Methane must be reduced – What about rumen performance?
Authors: Valentina Mayorga, Predrag Persak, and Inge Heinzl, EW Nutrition
Every day, dairy cows convert large amounts of feed into milk, but part of that valuable energy is inevitably lost in the form of methane produced during rumen fermentation. This gas not only represents a metabolic inefficiency for the animal but has also become one of the most discussed environmental impacts. Some organizations, such as the Institute for European Environmental Policy (IEEP), state that livestock production in the European Union accounts for approximately 65% of agricultural greenhouse gas (GHG) emissions (Hart et al., 2025). A very high number! As sustainability requirements and pressure from policymakers, processors, and consumers intensify, the dairy industry faces a critical challenge: reducing methane emissions while maintaining rumen health, fermentation efficiency, and productive performance.
Can feed additives master this difficult task?
In response to this challenge, a variety of feed additives and nutritional strategies have been developed to mitigate methane emissions in ruminants. However, methane mitigation must be approached carefully. Some products aim to suppress specific microbial pathways involved in methane formation, potentially altering rumen fermentation dynamics if not properly balanced.
One of the key mechanisms involved in methane mitigation is the redirection of hydrogen within the rumen. During ruminal fermentation, hydrogen produced by microbial activity can follow different metabolic pathways:
1. Traditionally, a significant portion of this hydrogen is utilized by methanogenic archaea to produce methane
2. However, hydrogen can also be incorporated into alternative pathways, particularly the formation of propionate. When rumen fermentation shifts toward propionate production, less hydrogen becomes available for methanogenesis, resulting in lower methane emissions. This process, often referred to as hydrogen redirection, enables methane reduction without suppressing overall microbial fermentation.
Among the nutritional approaches explored, plant-derived compounds, such as essential oils, have gained increasing attention for their ability to modulate rumen microbial populations. With essential oils, it is possible to influence specific groups of microorganisms involved in rumen fermentation, but also in methane production.
Many methanogens, e.g., are closely associated with rumen protozoa; therefore, reducing protozoal populations may indirectly decrease methane formation while maintaining normal fermentation processes.
Activo Premium trial gives reason for hope
Activo Premium, a blend of carefully selected essential oils, has been evaluