslund, and C. Björkman. “Is There a Need for Improved Cryptosporidium Diagnostics in Swedish Calves?” International Journal for Parasitology 43, no. 2 (2013): 155–61. https://doi.org/10.1016/j.ijpara.2012.10.009.
Thomson, Sarah, Carly A. Hamilton, Jayne C. Hope, Frank Katzer, Neil A. Mabbott, Liam J. Morrison, and Elisabeth A. Innes. “Bovine Cryptosporidiosis: Impact, Host-Parasite Interaction, and Control Strategies.” Veterinary Research 48, no. 1 (2017). https://doi.org/10.1186/s13567-017-0447-0.
Uhde, F., Kaufmann, T., Sager, H., Albini, S., Zanoni, R., & Schelling, E. (2008). Prevalence of four enteropathogens in the feces of young diarrhoeic dairy calves in Switzerland. Veterinary Record (163), 362-366.
Piglet Nutrition Scenarios for AGP Removal
Over the past 60 years, antibiotics have played an essential role in the swine industry as a tool that swine producers rely on to control diseases and to reduce mortality. Besides, antibiotics are also known to improve performance, even when used in subtherapeutic doses. The perceived overuse of antibiotics in pig production, especially as growth promoters (AGP), have raised concerns from governments and public opinion, regarding the emergence of multidrug-resistant bacteria, adding a threat not only to animal but also human health. The challenges raised regarding AGPs and the need for their reduction in livestock led to the development of combined strategies such as the “One Health Approach”, where animal health, human health, and the environment are interlaced and must be considered in any animal production system.
In this scenario of intense changes, swine producers must evaluate strategies to adapt their production systems to accomplish the global pressure to reduce antibiotics and still have a profitable operation.
Many of these concerns focus on piglet nutrition, since the use of sub-therapeutic levels of antimicrobials as growth promotors is still a regular practice for preventing post-weaning diarrhea in many countries (Heo et al., 2013; Waititu et al., 2015). Taking that into consideration, this article serves as a practical guide to swine producers through AGP removal and its impacts on piglet performance and nutrition Three crucial points will be addressed:
Why is AGP removal a global trend?
What are the major consequences for piglet nutrition and performance?
What alternatives do we have to guarantee optimum piglet performance in this scenario?
AGP removal: a global issue
Discussions on the future of the swine industry include understanding how and why AGP removal became such important topic worldwide. Historically, European countries have led discussions on eliminating AGP from livestock production. In Sweden, AGPs were banned from their farms as early as 1986. This move culminated into a total ban of AGPs in the European Union in 2006. Other countries followed same steps. In Korea, AGPs were removed from livestock operations in 2011. The USA is also putting efforts into limiting AGPs and the use of antibiotics in pig farms, as published in guidance revised by the Food and Drug Administration (FDA, 2019). In 2016, Brazil and China banned Colistin, and the Brazilian government also announced the removal of Tylosin, Tiamulin, and Lincomycin in 2020. Moreover, countries like India, Vietnam, Bangladesh, Buthan, and Indonesia have announced strategies for AGP restrictions (Cardinal et al., 2019; Davies and Walsh, 2018).
The major argument against AGPs and antibiotics in general is the already mentioned risk of the development of antimicrobial resistance, limiting the available tools to control and prevent diseases in human health. This point is substantiated by the fact that resistant pathogens are not static and exclusive to livestock, but can also spread to human beings (Barbosa and Bünzen, 2021). Moreover, concerns have been raised in regard to the fact that antibiotics in pig production are also used by humans – mainly third-generation antibiotics. The pressure on pig producers increased and it is today multifactorial: from official regulatory departments and stakeholders at different levels, who need to consider public concerns about antimicrobial resistance and its impact on livestock, human health, and the sustainability of farm operations (Stein, 2002).
It is evident that the process of reducing or banning antibiotics and AGPs in pig production is already a global issue and increasing as it takes on new dimensions. As Cardinal et al. (2019) suggest, that process is irreversible. Companies that want to access the global pork market and comply with increasingly stricter regulations on AGPs must re-invent their practices. This, however, is nothing new for the pig industry. For example, pig producers from the US and Brazil have adapted their operations in order to not use ractopamine to meet the requirements from the European and Asian markets. We can be sure, therefore, that the global pig industry will find a way to replace antibiotics.
With that in mind, the next step is to evaluate the consequences of AGP withdrawal from pig diets and how that affects the animals’ overall performance.
Consequences in piglet health and performance
Swine producers know very well that weaning pigs is challenging. Piglets are exposed to many biological stressors during that transitioning period, including introducing the piglets to new feed composition (going from milk to plant-based diets), abrupt separation from the sow, transportation and handling, exposure to new social interactions, and environmental adaptations, to name a few. Such stressors and physiological challenges can negatively impact health, growth performance, and feed intake due to immune systems dysfunctions (Campbell et al. 2013). Antibiotics have been a very powerful tool to mitigate this performance drop. The question then is, how difficult can this process become when AGPs are removed entirely?
Many farmers around the world still depend on AGPs to make the weaning period less stressful for piglets. One main benefit is that antibiotics will reduce the incidence of PWD, with subsequent improved growth performance (Long et al., 2018). The weaning process can create ideal conditions for the overgrowth of pathogens, as the piglets’ immune system is not completely developed and therefore not able to fight back. Those pathogens present in the gastrointestinal tract can lead to post-weaning diarrhea (PWD), among many other clinical diseases (Han et al., 2021). PWD is caused by Escherichia coli and is a global issue in the swine industry, as it compromises feed intake and growth performance throughout the pig’s life, also being a common cause for losses due to young pig death (Zimmerman, 2019).
Cardinal et al. (2021) also highlight that the hypothesis of a reduced intestinal inflammatory response is one explanation for the positive relationship between the use of AGPs and piglet weight gain. Pluske et al. (2018) point out that overstimulation of the immune system can negatively affect pig growth rate and feed use efficiency. The process is physiologically expensive in terms of energy and also can cause excessive prostaglandin E2 (PGE2) production, leading to fever, anorexia, and reduction in pig performance. For instance, Mazutti et al. (2016) showed an increased weight gain of up to 1.74 kg per pig in animals that received colistin or tylosin in sub-therapeutic levels throughout the nursery. Helm et al. (2019) found that pigs medicated with chlortetracycline in sub-therapeutic levels increased average daily gain in 0.110 kg/day. Both attribute the higher weight to the decreased costs of immune activation determined by the action of AGPs on intestinal microflora.
On the other hand, although AGPs are an alternative for controlling bacterial diseases, they have also proved to be potentially deleterious to the beneficial microbiota and have long-lasting effects caused by microbial dysbiosis – abundance of potential pathogens, such as Escherichia and Clostridium; and a reduction of beneficial bacteria, such as Bacteroides, Bifidobacterium, and Lactobacillus (Guevarra et al., 2019; Correa-Fiz, 2019). Furthermore, AGPs reduced microbiota diversity, which was accompanied by general health worsening in the piglets (Correa-Fiz, 2019).
It is also important to highlight that the abrupt stress caused by suckling to weaning transition has consequences in diverse aspects of the function and structure of the intestine, which includes crypt hyperplasia, villous atrophy, intestinal inflammation, and lower activities of epithelial brush border enzyme (Jiang et al., 2019). Also, the movement of bacteria from the gut to the body can occur when the intestinal barrier function is deteriorated, which results in severe diarrhea and growth retardation. Therefore, nutrition and management strategies during that period are critical, and key gut nutrients must be used to support gut function and growth performance.
With all of that, it is more than never necessary to better understand the intestinal composition of young pigs and finding strategies to promote gut health are critical measures for preventing the overgrowth and colonization of opportunistic pathogens, and therefore being able to replace AGPs (Castillo et al., 2007).
Viable alternatives for protecting the piglets
The good news is that the swine industry already has effective alternatives that can replace AGP products and guarantee good animal performance.
Immunoglobulins from egg yolk (IgY) have proven to be a successful alternative to weaned piglet nutrition. Investigations have shown that egg antibodies improve the piglets’ gut microbiota, making it more stable (Han et al., 2021). Moreover, IgY optimizes piglet immunity and performance while reducing occurrences of diarrhea caused by E. coli, rotavirus, and Salmonella sp. (Li et al., 2016).
Phytomolecules (PM) are also potential alternatives for AGP removal, as they are bioactive compounds with antibacterial, antioxidant, and anti-inflammatory characteristics (Damjanović-Vratnica et al., 2011; Lee and Shibamoto, 2001). When used for piglet diet supplementation, phytomolecules optimize intestinal health and improve growth performance (Zhai et al., 2018).
Han et al. (2021) evaluated a combination of IgY (Globigen® Jump Start, EW Nutrition) and phytomolecules (Activo®, EW Nutrition) supplementation in weaned piglets’ diets. Results from that study (Table 1 and 2) showed that this strategy decreases the incidence of PWD and coliforms, increases feed intake, and improves the intestinal morphology of weaned pigs, making that combination a viable AGP replacement.
Table 1. Effect of dietary treatments on the growth performance of weaned pigs challenged with E. coli K88 (SOURCE: Han et al., 2021).
Table 2. Effect of dietary treatments on the post-weaning diarrhea incidence of weaned pigs challenged with E. coli K88 (%) (SOURCE: Han et al., 2021).
A trial conducted at the Institute of Animal Sciences of the Chinese Academy of Agricultural Sciences, China, supplemented weaning pigs challenged by E. coli K88 with a combination of PM (Activo®, EW Nutrition) and IgY (Globigen® Jump Start). The trial reported that this combination (AC/GJS) showed fewer diarrhea occurrences than in animals from the positive group (PC) during the first week after the challenge and similar diarrhea incidence to the AGP group during the 7th and 17th days after challenge (Figure 1).
Figure 1 – Incidence of diarrhea (%). NC: negative group, PC: positive group, AGP: supplementation with AGP, AC/GJS: combination of PM (Activo, EW Nutrition) and IgY (Globigen Jump Start).
The same trial also showed that the combination of these non-antibiotic additives was as efficient as the AGPs in improving pig performance under bacterial enteric challenges, showing positive effects on body weight, average daily gain (Figure 2), and feed conversion rate (Figure 3).
Figure 2 – Body weight (kg) and average daily gain (g). NC: negative group, PC: positive group, AGP: supplementation with AGP, AC/GJS: combination of PM (Activo, EW Nutrition) and IgY (Globigen Jump Start).
Figure 3 – Feed conversion rate. NC: negative group, PC: positive group, AGP: supplementation with AGP, AC/GJS: combination of PM (Activo, EW Nutrition) and IgY (Globigen Jump Start).
The multiple benefits of using IgY in piglet nutrition strategies are also highlighted by Rosa et al. (2015), Figure 4, and Prudius (2021).
Figure 4. Effect of treatments on the performance of newly weaned piglets. Means (±SEM) followed by letters a,b,c in the same group of columns differ (p < 0.05). NC (not challenged with ETEC, and diet with 40 ppm of colistin, 2300 ppm of zinc, and 150 ppm of copper). Treatments challenged with ETEC: GLOBIGEN® (0.2% of GLOBIGEN®); DPP (4% of dry porcine plasma); and PC (basal diet) (SOURCE: Rosa et al., 2015).
Conclusions
AGP removal and overall antibiotic reduction seems to be the only direction that the global swine industry must take for the future. From the front line, swine producers demand cost-effective AGP-free products that don’t compromise growth performance and animal health. Along with this demand, finding the best strategies for piglet nutrition in this scenario is critical in minimizing the adverse effects of weaning stress. With that in mind, alternatives such as egg immunoglobulins and phytomolecules are commercial options that are already showing great results and benefits, helping swine producers to go a step further into the future of swine nutrition.
References
Damjanović-Vratnica, Biljana, Tatjana Đakov, Danijela Šuković and Jovanka Damjanović, “Antimicrobial effect of essential oil isolated from Eucalyptus globulus Labill. from Montenegro,” Czech Journal of Food Sciences 29, no. 3 (2011): 277-284.
Pozzebon da Rosa, Daniele, Maite de Moraes Vieira, Alexandre Mello Kessler, Tiane Martin de Moura, Ana Paula Guedes Frazzon, Concepta Margaret McManus, Fábio Ritter Marx, Raquel Melchior and Andrea Machado Leal Ribeiro, “Efficacy of hyperimmunized hen egg yolks in the control of diarrhea in newly weaned piglets,” Food and Agricultural Immunology 26, no. 5 (2015): 622-634. https://doi.org/10.1080/09540105.2014.998639
Freitas Barbosa, Fellipe, Silvano Bünzen. Produção de suínos em épocas de restrição aos antimicrobianos–uma visão global. In: Suinocultura e Avicultura: do básico a zootecnia de precisão (2021): 14-33. https://dx.doi.org/10.37885/210203382
Correa-Fiz, Florencia, José Maurício Gonçalves dos Santos, Francesc Illas and Virginia Aragon, “Antimicrobial removal on piglets promotes health and higher bacterial diversity in the nasal microbiota,” Scientific reports 9, no. 1 (2019): 1-9. https://doi.org/10.1038/s41598-019-43022-y
Stein, Hans H , “Experience of feeding pigs without antibiotics: a European perspective,” Animal Biotechnology13 no. 1(2002): 85-95. https://doi.org/10.1081/abio-120005772
Helm, Emma T, Shelby Curry, Julian M Trachsel, Martine Schroyen, Nicholas K Gabler, “Evaluating nursery pig responses to in-feed sub-therapeutic antibiotics”, PLoS One 14 no. 4 (2019). https://doi.org/10.1371/journal.pone.0216070.
Hengxiao Zhai, Hong Liu, Shikui Wang, Jinlong Wu and Anna-Maria Kluenter, “Potential of essential oils for poultry and pigs,” Animal Nutrition4, no. 2 (2018): 179-186. https://doi.org/10.1016/j.aninu.2018.01.005
Pluske, J. R., Kim, J. C., Black, J. L. “Manipulating the immune system for pigs to optimise performance,” Animal Production Science 58, no 4, (2018): 666-680. https://doi.org/10.1071/an17598
Zimmerman, Jeffrey, Locke Karriker, Alejandro Ramirez, Kent Schwartz, Gregory Stevenson, Jianqiang Zhang (Eds.), “Diseases of Swine,” 11 (2019), Wiley Blackwell.
Campbell, Joy M, Joe D Crenshaw & Javier Polo, “The biological stress of early weaned piglets”, Journal of animal science and biotechnology 4, no. 1 (2013):1-4. https://doi.org/10.1186/2049-1891-4-19
Jung M. Heo, Opapeju, F. O., Pluske, J. R., Kim, J. C., Hampson, D. J., & Charles M. Nyachoti, “Gastrointestinal health and function in weaned pigs: a review of feeding strategies to control post‐weaning diarrhoea without using in‐feed antimicrobial compounds,” Journal of animal physiology and animal nutrition 97, no. 2 (2013): 207-237. https://doi.org/10.1111/j.1439-0396.2012.01284.x
Junjie Jiang, Daiwen Chen, Bing Yu, Jun He, Jie Yu, Xiangbing Mao, Zhiqing Huang, Yuheng Luo, Junqiu Luo, Ping Zheng, “Improvement of growth performance and parameters of intestinal function in liquid fed early weanling pigs,” Journal of animal science 97, no. 7 (2019): 2725-2738. https://doi.org/10.1093/jas/skz134
Cardinal, Kátia Maria, Ines Andretta, Marcos Kipper da Silva, Thais Bastos Stefanello, Bruna Schroeder and Andréa Machado Leal Ribeiro, “Estimation of productive losses caused by withdrawal of antibiotic growth promoter from pig diets – Meta-analysis,” Scientia Agricola78, no.1 (2021): e20200266. http://doi.org/10.1590/1678-992X-2020-0266
Cardinal, Katia Maria, Marcos Kipper, Ines Andretta and Andréa Machado Leal Ribeiro, “Withdrawal of antibiotic growth promoters from broiler diets: Performance indexes and economic impact,” Poultry science 98, no. 12 (2019): 6659-6667. https://doi.org/10.3382/ps/pez536
Mazutti, Kelly, Leandro Batista Costa, Lígia Valéria Nascimento, Tobias Fernandes Filho, Breno Castello Branco Beirão, Pedro Celso Machado Júnior, Alex Maiorka, “Effect of colistin and tylosin used as feed additives on the performance, diarrhea incidence, and immune response of nursery pigs”, Semina: Ciências Agrárias 37, no. 4 (2016): 1947. https://doi.org/10.5433/1679-0359.2016v37n4p1947
Lee, Kwang-Geun and Takayuki Shibamoto, “Antioxidant activities of volatile components isolated from Eucalyptus species,” Journal of the Science of Food and Agriculture81, no. 15 (2001): 1573-1579. https://doi.org/10.1002/jsfa.980
Long, S. F., Xu, Y. T., Pan, L., Wang, Q. Q., Wang, C. L., Wu, J. Y., … and Piao, X. S. Mixed organic acids as antibiotic substitutes improve performance, serum immunity, intestinal morphology and microbiota for weaned piglets,” Animal Feed Science and Technology 235, (2018): 23-32.
Castillo, Marisol, Susana M Martín-Orúe, Miquel Nofrarías, Edgar G Manzanilla and Josep Gasa, “Changes in caecal microbiota and mucosal morphology of weaned pigs”, Veterinary microbiology 124, no. 3-4 (2007): 239-247. https://doi.org/10.1016/j.vetmic.2007.04.026
Dyar, Oliver J, Jia Yin, Lilu Ding, Karin Wikander, Tianyang Zhang, Chengtao Sun, Yang Wang, Christina Greko, Qiang Sun and Cecilia Stålsby Lundborg, “Antibiotic use in people and pigs: a One Health survey of rural residents’ knowledge, attitudes and practices in Shandong province, China”, Journal of Antimicrobial Chemotherapy73, no. 10 (2018): 2893-2899. https://doi.org/10.1093/jac/dky240
Prudius, T. Y., Gutsol, A. V., Gutsol, N. V., & Mysenko, O. O “Globigen Jump Start usage as a replacer for blood plasma in prestarter feed for piglets,” Scientific Messenger of LNU of Veterinary Medicine and Biotechnologies, Series: Agricultural sciences 23, no. 94 (2021): 111-116. https://doi.org/10.32718/nvlvet-a9420
Guevarra, Robin B., Jun Hyung Lee, Sun Hee Lee, Min-Jae Seok, Doo Wan Kim, Bit Na Kang, Timothy J. Johnson, Richard E. Isaacson and Hyeun Bum, “Piglet gut microbial shifts early in life: causes and effects,” Journal of animal science and biotechnology10, no. 1 (2019): 1-10. https://dx.doi.org/10.1186%2Fs40104-018-0308-3
Waititu, Samuel M., Jung M. Heo, Rob Patterson and Charles M. Nyachoti, “Dose-response effects of in-feed antibiotics on growth performance and nutrient utilization in weaned pigs fed diets supplemented with yeast-based nucleotides,” Animal Nutrition 1, no. 3 (2015): 166-169. https://doi.org/10.1016/j.aninu.2015.08.007
Xiaoyu Li, Ying Yao, Xitao Wang, Yuhong Zhen, Philip A Thacker, Lili Wang, Ming Shi, Junjun Zhao, Ying Zong, Ni Wang, Yongping Xu. “Chicken egg yolk antibodies (IgY) modulate the intestinal mucosal immune response in a mouse model of Salmonella typhimurium infection,” International immunopharmacology36, (2016) 305-314. https://doi.org/10.1016/j.intimp.2016.04.036
Yunsheng Han, Tengfei Zhan, Chaohua Tang, Qingyu Zhao, Dieudonné M Dansou, Yanan Yu, Fellipe F Barbosa, Junmin Zhang. Effect of Replacing in-Feed Antibiotic Growth Promoters with a Combination of Egg Immunoglobulins and Phytomolecules on the Performance, Serum Immunity, and Intestinal Health of Weaned Pigs Challenged with Escherichia coli K88. Animals 11, no. 5 (2021): 1292. https://doi.o
5 principles to consider when designing biosecurity programmes
Biosecurity is the foundation for all disease prevention programs and all the more important in antibiotic reduction scenarios. It includes the combination of all measures taken to reduce the risk of introduction and spread of diseases and is based on the prevention of and protection against infectious agents. Its fundament is the knowledge of disease transmission processes.
Although biosecurity is considered the cheapest and most effective intervention in antibiotic reduction programmes, compliance is often low and difficult.
The application of consistently high standards of biosecurity can substantially contribute to the reduction of antimicrobial resistance, not only by preventing the introduction of resistance genes into the farm but also by lowering the need to use antimicrobials.
Lower use of antimicrobials with higher biosecurity
Studies and assessments such as those done by (Laanen, et al., 2013), (Gelaude, et al., 2014), (Postma, et al., 2016), (Collineau, et al., 2017) and (Collineau, et al., 2017a) relate a high farm biosecurity or improvements in biosecurity with lower antimicrobial use. Laanen, Postma, and Collineau studied the profile of swine farmers in different European countries, finding a relation between a high level of internal biosecurity, efficient control of infectious diseases, and a reduced need for antimicrobials.
Others such as Gelaude and Collineau studied the effect of interventions. The former examined Belgian broiler farms, finding a reduction of antimicrobial use by almost 30% when biosecurity and other farm issues were improved within a year. The latter studied swine farms located in Belgium, France, Germany and Sweden, in which antimicrobial use was also reduced in 47% across all farms and observed that farms with the higher biosecurity compliance and who also took a holistic approach, making other changes (e.g. management and nutrition), achieved a higher reduction in antimicrobial use.
Biosecurity interventions pay off
Of course, the interventions necessary to achieve an increased level of biosecurity carry some costs. However, the interventions, especially if taken with other measures such as improved management of new-born animals and nutritional improvements, also improve productivity. The same studies which report that biosecurity improvements decrease antimicrobial use also report an improvement in animal performance. In the case of broilers, Laanen (2013) found a reduction of 0.5 percentual points in mortality and one point in FCR; and Collineau (2017) obtained an improvement during both the pre-weaning and the fattening period of 0.7 and 0.9 percentual points, respectively.
Implementation, application and execution
Although biosecurity is considered the cheapest and most effective intervention in antibiotic reduction programmes, compliance is often low and difficult. The implementation, application, and execution of any biosecurity programme involve adopting a set of attitudes and behaviours to reduce the risk of entrance and spread of disease in all activities involving animal production or animal care. Measures should not be constraints but part of a process aimed at improving the health of animals and people, and a piece of the holistic approach to reduce antibiotics and improve performance.
Designing effective biosecurity programmes: Consider these 5 principles
When designing or evaluating biosecurity programmes, we can identify 5 principles that need to be applied. These principles set the ground for considering and evaluating biosecurity interventions:
1. Separation: Know your enemy, but don’t keep it close
It is vital to have a good separation between high and low-risk animals or areas on the farm, as well as dirty (general traffic) and clean (internal movements) areas on the farm. This avoids not only the entrance but the spread of disease, as possible sources of infection (e.g. wild birds) cannot reach the sensitive population.
2. Reduction:Weaken your enemy, so it doesn’t spread
The goal of the biosecurity measures is to keep infection pressure beneath the level which allows the natural immunity of the animals to cope with the infections, lowering the pressure of infection e.g. by an effective cleaning and disinfection programme, by the reduction of the stocking density, and by changing footwear when entering a production house.
3. Focus:Hunt the elephant in the room, shoo the butterflies
In each production unit, some pathogens can be identified as of high economic importance. For each of these, it is necessary to understand the likely routes of introduction into a farm and how it can spread within it. Taking into account that not all disease transmission routes are equally important, the design of the biosecurity programme should focus first on high-risk transmission routes, and only subsequently on the lower-risk transmission routes.
4. Repetition: Increasing the probability of infection
In addition to the probability of pathogen transmission via the different transmission routes, the frequency of occurrence of the transmission route is also highly significant when evaluating a risk (Alarcon, et al., 2013). When designing biosecurity programmes, risky actions such as veterinary visits, if repeated regularly must be considered with a higher risk.
5. Scaling:In the multitude, it is easy to disguise
The risks related to disease introduction and spread are much more important in big; more animals may be infected and maintain the infection cycle, also large flocks/herds increase the infection pressure and increase the risk by contact with external elements such as feed, visitors, etc.
Can we still improve our biosecurity?
Almost 100% of poultry and swine operations already have a nominal biosecurity programme, but not in all cases is it effective or completely effective. BioCheck UGent, a standardised biosecurity questionnaire applied worldwide, shows an average of 65% and 68% of conformity, from more than 1000 broiler and 2000 swine farms between respectively; opportunities to improve can be found in farms globally, and they pay off.
The bottom line
Biosecurity is necessary for disease prevention in any profitable animal production system. To make effective plans, these 5 principles should be applied to choose the right interventions that prevent the entrance and spread of disease. However, maintaining a successful production unit requires a holistic approach in which other aspects of biosecurity need to also be taken seriously, as well as actions to improve in other areas such as management, health and nutrition.
Authors: Marisabel Caballero, Global Technical Manager Poultry – EW Nutrition, and Fellipe Freitas Barbosa, Team Lead Global Technical Management Swine – EW Nutrition
References available under request. Article published in Pig Progress.
Understanding and managing Strep suis in swine: The essentials
Strep suis causes vast losses in pig production and threatens human health, too. We still rely on antibiotics to control it – but we will have to change tactics to contain antimicrobial resistance.
Streptococcus suis is one of the most economically harmful pathogens for the global swine industry. When I started working in pig production 25 years ago, S. suis was already a problem on practically all the farms that I visited. Back then, our understanding of the pathogen and hence our control strategies were rudimentary: in farrowing rooms, we cut piglets’ teeth, used gentian violet spray on their navels, and sometimes applied penicillin lyophilized with iron. For the nursery phase, we only had penicillin or phenoxymethylpenicillin at our disposal – until the first amoxicillin-based premixes arrived, which turned out to be highly effective.
To this day, we control S. suis mainly through oral beta-lactam antibiotics (in feed or water) or injectable solutions, administered to piglets at an early age. However, pig production has evolved dramatically over the past decades, and so has the scientific research on this complex pathogen. Crucially, we now know that the excessive use of antibiotics contributes to the development of antimicrobial resistance.
Recent Australian research has discovered S. suis strains (both in humans and pigs) with a high degree of resistance to macrolides or tetracyclines, strains with intermediate sensitivity to Florfenicol, and others that are developing resistance to penicillin G. Additionally, we now know that S. suis is a zoonotic bacteria that affects not only at-risk farm or slaughterhouse personnel: S. suis is among the leading causes of death from meningitis in countries such as Thailand, China or Vietnam. In light of these threats to human health, we in the swine industry more than ever have a duty to help control this pathogen.
This article first reviews our current state of knowledge about the epidemiology and pathogenesis of Strep suis; it then lays out virulence factors and the role of coinfections. The second part considers the dimensions of a holistic approach to S. suis prevention and control and highlights the central role of intestinal health management.
What we know about S. suis epidemiology and pathogenesis
Practically all farms worldwide have carrier animals, but the percentage of animals colonized “intra-farm” varies between 40 and 80%, depending on several factors such as environmental conditions, hygiene measures, and the virulence of the S. suis strains involved.
How S. suis strains are classified
S. suis strains were once classified into 35 serotypes, according to their different capsular polysaccharides(CPS), theoutermost layer of the bacterial cell. Due to phylogenic and genomic sequencing, some of the old serotypes (20, 22, 26, 32, 33, and 34) are now reclassified, either in other bacterial genera or in other Streptococcus species. This has reduced the total to 29 S. suis serotypes.
Globally, the prevalence of the disease varies between 3% and 30%. The main serotypes affecting pig population are type 2 (28%), 9 (20%), and 3 (16%); differences in the geographical distribution are shown in Figure 1.
In addition to the serotype classification based on CPS antigens, S. suis has also been genetically differentiated into “sequence types” using the MLST (Multi Locus Sequence Typing) technique. The distribution of both porcine and human sequence types is detailed in Figure 2.
Figure 2: S. suis sequence types and their worldwide distribution
How S. suis is transmitted in swine
The main transmission routes are, firstly, the vertical sow-piglet route; the mucosa of the vagina is the first point of contamination. In the farrowing room, respiratory transmission from the sow to the piglets takes place. Horizontal transmission between piglets has also been proven to occur, especially during outbreaks in the post-weaning phase. This form of transmission happens through aerosols, feces, and saliva.
While in humans, the possibility of infection via the digestive tract has been confirmed, there are discussions about this route for swine. De Greeff et al. (2020) argue, based on in vitro and in vivo data, that infection through the digestive tract is associated with specific serotypes. Serotype 9, for example, would have a greater capacity for colonizing the gastrointestinal tract, and from there, the bacteria’s translocation takes place. The same authors point out that, in Western Europe, S. suis serotype 9 has become one of the most prevalent serotypes in recent years.
How S. suis colonization occurs
Although there are still unknown mechanisms in the pathogenesis of the disease, it can be schematically summarized how colonization occurs (Figure 3). From the different infection routes, the pathogen always passes through the mucosa. When S. suis enter the bloodstream, it can lead to a systemic infection, ending in septicemia, meningitis, endocarditis, or pneumonia, or a local infection at the joints level, causing arthritis.
According to Haas and Grenier (2018), different pathogenicity factors intervene in each of the processes. The CPS, for example, are relevant during colonization and the initial progression (indicated by black arrows). Microvesicles released by S. suis cell membranes are more involved in the passage to the bloodstream or, for example, the progression towards local or systemic infection (indicated by white arrows).
Depending on the host and the immune response, the well-known clinical signs of the disease will occur. Although they may begin in the lactation phase, the highest prevalence of meningitis (the main clinical symptom) usually occurs between the 5th and the 10th week of life, that is, between two and three weeks after weaning.
How to diagnose S. suis infection
Diagnosing S. suis is relatively simple at a clinical level; however, we need to know how to differentiate it from G. parasuis in the case of animals with nervous symptoms. We also need to distinguish S. suis from other pathogens responsible for producing arthritis, such as M. hyosynoviae or the fibrin-producing agent M. hyorhinis.
Laboratory techniques are developing on two fronts. Among molecular techniques, multilocus sequence typing (MLST) is considered the gold standard for serotyping. It is still costly and not yet practicable for large samples at the farm level. In contrast, several types of polymerase chain reaction (PCR) show greater practical applicability. Quantitative PCRs (qPCR) are used for the evaluation of bacterial load, and some PCRs are based on the identification of specific virulence genes.
Due to the relevance of S. suis for human health, more complex techniques are also available, such as the complete sequencing of the bacterial genome. This type of method aims to develop epidemiological analyzes together with the differentiation between virulent and non-virulent S. suis strains. Research is also underway in serology, particularly on evaluating maternal immunity and its interference with the piglet, as well as autogenous vaccines monitoring.
Why S. suis sometimes causes disease: Virulence factors and coinfections
Streptococcus suis is a pathobiont, i.e., a microorganism that belongs to the commensal flora of animals but generates disease under certain conditions. In their daily work on farms, clinical veterinarians, for instance, find that S. suis often colonizes the upper respiratory tract, nasal cavity, and tonsils without causing disease. S. suis pathogenicity is associated with an astounding range of different circumstances or triggering factors; some sources list more than 100 virulence factors. Several factors are considered essential in the development of pathogenesis; others, however, are the subject of ongoing research (cf. Xia et al., 2019, and Segura et al., 2017).
Critical virulence factors
One of the most important proteins is the CPS that establishes serotypes. The CPS largely determines the bacteria’s adhesion and colonization behavior. It can modify its thickness depending on the stage: it becomes thinner when adhering to the mucociliary apparatus and thicker when circulating through the bloodstream, protecting the bacteria against possible attacks by immune system cells.
Likewise, suis has an adhesin known as Protection Factor H (FHB) that protects it from phagocytosis by macrophages and can also interfere with the complement activation pathways of the immune system.
Suilysin is one of the most critical suis‘ protein toxins. This toxin plays a fundamental role in the interaction with host cells (modulating them to facilitate invasion and replication within the host cells) as well as in the inflammatory response.
S. suis is a mucosal pathogen and, hence, triggers a mucosal immunity response, mainly by immunoglobulins A (IgA). S. suis has developed proteases capable of destroying both IgA and IgG.
Research is still in progress, but both suis serotype 2 and 9 encode the development of adhesion proteins that facilitate mucociliary colonization when salivary glycoproteins are present (these are called antigens 1 and 2).
Other than Suilysin, two of the bacteria’s protein components that have been studied in-depth to develop subunit vaccines are the MRP (Muramidase Release Protein) and EF (Extracellular Factor) protein. Whether the expression of these proteins is associated with virulence depends on the serotype.
Recent research indicates that greater biofilm production capacity is associated with the more virulent suis strains. The production of biofilm is closely related to the production of fibrinogen, which allows the bacteria to develop resistance to the action of antimicrobials, to colonize tissues, to evade the immune system, etc.
Concomitant factors for S. suis infection
Even though S. suis is a primary pathogen that can cause disease by itself, many factors can exert a direct or indirect influence on whether or not and to which extent disease develops.
Veterinarians and producers are well aware of the influence of environmental and management factors such as temperature variations, poor ventilation together with poor air quality, irritants for the respiratory tract, as well as correct densities for animals’ welfare. Occasionally, depending on the geographical location, S. suis can be considered as a seasonal pathogen, showing a higher prevalence during the coldest months of the year when ventilation is lower or not well-controlled.
At the level of the individual animal, concomitant pathogens, environmental changes, diet changes, previous pathologies, piglet handling problems, etc., all come into play. Younger piglets tend to be more susceptible because of the decrease in maternal immunity or insufficient colostrum intake; diarrhea during the lactation phase also increases disease vulnerability.
Recently, researchers have started to explore the hypothesis that a change in the digestive tract microbiome balance may favor a pathogenic trajectory. Some results indicate that changes in the microbiota around the moment of weaning could indeed trigger disease. I will return to the vital topic of the digestive tract in S. suis pathogenesis below.
The role of coinfections
The virulence of S. suis can increase in the presence of other pathogens, both viral and bacterial. Among the main viruses, key interactants are the PRRS virus, the influenza virus (SIV), as well as Porcine Circovirus (PCV) and Porcine Respiratory Coronavirus (PRCV). At the bacterial level, Bordetella bronchiseptica and Glaesserella parasuis have the most direct interaction with S. suis (Brockmeier, 2020).
There are several mechanisms by which coinfections might increase S. suis virulence: some of them (i.e., B. bronchiseptica and SIV) alter the epithelial barrier, facilitating the translocation of S. suis. Moreover, viruses such as PRRS either cause an alteration in the response of the immune system or destroy relevant immune system cells.
Valentin-Weigand et al. (2020) posit that the influenza virus increases the pathogenic capacity of S. suis so that, for specific strains, the disease can develop even in the absence of the key virulence factor suilysin. This highlights the importance of controlling coinfections for successful S. suis management.
The five pillars of holistic S. suis management in swine
The challenge of managing this problematic pathogen with limited use of antibiotics prompts a review of all strategies within our reach. From birth to slaughterhouse, interventions must be coordinated and cannot work independently.
1. Biosecurity
The principles of biosecurity are easily understood. Yet, across different locations and production systems, farms struggle with consistently executing biosecurity protocols. For the moment, it appears unrealistic to avoid the introduction of new S. suis strains altogether. Also, complete eradication is not feasible with the currently available tools.
Genetic companies and research centers will likely continue to explore how to reduce bacterial colonization in animals, to produce piglets that have no or only minimal S. suis populations. Again, this option is not available for now.
At the farm level, the most promising and feasible approach is to reduce the risk of bacterial transmission, i.e., to optimize internal biosecurity. This extends to controlling both viral and bacterial coinfections. The two major viruses affecting the nursery stage are the PRRS virus and Swine Influenza virus. Bacteria that can contribute to the disintegration of the mucosa, both at the respiratory level and the digestive level, are Atrophic Rhinitis (progressive or not) and digestive pathogens such as E. coli, Rotavirus and Eimeria suis. All possible measures to reduce the prevalence and spread of these co-infectants must be executed to help control S. suis.
2. The pre-weaning period
We need to consider several elements in the first hours after birth that influence the spread of the bacteria in the farrowing rooms:
How is the colostrum distribution between the litters and the subsequent distribution of the piglets carried out?
How is the “processing” of the piglets carried out after farrowing: iron administration, wound management, and tail docking?
Are we taking any measure to prevent iatrogenic transmission of pathogens through needle exchange?
Until today, it is common practice to administer systemic (in-feed) or local (vaginally applied) antibiotics during the pre-weaning phase, albeit with partial or inconsistent successes in terms of reducing infection pressure. Notably, during the pre-weaning phase, the development of the piglet’s microbiota begins to take shape, and the systematic and prophylactic application of antibiotics in young animals can reduce bacterial diversity of the microbiome (Correa-Fiz et al., 2019). This, in turn, leads to a proliferation of bacteria with a pathogenic profile that could detrimentally influence subsequent pathology.
S. suis is an ultra-early colonizer; piglets can get infected at birth already
3. The post-weaning period
The post-weaning period undoubtedly constitutes the most critical stage of the piglets’ first weeks of life. In addition to social and nutritional stress, piglets are exposed to new pathogens. While maternal immunity is decreasing, piglets have not developed innate immunity yet; they are now most susceptible to the horizontal transmission of diseases. Hence, S. suis prevention during this phase center on measures that improve piglet quality. Key parameters include:
Do we have a correct and homogeneous weight/age ratio at weaning?
What is the level of anorexia in piglets? Do we practice suitable corrective measures to encourage the consumption of post-weaning feed?
How are we feeding them? What medications do they routinely receive?
How are housing facilities set up concerning density, environment, and hygiene?
Again, gut health is critical: Ferrando and Schultsz (2016) suggest that the status of the piglet’s weaning gastrointestinal tract centrally influences the subsequent development of the disease. Their research supports the idea that some specific S. suis serotypes can develop their pathogenesis from the digestive tract, just as in human medicine. While in humans, this digestive route is associated with the consumption of raw or insufficiently processed pork, in swine, the most susceptible moments are sudden changes in diet. The transition from milk to solid feed, in particular, leads to an increase in alpha-glucans that favor bacteria proliferation. Likewise, an increase in susceptibility occurs when the integrity of the intestinal wall is lost, for example, due to viral and bacterial coinfections.
4. Treatments and vaccination
Since weaning is such a difficult phase for the life of the piglet, it is a common practice on farms across the world to include one or several antibiotics in the post-weaning phase. Sometimes, when the legal framework allows, producers use a systematic antibiotic (i.e., beta-lactams or tetracyclines) and another one with a digestive profile (e.g., pharmacological doses of ZnO, trimethoprim, sulfa drugs and derivatives).
While antibiotics, for the most part, effectively prevent infection in the post-weaning phase, they can have adverse effects on the digestive tract. According to Zeineldin, Aldrige, and Lowe (2019), continued antibiotics use:
might increase the susceptibility to other infections because of the imbalance of the microbiome,
the immune system might be weakened, together with an alteration in metabolism,
and it fosters a greater accumulation of bacteria that are resistant to antibiotics.
The effectiveness of curative antibiotics treatments varies considerably. In any case, early detection is critical; affected animals need to be isolated and provided with a comfortable environment. Therapeutic parenteral antibiotics are best combined with high-dose corticosteroids. Some sick animals are unable to stand or walk. As a complementary measure, it is recommended, where possible, to help them ingest some feed and water.
Much research attention is focused on finding suitable vaccines to control the disease. This is a challenging task: S. suis shows high genetic diversity, making the identification of common proteins difficult, and is protected against antibody binding by a sugar-based envelope. The research group around Mariela Segura and Marcelo Gottschalk, for example, is working on a subunit vaccine strategy that addresses both dimensions. Recently, Arenas et al. (2019) identified infection-site specific patterns of S. suis gene expression, which could serve as a target for future vaccines.
The arrival of a universal, affordable S. suis vaccine is still a distant hope, though. Inactivated vaccines generally offer low levels of antibodies at the mucosal level and would need some adjuvant to increase them. A multiple injection protocol will not work from a commercial and practical point of view. On the other hand, live attenuated vaccines risk re-developing virulence with potentially drastic effects on human health. To complicate the topic of vaccination further, there is a controversy regarding the time of application and what animals we should vaccinate – sows, piglets, both?
Today, though with variable results, the alternative to scarce commercial vaccines is autogenous vaccines. These are based on the suspected serotype(s) present on a particular farm. This strategy hinges on the difficult procedure of isolating the strain from the meninges, spleen, or joints of the animals. If this step is successful, a laboratory can then develop the autogenous vaccine. Immunization occurs mainly in piglets, but occasionally some sows are vaccinated during the lactation period.
5. Hygiene
Just as for any other pathogen, hygiene management is critical. The infection pressure can be lowered through simple steps, such as washing the breeders before they enter the farrowing room. It is, or it should be, standard practice to maximize hygiene in the processing of piglets, avoiding injuries or pinching of the gums during teeth cutting, as well as disinfecting the umbilical area.
We know that S. suis is usually very sensitive to most disinfectants, but that is can form a biofilm that allows it to withstand hostile conditions. Physical or chemical methods to eliminate biofilm-formation are thus vital for combatting S. suis effectively.
Figure 4: The 5 pillars of S. suis control and prevention
S. suis control and prevention: The future lies in the gut
There is no ideal solution for totally controlling S. suis yet: autogenous vaccines are only partially effective, and since we cannot continue to administer antibiotics systematically, it is necessary to look for alternatives. Pending the arrival of a universal vaccine, the most promising efforts focus on the gastrointestinal tract.
Microbiome balance to keep S. suis in check
The gastrointestinal tract is not only the site where nutrient absorption takes place. The gut is the largest immune system organ in the body and most exposed to different antigens; therefore, what happens at the digestive level has a considerable influence on the immune system, locally and systemically.
The microbiome can be defined as the set of autochthonous bacteria that reside in the digestive system of animals. This group of bacteria is continually evolving and changes at critical moments in the life of animals. Simply put, a healthy microbiome is one that has a high bacterial diversity in the digestive tract (alpha diversity). The diversity between animals, on the other hand, should be low (beta diversity). A healthy microbiota implies the absence of dysbiosis and pathogens. Finally, one wants to promote the presence of bacteria that can produce substances with a bactericidal effect, such as short-chain fatty acids or bacteriocins.
Can we influence the microbiome to have fewer S. suis problems? Research by Wells, Aragon, and Bessems (2019) compared microbiota samples of the palatine tonsils from healthy and infected animals. They found that animals that would later develop the disease showed less diversity and, in particular, a diminished presence of the genus Moxarella. Importantly, they found that these differences in the microbiome’s composition of animals that later developed the disease were noticeable before weaning and at least two weeks before the outbreak occurred.
The same authors investigated in more depth, which bacteria in the microbiome were able to maintain homeostasis at the digestive level, finding that this was mostly the case for the genera Actinobacillus, Streptocuccus, and Moraxella. Moreover, they found that Prevotellacea and Rhotia produce antibacterial substances against S. suis.
Nutrition can impact the microbiome through targeted ingredients
We have to think about the microbiome of locations other than the digestive system as well. As we previously saw, the bacteria are transmitted through the mucosal route in the vagina, through the respiratory route, and there are recent studies that consider saliva as a leading source of infection in oral transmission.
This research contributes insights into how we might approach S. suis management through nutritional strategies. The question for nutritionists is, can you formulate feed that reduces the availability of S. suis’ favorite nutrients? S. suis appears to develop best when the feed contains large quantities of carbohydrates or starches. Other nutritional factors include the feed’s buffering capacity and the stomach pH of the piglets.
In times of antimicrobial resistance, additives are crucial for S. suis control and prevention
Gut health and nutrition approaches come together in the area of additives: targeted gut health-enhancing additives to feed or water will become a cornerstone of S. suis control. What we want to see in such products are molecules or substances that are capable of limiting, inhibiting, or slowing down the growth of S. suis by altering the membrane or interfering with the energy mechanisms of the bacteria.
There are already several products on the market with different active ingredients, such as phytomolecules, medium-chain fatty acids, organic acids, prebiotics, probiotics, etc. Soon, those products or combinations of them will be a part of our strategy for controlling this pathogen of such importance to our industry.
Author: Rafa Pedrazuela, Global Technical Manager Swine – EW Nutrition
References
Arenas, Jesús, Ruth Bossers-De Vries, José Harders-Westerveen, Herma Buys, Lisette M. F. Ruuls-Van Stalle, Norbert Stockhofe-Zurwieden, Edoardo Zaccaria, et al. “In Vivo Transcriptomes of Streptococcus Suis Reveal Genes Required for Niche-Specific Adaptation and Pathogenesis.” Virulence 10, no. 1 (2019): 334–51. https://doi.org/10.1080/21505594.2019.1599669.
Brockmeier, Susan L. “Appendix F – The role of concurrent infections in predisposing to Streptococcus suis and other swine diseases: Proceeding from the 4th International Workshop on S. suis.” Pathogens 9, no. 5 (2020): 374. https://doi.org/10.3390/pathogens9050374.
Correa-Fiz, Florencia, José Maurício Gonçalves Dos Santos, Francesc Illas, and Virginia Aragon. “Antimicrobial Removal on Piglets Promotes Health and Higher Bacterial Diversity in the Nasal Microbiota.” Scientific Reports 9, no. 1 (2019): Article number: 6545. https://doi.org/10.1038/s41598-019-43022-y.
De Greeff, Astrid, Xiaonan Guan, Francesc Molist, Manon Houben, Erik van Engelen, Ton Jacobs, Constance Schultsz et al. “Appendix A – Streptococcus suis serotype 9 infection: Novel animal models and diagnostic tools: Proceeding from the 4th International Workshop on S. suis.” Pathogens 9, no. 5 (2020): 374. https://doi.org/10.3390/pathogens9050374.
Ferrando, M. Laura, Peter Van Baarlen, Germano Orrù, Rosaria Piga, Roger S. Bongers, Michiel Wels, Astrid De Greeff, Hilde E. Smith, and Jerry M. Wells. “Carbohydrate Availability Regulates Virulence Gene Expression in Streptococcus Suis.” PLoS ONE 9, no. 3 (2014). https://doi.org/10.1371/journal.pone.0089334.
Ferrando, Maria Laura, and Constance Schultsz. “A Hypothetical Model of Host-Pathogen Interaction OfStreptococcus Suisin the Gastro-Intestinal Tract.” Gut Microbes 7, no. 2 (2016): 154–62. https://doi.org/10.1080/19490976.2016.1144008.
Goyette-Desjardins, Guillaume, Jean-Philippe Auger, Jianguo Xu, Mariela Segura, and Marcelo Gottschalk. “Streptococcus Suis, an Important Pig Pathogen and Emerging Zoonotic Agent—an Update on the Worldwide Distribution Based on Serotyping and Sequence Typing.” Emerging Microbes & Infections 3, no. 1 (2014): 1–20. https://doi.org/10.1038/emi.2014.45.
Haas, B., and D. Grenier. “Understanding the Virulence of Streptococcus Suis : A Veterinary, Medical, and Economic Challenge.” Médecine et Maladies Infectieuses 48, no. 3 (2018): 159–66. https://doi.org/10.1016/j.medmal.2017.10.001.
Murase, Kazunori, Takayasu Watanabe, Sakura Arai, Hyunjung Kim, Mari Tohya, Kasumi Ishida-Kuroki, Tấn Hùng Võ, et al. “Characterization of Pig Saliva as the Major Natural Habitat of Streptococcus Suis by Analyzing Oral, Fecal, Vaginal, and Environmental Microbiota.” Plos One 14, no. 4 (2019). https://doi.org/10.1371/journal.pone.0215983.
O’Dea, Mark A., Tanya Laird, Rebecca Abraham, David Jordan, Kittitat Lugsomya, Laura Fitt, Marcelo Gottschalk, Alec Truswell, and Sam Abraham. “Examination of Australian Streptococcus Suis Isolates from Clinically Affected Pigs in a Global Context and the Genomic Characterisation of ST1 as a Predictor of Virulence.” Veterinary Microbiology 226 (2018): 31–40. https://doi.org/10.1016/j.vetmic.2018.10.010.
Segura, Mariela, Nahuel Fittipaldi, Cynthia Calzas, and Marcelo Gottschalk. “Critical Streptococcus Suis Virulence Factors: Are They All Really Critical?” Trends in Microbiology 25, no. 7 (2017): 585–99. https://doi.org/10.1016/j.tim.2017.02.005.
Segura, Mariela, Virginia Aragon, Susan Brockmeier, Connie Gebhart, Astrid Greeff, Anusak Kerdsin, Mark O’Dea, et al. “Update on Streptococcus Suis Research and Prevention in the Era of Antimicrobial Restriction: 4th International Workshop on S. Suis.” Pathogens 9, no. 5 (2020): 374. https://doi.org/10.3390/pathogens9050374.
Tenenbaum, Tobias, Tauseef M Asmat, Maren Seitz, Horst Schroten, and Christian Schwerk. “Biological Activities of Suilysin: Role InStreptococcus Suispathogenesis.” Future Microbiology 11, no. 7 (2016): 941–54. https://doi.org/10.2217/fmb-2016-0028.
Valentin-Weigand, Peter, Fandan Meng, Jie Tong, Désirée Vötsch, Ju-Yi Peng, Xuehui Cai, Maren Willenborg et al. “Appendix G – Viral coinfection replaces effects of suilysin on adherence and invasion of Streptococcus suis into respiratory epithelial cells grown under air–liquid interface conditions: Proceeding from the 4th International Workshop on S. suis.” Pathogens 9, no. 5 (2020): 374. https://doi.org/10.3390/pathogens9050374.
Wells, Jerry, Virginia Aragon, and Paul Bessems. “Report on the deep analysis of the microbiota composition in healthy and S. suis-diseased piglets.” European Commission Program for Innovative Global Prevention of Streptococcus suis. Ref. Ares(2019)6305977, 2019. https://cordis.europa.eu/project/id/727966/results
Xia, Xiaojing, Wanhai Qin, Huili Zhu, Xin Wang, Jinqing Jiang, and Jianhe Hu. “How Streptococcus Suis Serotype 2 Attempts to Avoid Attack by Host Immune Defenses.” Journal of Microbiology, Immunology and Infection 52, no. 4 (2019): 516–25. https://doi.org/10.1016/j.jmii.2019.03.003.
Zeineldin, Mohamed, Brian Aldridge, and James Lowe. “Antimicrobial Effects on Swine Gastrointestinal Microbiota and Their Accompanying Antibiotic Resistome.” Frontiers in Microbiology 10 (2019). https://doi.org/10.3389/fmicb.2019.01035.
Zimmerman, Jeffrey J., Locke A. Karriker, Alejandro Ramirez, Kent J. Schwartz, Gregory W. Stevenson, and Jianqiang Zhang. Diseases of Swine. 11th ed. Hoboken, NJ: Wiley-Blackwell, 2019.
Necrotic enteritis: The complete overview
byInge Heinzl, Marisabel Caballero, Ajay Bhoyar, EW Nutrition
Eliminating necrotic enteritis from your operations starts from a good understanding of what it is, how to prevent it, and how to mitigate its effects on your poultry production.
Necrotic enteritis is a poultry disease caused by an overgrowth of Clostridium perfringens type A, and to a lesser extent type C, in the small intestine. The toxins produced by C. perfringens also damage the intestinal wall. In general, it occurs in broiler chickens of 2-6 weeks of age. In subclinical forms, it is characterized by impaired digestion. Clinical forms lead to severe problems and increased flock mortality in a very short time.
Necrotic enteritis is the cause of USD 6 billion annual losses in global poultry production and this controllable disease is on the rise. One reason is the voluntary or legally required reduction of antibiotics in animal production. This trend is driven by the increasing occurrence of antimicrobial resistance, as well as by consumer demand. Another reason is the reduction of ionophores which, besides their activity against coccidia, also show efficacy against clostridia. When anticoccidial live vaccines are used, the application of these ionophores is not possible and clostridia / necrotic enteritis increase (Williams, 2005).
While this is a widespread problem in all poultry, for broilers in particular, necrotic enteritis and coccidiosis are the most significant health problem.
Clinical and subclinical forms of NE
The clinical form
(c) Rob Moore
…is characterized by acute, dark diarrhea resulting in wet litter and suddenly increasing flock mortality of up to 1% per day after the first clinical signs appear (Ducatelle and Van Immerseel, 2010), sometimes summing up to mortality rates of 50% (Van der Sluis, 2013). The birds have ruffled feathers, lethargy, and inappetence.
Necropsy typically shows ballooned small intestines with a roughened mucosal surface, lesions, and brownish (diphtheritic) pseudo-membranes. There is a lot of watery brown, blood-tinged fluid and a foul odor during post-mortem examination. The liver is dark, swollen, and firm, and the gall bladder is distended (Hofacre et al., 2018).
In the case of peracute necrotic enteritis, birds may die without showing any preliminary signs.
The subclinical form
When birds suffer from the subclinical form, chronic damage to the intestinal mucosa and an increased quantity of mucus in the small intestine lead to impaired digestion and absorption of nutrients resulting in poor growth performance.
The deteriorated feed conversion and the resulting decreased performance become noticeable around day 35 of age. As feed contributes approximately 65-75% of the input cost to produce a broiler chicken, poor feed conversion increases production costs and significantly influences profitability. Often, due to a lack of clear symptoms, this subclinical disease remains untreated and permanently impacts the efficiency of production.
Pathogens
Responsible for necrotic enteritis are Gram-positive, anaerobic bacteria, specific strains of Clostridium perfringens type A and, to a lesser extent, type C (Keyburn et al., 2008).
Clostridia primarily occur in the soil where organic substances are degraded, in sewage, and the gastrointestinal tract of animals and humans. These bacteria produce spores, which are extremely resistant to environmental impact (heat, irradiation, exsiccation) as well as some disinfectants, and can survive for several years. Under suitable conditions, C. perfringens spores can even proliferate in feed or litter.
Clostridium perfringens is a natural inhabitant of the intestine of chickens. In healthy birds, it occurs in a mixture of diverse strains at 102-104 CFU/g of digesta (McDevitt et al., 2006). The disease starts when C. perfringens proliferates in the small intestine, usually due to a combination of factors such as high amount protein, low immunity, and an imbalance in the gut flora. Then, the number rises to 107-109 CFU/g of digesta (Dahiya et al., 2005).
NetB, a key virulence factor for NE
To establish in the host, Clostridium Spp. and other pathogens depend on virulence factors (see infobox). These virulence factors include, for example, “tools” for attachment, evasion or suppression of the host’s immune system, “tools” for getting nutrients, and “tools” for entry into intestinal cells. Over the years, the α-toxin produced by C. perfringens was assumed to be involved in the development of the disease and a key virulence factor. In 2008, Keyburn and coworkers found another key virulence factor by using a C. perfringens mutant unable to produce α-toxin, yet still causing necrotic enteritis.
Thus, another toxin was identified occurring only in chickens suffering from necrotic enteritis: C. perfringens necrotic enteritis B-like toxin (NetB). NetB is a pore-forming toxin. Pore-forming toxins are exotoxins usually produced by pathogenic bacteria, but may also be produced by other microorganisms. These toxins destroy the integrity of gut wall cell membranes. The leaking cell contents serve as nutrients for the bacteria. If immune cells are destroyed, an immune reaction might be partially impacted (Los et al., 2013).
Additionally, pathogenic strains of C. perfringens produce bacteriocins – the most important being Perfrin (Timbermont et al., 2014) – to inhibit the proliferation of harmless Clostridium Spp. strains and to replace the normal intestinal flora of chickens (Riaz et al., 2017).
Examples of virulence factors
1. Adhesins
Enable the pathogen to adhere or attach within the target host site, e.g. via fimbria. Pili enable the exchange of RNA or DNA between pathogens.
2. Invasion factors
Facilitate the penetration and the distribution of the pathogens in the host tissue (invasion and spreading enzymes). For example: hyaluronidase attacking the hyaluronic acid of the connective tissue or flagella enabling the pathogens to actively move.
3. Toxins
Damage the function of the host cells or destroy them (e.g. endotoxins – lipopolysaccharides, exotoxins)
4. Strategies of evasion
Enable the pathogen to bypass the strategies of defense of the immune system (e.g. antiphagocytosis factors provide protection against an attack by phagocytes; specific antibodies are inactivated by enzymes).
A chicken with optimal gut health may be less susceptible to NE. Additional predisposing factors are necessary to allocate nutrients and make the gut environment suitable for the proliferation of these pathogens, enabling them to cause disease (Van Immerseel et al., 2008; Williams, 2005).
Predisposing factors
Feed: composition and particle size
The role of feed in the development of necrotic enteritis should not be underestimated. This is where substances creating an intestinal environment favorable for C. perfringens come into play.
Mycotoxin contamination
Mycotoxins harm gut integrity and create ideal conditions for the proliferation of Clostridium perfringens.
Mycotoxins do not have a direct effect on C. perfringens proliferation, toxin production, or NetB transcription. However, mycotoxins disrupt gut health integrity, creating a favorable environment for the pathogen. For example:
DON provides good conditions for proliferation of C. perfringens by disrupting the intestinal barrier and damaging the epithelium. The possibly resulting permeability of the epithelium and a decreased absorption of dietary proteins can lead to a higher amount of proteins in the small intestine. These proteins may serve as nutrients for the pathogen (Antonissen et al., 2014).
DON and other mycotoxins decrease the number of lactic acid producing bacteria indicating a shift in the microbial balance (Antonissen et al., 2016.).
Eimeria ssp.
An intact intestinal epithelium is the best defense against potential pathogens such as C. perfringens. Here, Coccidiosis comes into play. Moore (2016) showed that by damaging the gut epithelium, Eimeria species give C. perfringens access to the intestinal basal domains of the mucosal epithelium. Then, the first phase of the pathological process takes place and from there, C. perfringens invades the lamina propria. Damage to the epithelium follows (Olkowski et al., 2008). The plasma proteins leaking to the gut and the mucus produced are rich nutrient sources (Van Immerseel et al., 2004; Collier et al., 2008). A further impact of Coccidiosis is shifting the microbial balance in the gut by decreasing the number of e.g., Candidatus savagella which activates the innate immune defense.
Eimeria induce leakage of plasma proteins by killing epithelial cells
They enhance mucus production in the intestine
1+2 lead to an increase in available nutrients and create an environment favorable for the proliferation of C. perfringens.
Not only Eimeria Spp., also other pathogens (e.g. Salmonella Spp., Ascarid larvae, viruses) and agents, such as mycotoxins damaging the intestinal mucosa can pave the way for a C. perfringens infection.
Predisposing factors like wet litter, the moisture of which is essential for the sporulation of Eimeria Spp. oocysts, must also be considered as promoting factors for necrotic enteritis (Williams, 2005).
Immunosuppressive factors
Besides the already explained influencers feed, mycotoxins and coccidia, also other predisposing factors must be mentioned. In general, any factor which induces stress in the animals disrupts the balance of the intestinal flora. The resulting suppression of the immune system contributes to the risk of necrotic enteritis (Tsiouris, 2016). These factors include:
Bacteria: Shivaramaiah and coworkers (2011) investigated a neonatal Salmonella typhimurium infection as a predisposing factor for NE. The early infection causes significant damage to the gut (Porter et al., 1998) Additionally, Hassan et al. (1994) showed that the challenge with Salmonella typhimurium negatively impacted the development of lymphocytes which might also promote a colonization of Clostridium perfringens.
Viruses: Infectious Bursal Disease is known to increase the severity of infections with salmonella, staphylococci, but also clostridia. Another clostridia-promoting viral disease is Marek’s Disease.
Stress: The intestinal tract is particularly sensitive to any type of stress. This stress can be caused by e.g. too high temperatures, high stocking densities, an abrupt change of feed.
Treatment
In acute cases, the farmer should consult a veterinarian and treat his birds.
It must be mentioned that, as the treatment takes place via feed or water, only birds which still consume water or feed may be treated.
Antibiotics
Antibiotics targeting Gram-positive bacteria are commonly used for the treatment of acute NE. The antibiotic choice shall be addressed by a veterinarian, taking into account mode of action and the presence of resistance genes in the farm/flock.
The prophylactic use of antibiotics is not recommened and many countries have already banned it in order to reduce antimicrobial resistance (AMR).
Antimicrobial Resistance (AMR)
Some bacteria are less sensitive to certain antibiotics due to genetic mutations. They are able to:
stimulate the production of enzymes, which break down or modify the antibiotics and inactivate them (1).
eliminate entrances for antibiotics or promote the development of pumps, which discharge the antibiotic before taking effect (2).
change or eliminate molecules to which the antibiotic would bind (targets for the antibiotics).
This means that, when the corresponding antibiotics are used, bacteria resistant against these antibiotics survive. Due to the fact that their competitors have been eliminated they are able to reproduce better. Additionally, this resistance may be transferred by means of “resistance genes”
to daughter cells
via their intake from dead bacteria (3)
through horizontal gene transfer (4)
through viruses (5)
Every application of antibiotics promotes the development of resistance (Robert Koch Institute, 2019). A short-term use, better biosecurity, or an application at low dosage give the bacteria a better chance to adapt.
Bacteriophages
Experimental use of phage treatments have shown to be effective in reducing disease progression and symptoms of necrotic enteritis (Miller et al., 2010). By oral application of a bacteriophage cocktail, Miller and coworkers could reduce mortality by 92% in C. perfringens challenged broilers compared to the untreated control.
Mode of action: the endolysins, highly evolved enzymes produced by bacteriophages, are able to digest the bacterial cell wall for phage progeny release (Fischetti, 2010). However, phages are still not approved by the EFSA.
Prevention
Preventing a disease is always better – and more cost-effective – that its treatment.
How, then, should it be done?
Preventing the conditions that favor the proliferation of Clostridium perfringens and strengthening the host’s immune response lowers the probability of disease.
Besides eliminating the predisposing factors, the main targets are:
There is evidence that most Clostridium strains isolated from birds suffering from necrotic enteritis could induce the disease experimentally, while strains isolated from healthy birds cannot. This confirms that only specific strains are problematic (Ducatelle and Van Immerseel, 2010).
It is therefore of the highest importance to avoid introducing these pathogenic strains to the farm.
Separate clothing, boots, and hand washing/disinfecting facilities in each poultry house
More than 14 days of down time between flocks
Specific measures against coccidiosis
Vaccination
According to parasitologists, 7 to 9 Eimeria species are found in chickens, and they do not cross-protect against each other. An effective vaccination must contain sporulated oocysts of the most critical pathogenic Eimeria species (E. acervulina, E. maxima, E. tenella, E. necatrix, and E. brunetti). The more species contained in the vaccine, the better. However, if not applied the correct way, vaccines can be ineffective or cause reactions in the birds that might lead to NE (Mitchell, 2017).
Anticoccidials
Alternate use of chemicals (synthetic compounds) and ionophores (polyether antibiotics) with different modes of action is important to avoid development of resistance.
Ionophores have a specific mode of action and kill oocysts before they are able to infect birds. Being very small, ionophore molecules can be taken up and diffused into the outer membrane of the sporozoite. There, it decreases the concentration gradient leading to an accumulation of water within the sporozoite causing its bursting.
Diet
Minimizing non-starch polysaccharides (NSPs) in cereals
To prevent a “feeding” of Clostridium perfringens, high content of water-soluble but indigestible NSPs such as wheat, wheat by-products, and barley should be avoided or at least minimized. Additionally, xylanases should be included in the feed formulation to reduce the deleterious effects of NSPs and improve feed energy utilization. Instead of these cereals, maize could be included in the diet. It is considered a perfect ingredient in broiler diets due to its high energy content and high nutrient availability.
Formulating low protein diets/diets with highly digestible amino acids
Feeding low-protein diets supplemented with crystalline amino acids might be beneficial to reduce the risk of necrotic enteritis (Dahiya et al., 2007). To improve protein digestibility and therefore reduce the proliferation of C. perfringens, proteases may be added to the feed.
Avoiding/Minimizing poor quality fats / animal fats in the diet
These fats tend to increase the count of Clostridium perfringens; thus, they should be replaced by higher quality and/or vegetable fats, respectively.
Feed form
In terms of feed form, Engberg et al. (2002) found that birds fed pellets showed a reduced number of Clostridium perfringens in the caeca and the rectum than mash-fed birds. Branton and co-workers (1987) reported a lower mortality by feeding roller-milled (coarsely ground) than hammer-milled feed.
Additives
Additives can be used either to prevent the proliferation of Clostridium perfringens or to change the environmental conditions in a way that proliferation of C. perfringens is prevented.
Probiotics
These live microbial supplements can be used to help to establish, maintain or re-establish the intestinal microflora.
Mode of action:
compete with pathogenic bacteria for substrates and attachment sites
produce antimicrobial substances inhibiting the growth of pathogenic bacteria (Gillor et al., 2008)
bind and neutralize enterotoxins (Mathipa and Thantsha, 2017)
promote immune function of the host (Yang et al., 2012)
Prebiotics
These feed ingredients serve as substrates to promote beneficial bacteria in the intestine.
Mode of action:
D-mannose or fructose, starches non-digestible by birds, selectively stimulate the growth and the activity of the “good” gut flora
Fructooligosaccharides decrease C.perfringens and E. coli in the gut and increase the diversity of Lactobacillus Spp. (Kim et al., 2011)
Galactooligosaccharides, in combination with a B. lactis based probiotic, have been reported to selectively promote the proliferation of Bifidobacterium ssp. (Jung et al., 2008).
Organic acids
Organic acids are often used in animal diets to improve intestinal health.
Mode of action:
decreased pH promotes beneficial bacteria
caprylic acid suppresses C. perfringens, but also Salmonella Spp. by inhibiting their utilization of glucose (Skrivanova et al., 2006)
lauric, citric, oleic and linoleic acid as well as medium-chain fatty acids (C8-C14) impede the growth of C. perfringens
Phytomolecules
Phytomolecules, also known as secondary plant compounds, have been used against pathogens for centuries. In general, two subgroups of these substances are known as effective against Clostridium perfringens:
Tannins
Many studies have shown the efficacy of tannins against different pathogens such as helminths, Eimeria, viruses, and bacteria
Extracts from the chestnut and quebracho trees are effective not only against C. perfringens, but also its toxins (Elizando et al., 2010)
Activity of tannins against Eimeria (Cejas et al., 2011) and Salmonella Sp., two predisposing factors for NE.
Essential Oils
Their hydrophobic characteristic enables them to interact with the lipids of the membrane of C. perfringens.
They can incorporate into the bacterial membrane and disrupt its integrity.
This increases the permeability of the cell membrane for ions and other small molecules such as ATP, leading to the decrease of the electrochemical gradient above the cell membrane and the loss of the cell’s energy equivalents.
Besides their direct effect on Clostridium Spp., a lot of phytomolecules improve gut health and help to prevent a proliferation of Clostridium ssp. and therefore necrotic enteritis.
Mycotoxin/bacterial toxin binders
These binders have two modes of action:
Binding mycotoxins, damage of the intestinal epithelium can be reduced or even prevented, so that the preconditions for Clostridium proliferation are not generated.
Binding toxins produced by Clostridium perfringens can reduce the occurrence or severity of lesions:
Alpha-toxin (phospholipase C) hydrolyses membrane phospholipids and damages erythrocytes, leucocytes, myocytes, and endothelial cells and causes their lysis (Songer, 1996). This leads to necrosis and tissue damage.
Binding NetB toxin, the key virulence factor, could reduce the severity of necrotic enteritis.
Conclusion
The ever-growing trend of reduced antibiotic and ionophore use is contributing to an increased incidence of necrotic enteritis in poultry production.
The subclinical form of necrotic enteritis generally goes unnoticed, resulting in poor feed efficiency and is a major cause of financial losses to poultry producers.
Maintaining optimum gut health is key to preventing the occurrence of necrotic enteritis. In the era of antibiotic-free poultry production, alternatives acting against this pathogenic bacterium and also against its predisposing factors must be considered to control this devastating disease.
References
Annett, C.B., J. R. Viste, M. Chirino-Trejo, H. L. Classen, D. M. Middleton, and E. Simko. “Necrotic enteritis: effect of barley, wheat and corn diets on proliferation of Clostridium perfringens type A.” Avian Pathology 31 (2002): 599– 602. https://doi.org/10.1080/0307945021000024544
Antonissen G, F. Van Immerseel, F. Pasmans, R. Ducatelle, F. Haesebrouck, L. Timbermont, M. Verlinden, G.P.J. Janssens, V. Eeckhaut, M. Eeckhout, S. De Saeger, S. Hessenberger, A. Martel, and S. Croubels. “The mycotoxin deoxynivalenol predisposes for the development of Clostridium perfringens-Induced necrotic enteritis in broiler chickens. PLoS ONE 9 no. 9 (2014): e108775. https://doi.org/10.1371/journal.pone.0108775
Antonissen, G., V. Eeckhaut, K. Van Driessche, L. Onrust , F. Haesebrouck, R. Ducatelle, R.J. Moore, and F. Van Immerseel. “Microbial Shifts Associated With Necrotic enteritis.” Avian Pathol. 45 no. 3 (2016): 308-312. https://doi.org/10.1080/03079457.2016.1152625
Branton, S.L., F.N. Reece, and W.M. Hagler. “Influence of a wheat diet on mortality of broiler chickens associated with necrotic enteritis.” Poultry Sci. 66 (1987): 1326-1330. https://doi.org/10.3382/ps.0661326
Cejas, E., S. Pinto, F. Prosdócimo, M. Batalle, H. Barrios, G. Tellez, and M. De Franceschi. “Evaluation of quebracho red wood (Schinopsis lorentzii) polyphenols vegetable extract for the reduction of coccidiosis in broiler chicks.” International Journal of Poultry Science 10 no. 5 (2011): 344–349. https://doi.org/10.3923/ijps.2011.344.349
Collier, C.T., C.L. Hofacre, A.M. Payne, D.B. Anderson, P. Kaiser, R.I. Mackie, and H.R. Gaskins. “Coccidia-induced mucogenesis promotes the onset of necrotic enteritis by supporting Clostridium perfringens growth.” Veterinary Immunology and Immunopathology 122 (2008):104–115.
Dahiya, J.P., D. Hoehler, A.G. Van Kessel, and M.D. Drew. “Effect of different dietary methionine sources on intestinal microbial populations in broiler chickens.” Poultry Science 86 (2007):2358–2366
Dahiya, J.P., D. Hoehler, D.C. Wilkie, A.G. van Kessel, and M.D. Drew. “Dietary glycine concentration affects intestinal Clostridium perfringens and Lactobacilli populations in broiler chickens.” Poultry Science 84 no.12 (2005):1875-85. https://doi.org/10.1093/ps/84.12.1875
Diaz Carrasco, J.M., L.M. Redondo, E.A. Redondo, J.E. Dominguez, A.P. Chacana, and M.E. Fernandez Miyakawa. “Use of plant extracts as an effective manner to control Clostridium perfringens induced necrotic enteritis in poultry.” BioMed Research International (2016): Article ID 3278359. https://dx.doi.org/10.1155/2016/3278359
Ducatelle, R. and F. van Immerseel. “Necrotic enteritis: emerging problem in broilers.” WATTAgNet.com – Poultry Health and Disease (April 9, 2010).
Elizondo, A.M., E.C. Mercado, B.C. Rabinovitz, and M.E. Fernandez-Miyakawa. “Effect of tannins on the in vitro growth of Clostridium perfringens.” Veterinary Microbiology 145 no. 3-4 (2010): 308–314. https://doi.org/10.1016/j.vetmic.2010.04.003
Engberg, R.M., M.S. Hedemann, and B.B. Jensen. “The influence of grinding and pelleting of feed on the microbial composition and activity in the digestive tract of broiler chickens.” · British Poultry Science 43 no. 4 (2002):569-579. https://doi.org/10.1080/0007166022000004480
Fischetti, V.A. “Bacteriophage endolysins: A novel anti-infective to control Gram-positive pathogens.” J Med Microbiol. 300 no. 6 (2010): 357–362. https://doi.org/10.1016/j.ijmm.2010.04.002
Gillor, O., A. Etzion and M.A. Riley. “The dual role of bacteriocins as anti- and probiotics.” Appl Microbiol Biotechnol. 81 no. 4 (2008): 591–606. https://doi.org/10.1007/s00253-008-1726-5
Hassan, J. O., and R. Curtiss III. “Virulent Salmonella typhimurium induced lymphocyte depletion and immunosuppression in chickens.” Infect. Immun. 62 (1994):2027–2036 https://doi.org/10.1128/IAI.62.5.2027-2036.1994
Hofacre, C.L., J.A. Smith, and G.F. Mathis. “Invited Review. An optimist’s view on limiting necrotic enteritis and maintaining broiler gut health and performance in today’s marketing, food safety, and regulatory climate.” Poultry Science 97 (2018):1929–1933. https://dx.doi.org/10.3382/ps/pey082
Jung, S.J., R. Houde, B. Baurhoo, X. Zhao, and B. H. Lee. “Effects of galacto-oligosaccharides and a bifidobacteria lactis-based probiotic strain on the growth performance and fecal microflora of broiler chickens.” Poultry Science 87 (2008):1694–1699. https://doi.org/10.3382/ps.2007-00489
Kaldhusdal and Skjerve. “Association between cereal contents in the diet and incidence of necrotic enteritis in broiler chickens in Norway.” Preventive Veterinary Medicine 28 (1996):1-16. https://doi.org/10.1016/0167-5877(96)01021-5
Keyburn, A. L., S. A. Sheedy, M. E. Ford, M. M. Williamson, M. M. Awad, J. I. Rood, and R. J. Moore. “Alpha-toxin of Clostridium perfringens is not an essential virulence factor in necrotic enteritis in chickens.” Infect. Immun. 74 (2006): 6496–6500. https://doi.org/10.1128/IAI.00806-06
Keyburn, A.L., J.D. Boyce, P. Vaz, T.L. Bannam, M.E. Ford, D. Parker, A. Di Rubbo, J.I. Rood, and R.J. Moore. “NetB, a new toxin that is associated with avian necrotic enteritis caused by Clostridium perfringens.” PLoS Pathog 4 no. 2, e26 (2008): 0001-0011. https://doi.org/10.1371/journal.ppat.0040026
Kim, G.-B., Y. M. Seo , C. H. Kim , and I. K. Paik. “Effect of dietary prebiotic supplementation on the performance, intestinal microflora, and immune response of broilers.” Poultry Science 90 (2011):75–82. https://doi.org/10.3382/ps.2010-00732
Knap, I., B. Lund, A. B. Kehlet, C. Hofacre, and G. Mathis. “Bacillus licheniformis prevents necrotic enteritis in broiler chickens.” Avian Diseases 54 no. 2 (2010):931-935. https://doi.org/10.1637/9106-101509-ResNote.1
Knarreborg, A., M.A. Simon, R.M. Engberg, B.B. Jensen, and G.W. Tannock. “Effects of Dietary Fat Source and Subtherapeutic Levels of Antibioticon the Bacterial Community in the Ileum of Broiler Chickensat Various Ages.” Applied and Environmental Microbiology 68 no. 12 (2002): 5918-5924. https://doi.org/0.1128/AEM.68.12.5918–5924.2002
Kocher, A. and M. Choct. “Improving broiler chicken performance. The efficacy of organic acids, prebiotics and enzymes in controlling necrotic enteritis.” Australian Government-Rural Industries Research and Development Corporation. Publ. no. 08/149 (2008).
Kondo, F. “In vitro lecithinase activity and sensitivity to 22 antimicrobial agents of Clostridium perfringens isolated from necrotic enteritis of broiler chickens.” Research in veterinary Science 45 (1988): 337-340. https://doi.org/10.1016/S0034-5288(18)30961-5
Kubena, L.F., J.A. Byrd, C.R. Young, and D.E. Corrier. “Effects of tannic acid on cecal volatile fatty acids and susceptibility to Salmonella typhimurium colonization in broiler chicks.” Poultry Science 80, no. 9 (2001): 1293–1298. https://doi.org/10.1093/ps/80.9.1293
Los, F.C.O., T.M. Randis, R.V. Aroian, and A.J. Ratner. “Role of pore-forming toxins in bacterial infectious diseases.” Microbiology and Molecular Biology Reviews 77 (2013): 173-207 https://doi.org/10.1128/MMBR.00052-12
M’Sadeq S.A., Shubiao Wu, Robert A. Swick, Mingan Choct. “Towards the control of necrotic enteritis in broiler chickens with in-feed antibiotics phasing-out worldwide.” Animal Nutrition 1 (2015): 1-11. https://dx.doi.org/10.1016/j.aninu.2015.02.004
Mathipa, M.G. and M.S. Thantsha. “Probiotic engineering: towards development of robust probiotic strains with enhanced functional properties and for targeted control of enteric pathogens.” Gut Pathog. 9 no. 28 (2017). https://doi.org/10.1186/s13099-017-0178-9
McDevitt, R.M., J.D. Brooker, T. Acamovic, and N.H.C. Sparks. “Necrotic enteritis, a continuing challenge for the poultry industry.” World’s Poultry Science Journal 62; World’s Poultry Science Association (June 2006). https://doi.org/10.1079/WPS200593
Miller, R.W., J. Skinner, A. Sulakvelidze, G.F. Mathis, and C.L. Hofacre. “Bacteriophage therapy for control of necrotic enteritis of broiler chickens experimentally infected with Clostridium perfringens.” Avian Diseases 54 no. 1 (2010): 33-40. https://doi.org/10.1637/8953-060509-Reg.1
Mitsch, P., K. Zitterl-Eglseer, B. Köhler, C. Gabler, R. Losa, and I. Zimpernik. “The Effect of Two Different Blends of Essential Oil Components on the Proliferation of Clostridium perfringens in the Intestines of Broiler Chickens.” Poultry Science 83 (2004):669–675. https://doi.org/10.1093/ps/83.4.669
Olkowski, A.A., C. Wojnarowicz, M. Chirino-Trejo, B. Laarveld, and G. Sawicki. “Sub-clinical necrotic enteritis in broiler chickens: Novel etiological consideration based on ultra-structural and molecular changes in the intestinal tissue.” Veterinary Science 85 (2008): 543–553. https://doi.org/10.1016/j.rvsc.2008.02.007
Pan, D. and Z. Yu. “Intestinal microbiome of poultry and its interaction with host and diet.” Gut Microbes 5 no. 1 (2014): 108–119. https://dx.doi.org/10.4161/gmic.26945
Rougière, N. and B. Carré. “Comparison of gastrointestinal transit times between chickens from D + and D- genetic lines selected for divergent digestion efficiency.” Animal 4 no. 11 (2010): 1861-1872. https://doi.org/10.1017/S1751731110001266
Santos, F.B.O., B.W. Sheldon, A.A. Santos Jr., and P.R. Ferket. ”Influence of housing system, grain type, and particle size on Salmonella colonization and shedding of broilers fed triticale or corn-soybean meal diets.” Poultry Science 87 (2008): 405-420. https://dx.doi.org/10.3382/ps.2006-00417
Schiavone, A. , K. Guo, S. Tassone, L .Gasco, E. Hernandez, R. Denti, and I. Zoccarato. “Effects of a Natural Extract of Chestnut Wood on Digestibility, Performance Traits, and Nitrogen Balance of Broiler Chicks.” Poult Sci. 87 no. 3 (2008): 521-527. https://doi.org/10.3382/ps.2007-00113
Shivaramaiah, S., R. E. Wolfenden, J. R. Barta, M. J. Morgan, A. D. Wolfenden, B. M. Hargis, and G. Téllez. „The role of an early Salmonella typhimurium infection as a predisposing factor for necrotic enteritis in a laboratory challenge model.” Avian Diseases 55 (2011): 319-323. https://doi.org/10.1637/9604-112910-ResNote.1
Singh, Y., V. Ravindran, T.J. Wester, A.L. Molan, and G. Ravindran. “ Influence of feeding coarse corn on performance, nutrient utilization, digestive tract measurements, carcass characteristics, and cecal microflora counts of broilers.” Poultry Science 93 (2014): 607–616. https://dx.doi.org/10.3382/ps.2013-03542
Skrivanova, E., M. Marounek, V. Benda, and P. Brezina. “Susceptibility of Escherichia coli, Salmonella sp. and Clostridium perfringens to organic acids and monolaurin.” Veterinarni Medicina 51 no. 3 (2006): 81–88. https://doi.org/10.17221/5524-VETMED
Stanley D., Wu S.-B., Rodgers N., Swick R.A., and Moore R.J. “Differential Responses of Cecal Microbiota to Fishmeal, Eimeria and Clostridium perfringens in a Necrotic Enteritis Challenge Model in Chickens.” PLoS ONE 9 no. 8 (2014): e104739. https://doi.org/10.1371/journal.pone.0104739
Tan, L., D. Rong, Y. Yang, and B. Zhang. “Effect of Oxidized Soybean Oils on Oxidative Status and Intestinal Barrier Function in Broiler Chickens.” Brazilian Journal of Poultry Science 18 no. 2 (2018): 333-342. http://dx.doi.org/10.1590/1806-9061-2017-0610
Tan, L., D. Rong, Y. Yang, and B. Zhang. “The Effect of Oxidized Fish Oils on Growth Performance, Oxidative Status, and Intestinal Barrier Function in Broiler Chickens.” J. Appl. Poult. Res. 28 (2019): 31-41. http://dx.doi.org/10.3382/japr/pfy013
Timbermont L., A. Lanckriet, J. Dewulf, N. Nollet, K. Schwarzer, F. Haesebrouck, R. Ducatelle, and F. Van Immerseel. “Control of Clostridium perfringens-induced necrotic enteritis in broilers by target-released butyric acid, fatty acids and essential oils.” Avian Pathol. 39 no. 2 (2010): 117-21. https://doi.org/10.1080/03079451003610586
Tsiouris, V. “Poultry management: a useful tool for the control of necrotic enteritis in poultry.” Avian Pathol. 45 no. 3 (2016):323-325. https://doi.org/10.1080/03079457.2016.1154502
Van der Most, P.J., B. de Jong, H.K. Parmentier and S. Verhulst. “Trade-off between growth and immune function: a meta-analysis of selection experiments.” Functional Ecology 25 (2011): 74-80. https://doi.org/0.1111/j.1365-2435.2010.01800.x
Van der Sluis, W. “Clostridial enteritis is an often underestimated problem.” Worlds Poult. Sci. J. 16 (2000):42–43.
Van Immerseel, F., J. De Buck, F. Pasmans, G. Huyghebaert, F. Haesebrouck, and R. Ducatelle. “Clostridium perfringens in poultry: an emerging threat of animal and public health.” Avian Pathology 33 (2004): 537-549. https://doi.org/10.1080/03079450400013162
Van Immerseel, F., J.I. Rood, R.J. Moore, and R.W. Titball. “Rethinking our understanding of the pathogenesis of necrotic enteritis in chickens.” Trends in Microbiology 17 no. 1 (2008):32-36. https://doi.org/10.1016/j.tim.2008.09.005
Wade, B., A.L. Keyburn, T. Seemann, J.I. Rood, and R.J. Moore. “Binding of Clostridium perfringens to collagen correlates with the ability to cause necrotic enteritis in chickens.” Veterinary Microbiology 180 no. 3–4 (2015): 299-303. https://doi.org/10.1016/j.vetmic.2015.09.019
Williams, R.B. “Intercurrent coccidiosis and necrotic enteritis of chickens: rational, integrated disease management by maintenance of gut integrity.” Avian Pathology 34 no. 3 (2005):159-180. https://doi.org/10.1080/03079450500112195
Yang , C.M., G.T. Cao, P.R. Ferket, T.T. Liu, L. Zhou, L. Zhang, Y.P. Xiao, and A. G. Chen. “ Effects of probiotic, Clostridium butyricum, on growth performance, immune function, and cecal microflora in broiler chickens.” Poultry Science 91 (2012): 2121–2129. https://dx.doi.org/10.3382/ps.2011-02131
Are endotoxins behind your low livestock productivity?
Impaired health status of the animals in stressful situations or an aggravation of the disease after antibiotic treatment? The culprit might be endotoxins.
What are endotoxins?
Origin
Endotoxins, together with exotoxins, are bacterial toxins. In contrast to exotoxins, which are actively secreted by living bacteria, endotoxins (name “endotoxin” greek; endo = inside; toxin = poison) are components of the outer cell membrane of gram-negative bacteria such as Escherichia coli, Salmonella, Shigella, and cyanobacteria (blue-green algae). They are only released in case of
bacterial death due to effective host defense mechanism or activities of certain antibiotics
Biochemically, endotoxins are lipopolysaccharides (LPS). They are composed of a relatively uniform lipid fraction (Lipid A) and a species-specific polysaccharides chain. Their toxicity is mainly due to the lipid A; the polysaccharide part modifies their activity. Unlike the bacteria, their endotoxins are very heat stable and resist sterilization. The names endotoxin and lipopolysaccharides are used synonymously with “endotoxin” emphasizing on the occurrence and biological activity and “lipopolysaccharide” on the chemical structure (Hurley, 1995).
General structure of Gram-negative lipopolysaccharides (according to Erridge et al., 2002)
Impact
Endotoxins belong to the so-called pyrogen-agents (they provoke fever), activating several immunocompetent cells’ signaling pathways. Early contact with endotoxins leads to activation and maturation of the acquired immune system. Braun-Fahrländer and co-workers (2002) found that children exposed to endotoxins had fewer problems with hay fever, atopic asthma, and atopic sensitization. This might be an explanation that in human populations, after the elevation of the hygiene standards, an increase of allergies could be observed.
Different animal species show different sensibilities to endotoxin infusions, e.g. (healthy) dogs, rats, mice, hens tolerate concentrations ≥1mg / kg body weight, whereas (healthy) ruminants, pigs, horses react very sensible already at concentrations <5μg / kg body weight (Olson et al., 1995 cited in Wilken, 2003).
Reasons for increased exposure of the organism to endotoxins
Endotoxins usually occur in the gut, as the microflora also contains gram-negative bacteria. The precondition for endotoxins to be harmful is their presence in the bloodstream. In the bloodstream, low levels of endotoxins can still be handled by the immune defense, higher levels can get critical. An increase of endotoxins in the organism results from higher input and/or lower clearance or detoxification rate.
Higher input of endotoxins into the organism
The “normal” small amounts of endotoxins arising in the gut due to regular bacterial activity and translocated to the organism have no negative impact as long as the liver performs its clearance function. Also, the endotoxins stored in the adipose tissue are not problematic. However, some factors can lead to a release of the endotoxins or translocation of endotoxins into the organism:
1. Stress
Stress situations such as parturition, surgeries, injuries can lead to ischemia in the intestinal tract and translocation of endotoxins into the organism (Krüger, 1997). Other stress situations in animal production, such as high temperatures and high stocking densities, contribute to higher endotoxin levels in the bloodstream. Stress leads to a higher metabolic demand for water, sodium, and energy-rich substances. For a higher availability of these substances, the intestinal barrier’s permeability is increased, possibly leading to a higher translocation of bacteria and their toxins into the bloodstream.
Examples:
Higher levels of endotoxins in pigs in an experimental study suffering from stress due to loading and transport, elevated temperatures (Seidler (1998) cited in Wilken (2003)).
Marathon runners (Brock-Utne et al., 1988) and racing horses (Baker et al., 1988) also showed higher endotoxin concentrations in the blood proportional to the running stress; thus, trained horses showed lower concentrations than untrained.
2. Lipolysis for energy mobilization
If endotoxins, due to continuous stress, consistently get into the bloodstream, they can be stored in the adipose tissue. The SR-B1 (Scavenger receptor B1, a membrane receptor belonging to the group of pattern recognition receptors) binds to lipids and the lipopolysaccharides, probably promoting the incorporation of LPS in chylomicrons. Transferred from the chylomicrons to other lipoproteins, the LPS finally arrives in the adipose tissue (Hersoug et al., 2016). The mobilization of energy by lipolysis e.g., during the beginning of lactation, for example, leads to a re-input of endotoxins into the bloodstream.
3. Damage of the gut barrier
In normal conditions, due to bacterial activity, endotoxins are present in the gut. Damage of the gut barrier allows translocation of these endotoxins (and bacteria) into the bloodstream.
4. Destruction of Gram-negative bacteria
Another “source” for endotoxins is the destruction of the bacteria. This can be done on the one hand by the organism’s immune system or by treatment with bactericidal substances targeting gram- bacteria (Kastner, 2002). To prevent an increased release of endotoxins, in the case of Gram-negative bacteria, a treatment with bacteriostatic substances only inhibiting the growth and not destroying the bacteria, or with bactericidal in combination with LPS-binding agents, would be a better alternative (Brandenburg, 2014).
5. Proliferation of gram-negative bacteria
As gram-negative bacteria also release small amounts of endotoxins when they grow, everything promoting their proliferation also leads to an increase of endotoxins:
Imbalanced feeding
High yielder cows e.g., are fed diets rich in starch, fat, and protein. Increased feeding of fat leads to a higher concentration of endotoxins in the organism, as the same “transporter” (scavenger receptor class B type 1, SR-BI) can be used (Hersoug et al., 2016) for the absorption of fat as well as for the absorption of endotoxins.
In a study with humans as representors of the monogastric species, Deopurkar and co-workers gave three different drinks (glucose – 100% carbohydrate, orange juice – 92% carbohydrate, and cream – 100% fat) to healthy participants. Only the cream drink increased the level of lipopolysaccharides in the plasma.
Infectious diseases
Infectious diseases like mastitis, metritis, and other infections caused by gram-bacteria such as E. coli, Salmonella, etc. can be regarded as sources of endotoxin release.
Decreased detoxification or degradation
Main responsible organ: the liver
Task: detoxification and degradation of translocated endotoxin. The liver produces substances such as lipopolysaccharide binding proteins (LBP) which are necessary for binding and neutralizing lipopolysaccharide structures.
During the post-partum period, the organism is in a catabolic phase, and lipolysis is remarkably increased for energy generation due to milk production. Increased lipolysis leads, as mentioned before, to a release of endotoxins out of the adipose tissue but also fatty degeneration of the liver. A fatty degenerated liver cannot bring the same performance in endotoxin clearance than a normal liver (Andersen, 2003; Andersen et al., 1996; Harte et al., 2010; Wilken, 2003). In a study conducted by Andersen and co-workers (1996), they couldn’t achieve complete clearance of endotoxins in cows with fatty livers. The occurrence of hepatic lipidoses increases after parturition (Reid and Roberts, 1993; Wilken, 2003).
Also, other diseases of the liver influence endotoxin clearance in the liver. Hanslin and co-workers (2019) found an impaired endotoxin elimination in pigs with pre-existing systemic inflammatory response syndrome.
Relation between lipid metabolism and endotoxin metabolism (according to Fürll, 2000, cited in Wilken, 2003)
Issues caused by endotoxins
Endotoxins, on the one hand, can positively stimulate the immune system when occurring in small amounts (Sampath, 2018). According to McAleer and Vella (2008), lipopolysaccharides are used as natural adjuvants to strengthen immune reaction in case of vaccination by influencing CD4 T cell responses. On the other hand, they are involved in the development of severe issues like MMA-Complex (Pig Progress) or a septic shock (Sampath, 2018).
MMA Complex in sows
MMA in sows is a multi-factorial disease appearing shortly after farrowing (12 hours to three days), which is caused by different factors (pathogens such as E. coli, Klebsiella spps., Staph. spps. and Mycoplasma spps., but also stress, diet). MMA is also known as puerperal syndrome, puerperal septicemia, milk fever, or toxemia. The last name suggests that one of the factors intervening in the disease is bacterial endotoxins. During the perinatal phase, massive catabolism of fat takes place to support lactation. The sows often suffer from obstipation leading to higher permeability of the intestinal wall, with bacteria, respectively endotoxins being transferred into the bloodstream. Another “source” of endotoxins can be the udder, as the prevalence of gram-negative bacteria in the mammary glands is remarkable (Morkoc et al., 1983).
The endotoxins can lead to an endocrine dysfunction: ↑ Cortisol, ↓ PGF2α, ↓Prolactin, ↓ Oxytocin. MMA stands for:
– Mastitis, a bacterial infection of the udder.
Mastitis can be provoked from two sides: on the one hand, endotoxemia leads to an elevation of cytokines (IL1, 6, TNFα). Lower Ca- and K-levels cause teat sphincter to be less functional, facilitating the entry of environmental pathogens into the udder and resulting in mastitis. On the other hand, due to farrowing stress, Cortisol concentrations get higher. The resulting immunosuppression enables E. coli to proliferate in the udder.
– Metritis, an infection of the uterus with vulvar discharges:
It leads to reduced contractions and, therefore, to prolonged and/or complicated farrowing or dead piglets. Metritis can be promoted by stress causing a decrease in oxytocin and prostaglandin F2α secretion. Morkoc and co-workers (1983) didn’t find a relation between metritis and endotoxins.
– Agalactia, a reduction or total loss of milk production:
In many cases, agalactia is not detected until the nursing litter shows signs of hunger and/or weight loss. At worst, the mortality rate in piglets increases. Probably, milk deficiency is caused by lower levels of the hormones involved in lactation. Prolactin levels e.g., may be dramatically reduced by small volumes of endotoxin (Smith and Wagner, 1984). The levels of oxytocin are often half those in normal sows (Pig Progress, 2020).
Endotoxin shock
A septic shock can be the response to the release of a high amount of endotoxins.
In the case of an infection with gram-negative bacteria, the animals are treated with (often bactericidal) antibiotics. Also, the immune system is eliminating the bacteria. Due to bacterial death, endotoxins are massively released. When not bound, they activate the immune system including macrophages, monocytes, and endothelial cells. Consequently, high amounts of cellular mediators like TNFα, Interleukin 1 (IL-1), IL-6, and leukotrienes are released. High levels of pro-inflammatory cytokines activate the complement and coagulation cascade. In some animals, then the production of prostaglandins and leukotrienes is stimulated, implicating high fever, decreased blood pressure, generation of thrombi in the blood, collapse, damaging several organs, and lethal (endotoxic) shock.
Endotoxic shock only occurs to a few susceptible animals, although the whole herd may have been immune-stimulated. A more severe problem is the decrease in the normally strong piglets’ performance, deviating resources from production to the immune system because of the endotoxemia.
Amplified diarrhea
Lipopolysaccharides lead to an augmented release of prostaglandins, which influence gastrointestinal functions. Together with leukotrienes and pro-inflammatory mediators within the mucosa, they reduce intestinal absorption (Munck et al., 1988; Chiossone et al., 1990) but also initiate a pro-secretory state in the intestine. Liang and co-workers (2005) observed a dose-dependent accumulation of abundant fluid in the small intestine resulting in increased diarrheagenic activity and a decreased gastrointestinal motility in rats.
Conclusion
Acting against Gram- bacteria can result in an even more severe issue – endotoxemia. Endotoxins, besides having a direct negative impact on the organism, also contribute to some diseases. Supporting gut health by different approaches, including the binding of toxins, helps to keep animals healthy.
By Inge Heinzl, EW Nutrition
References
Andersen, P.H. “Bovine endotoxicosis – some aspects of relevance to production diseases. A review.” Acta vet. scand. Suppl. 98 (2003): 141-155. DOI: 10.1186/1751-0147-44-S1-P57
Andersen, P.H., N. Jarløv, M. Hesselholt, and L. Bæk. “Studies on in vivo Endotoxin Plasma Disappearance Times in Cattle.” Zentralblatt für Veterinärmedizin. Reihe A 43 no. 2(1996): 93-101. DOI: 10.1111/j.1439-0442.1996.tb00432.x
Baker, B., S.L. Gaffin, M. Wells, B.C. Wessels and J.G. Brock-Utne. “Endotoxaemia in racehorses following exertion.” Journal of the South African Veterinary Association June (1988): 63-66. https://journals.co.za/docserver/fulltext/savet/59/2/1341.pdf?expires=1598542211&id=id&accname=guest&checksum=E50C766D318776E09CA41DA912F14CAD
Beutler, B. and T. Rietschel. “Innate immune sensing and its roots: The story of endotoxin.” Nature Reviews / Immunology 3(2003): 169-176. DOI: 10.1038/nri1004
Braun-Fahrländer, C., J. Riedler, U. Herz, W. Eder, M. Waser, L. Grize, S. Maisch, D. Carr, F. Gerlach, A. Bufe, R.P. Lauener, R. Schierl, H. Renz, D. Nowak and E. von Mutius. „Environmental exposure to endotoxin and its relation to asthma in school-age children. ”The New England Journal of Medicine 347 (2002): 869-877. DOI: 10.1056/NEJMoa020057.
Chiossone, D. C., P.L. Simon, P.L. Smith. “Interleukin-1: effects on rabbit ileal mucosal ion transport in vitro.” European Journal of Pharmacology 180 no. 2-3 (1990): 217–228. DOI: 10.1016/0014-2999(90)90305-P.
Deopurkar R., H. Ghanim, J. Friedman, et al. “Differential effects of cream, glucose, and orange juice on inflammation, endotoxin, and the expression of Toll-like receptor-4 and suppressor of cytokine signaling-3.” Diabetes care 33 no. 5 (2010):991–997.
Erridge, C., E. Bennett-Guerrero, and I.R. Poxton. “Structure and function of lipopolysaccharides.” Microbes and Infection 4 no. 8 (2002): 837-851. DOI: 10.1016/s1286-4579(02)01604-0
Fritsche, D. “Endotoxinpromovierte bakterielle Translokationen und Besiedelung von Uterus und Euter beim Hochleistungsrind im peripartalen Zeitraum.“ Dissertation. Leipzig, Univ., Veterinärmed. Fak. (1998)
Hanslin, K., J. Sjölin, P. Skorup, F. Wilske, R. Frithiof, A. Larsson, M. Castegren, E. Tano, and M. Lipcsey. “The impact of the systemic inflammatory response on hepatic bacterial elimination in experimental abdominal sepsis.” Intensive Care Medicine Experimental 7 (2019): art. 52. https://doi.org/10.1186/s40635-019-0266-x
Harte, A.L., N.F. da Silva, S.J. Creely, K.C. McGee, T. Billyard, E.M. Youssef-Elabd, G. Tripathi, E. Ashour, M.S. Abdalla, H.M. Sharada, A.I. Amin, A.D. Burt, S. Kumar, C.P. Day and P.G. McTernan. “Research Elevated endotoxin levels in non-alcoholic fatty liver disease.” Journal of Inflammation 7 (2010): 15-24. DOI: 10.1186/1476-9255-7-15
Hersoug, L.-G., P. Møller, and S. Loft. “Gut microbiota-derived lipopolysaccharide uptake and trafficking to adipose tissue: implications for inflammation and obesity.” Obesity Reviews 17 (2016): 297–312. DOI: 10.1111/obr.12370
Hurley, J. C. “Endotoxemia: Methods of detection and clinical correlates.” Clin. Microbiol. Rev. 8 (1995): 268–292. DOI: 10.1128/CMR.8.2.268
Kastner, A. “Untersuchungen zum Fettstoffwechsel und Endotoxin-Metabolismus bei Milchkühen vor dem Auftreten der Dislocatio abomasi.“ Inaug. Diss. Universität Leipzig, Veterinärmed. Fak. (2002). https://d-nb.info/967451647/34
Krüger M. “Escherichia coli: Problemkeim in der Nutztierhaltung.“ Darmflora in Symbiose und Pathogenität. Ökologische, physiologische und therapeutische Aspekte von Escherichia coli. 3. Interdisziplinäres Symposium. Alfred-Nissle-Gesellschaft (Ed.). Ansbach, 28.-29. Nov. (1997): 109-115.
Liang, Y.-C., H.-J. Liu, S.-H. Chen, C.-C. Chen, L.-S. Chou, and L. H. Tsai. “ Effect of lipopolysaccharide on diarrhea and gastrointestinal transit in mice: Roles of nitric oxide and prostaglandin E2.” World J Gastroenterol. 11 no. 3 (2005): 357–361. DOI: 10.3748/wjg.v11.i3.357
McAleer, J.P. and Vella, A.T. “Understanding how lipopolysaccharide impacts CD4 T cell immunity.” Crit. Rev. Immunol. 28 no. 4 (2008): 281-299. DOI:10.1615/CRITREVIMMUNOL.V28.I4.20
Morkok, A., L. Backstrom, L. Lund, A.R.Smith. “Bacterial endotoxin in blood of dysgalactic sows in relation to microbial status of uterus, milk, and intestine.” JAVMA 183 (1983): 786-789. PMID: 6629987
Munck, L.K., A. Mertz-Nielsen, H. Westh, K. Buxhave, E. Beubler, J. Rask-Madsen. “Prostaglandin E2 is a mediator of 5-hydroxytryptamine induced water and electrolyte secretion in the human jejunum.” Gut 29 no. 10 (1988): 1337-1341
Sampath, V.P. “Bacterial endotoxin-lipopolysaccharide; structure, function and its role in immunity in vertebrates and invertebrates.” Agriculture and Natural Resources 52 no. 2 (2018): 115-120. https://doi.org/10.1016/j.anres.2018.08.002
Seidler, T. “Freies Endotoxin in der Blutzirkulation von Schlachtschweinen: eine Ursache für bakterielle Translokationen?“ Diss. Universität Leipzig, Veterinärmed. Fak. (1998).
Smith, B.B. and W.C. Wagner. “Suppression of prolactin in pigs by Escherichia coli endotoxin.“ Science 224 no. 4649 (1984): 605-607
Wilken, H. “Endotoxin-Status und antioxidative Kapazität sowie ausgewählte Stoffwechselparameter bei gesunden Milch- und Mutterkühen.“ Inaugural Diss. Universität Leipzig (2003).



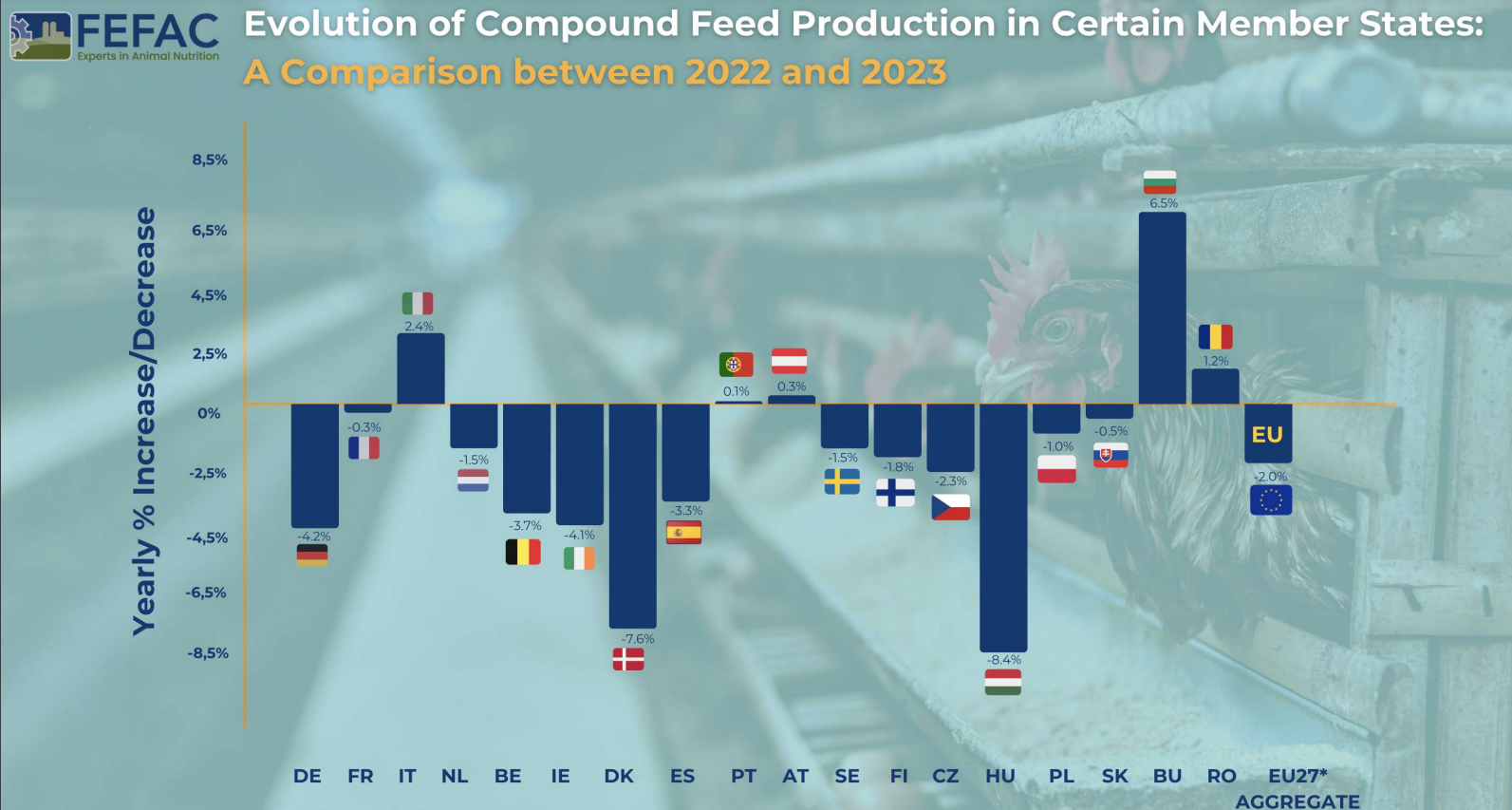






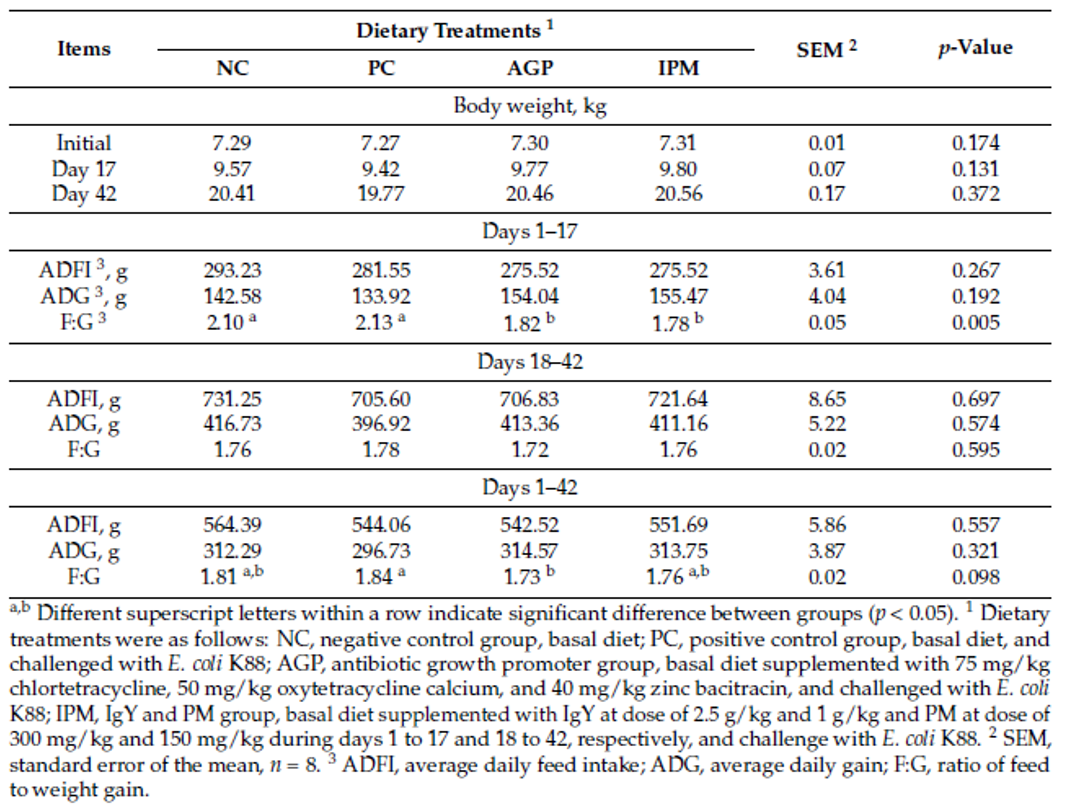
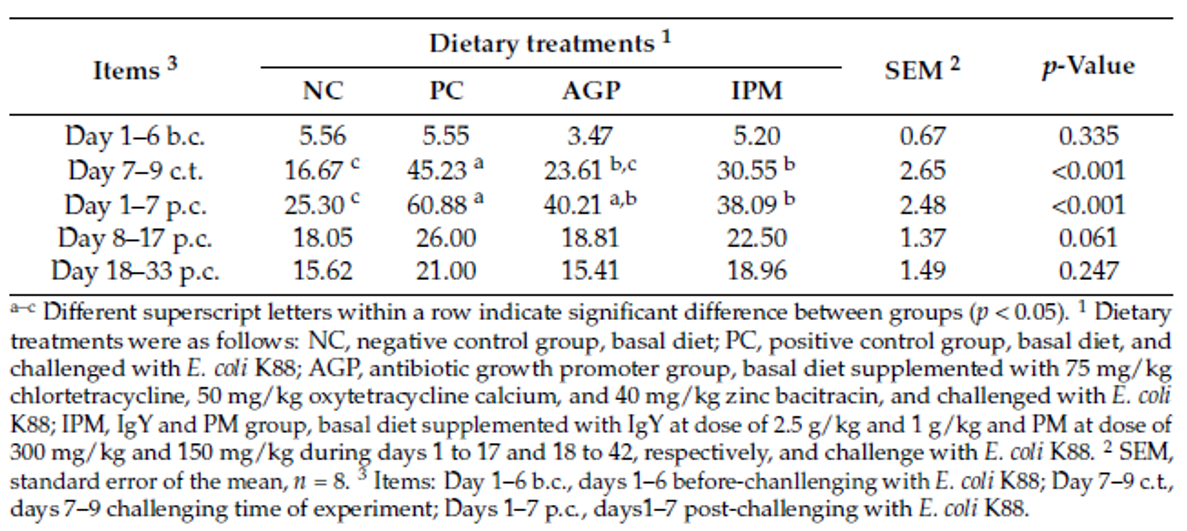
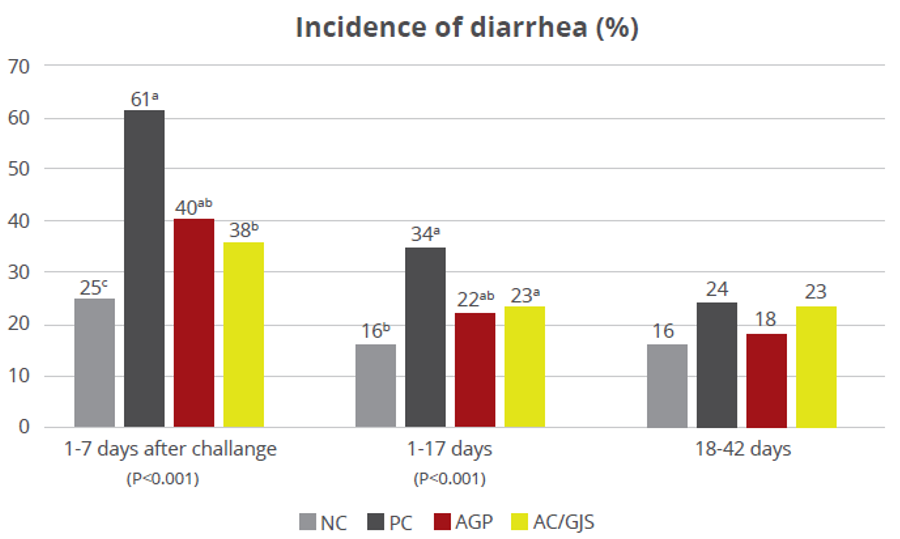
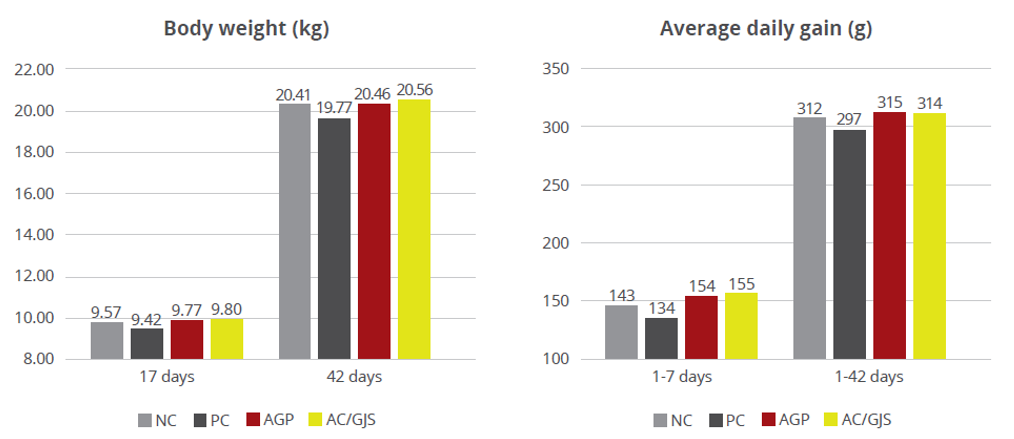
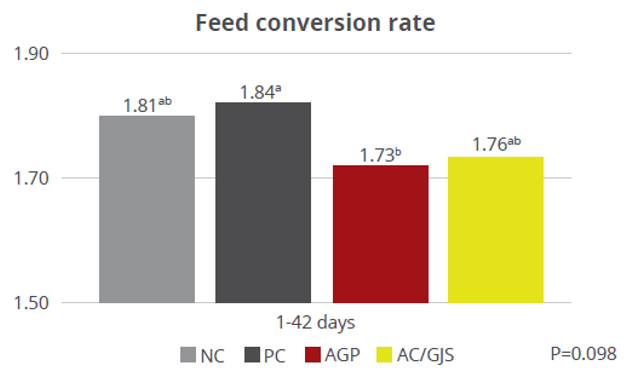
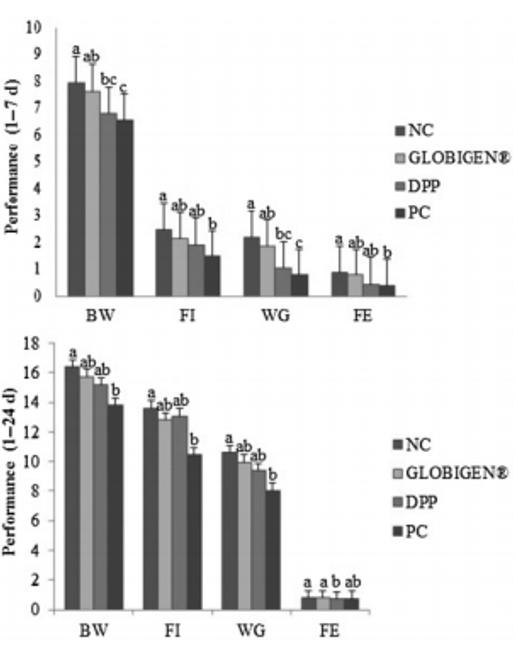















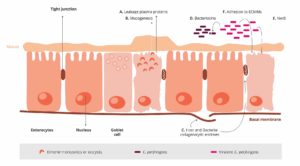
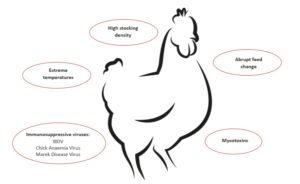
 To prevent a “feeding” of Clostridium perfringens, high content of water-soluble but indigestible NSPs such as wheat, wheat by-products, and barley should be avoided or at least minimized. Additionally, xylanases should be included in the feed formulation to reduce the deleterious effects of NSPs and improve feed energy utilization. Instead of these cereals, maize could be included in the diet. It is considered a perfect ingredient in broiler diets due to its high energy content and high nutrient availability.
To prevent a “feeding” of Clostridium perfringens, high content of water-soluble but indigestible NSPs such as wheat, wheat by-products, and barley should be avoided or at least minimized. Additionally, xylanases should be included in the feed formulation to reduce the deleterious effects of NSPs and improve feed energy utilization. Instead of these cereals, maize could be included in the diet. It is considered a perfect ingredient in broiler diets due to its high energy content and high nutrient availability.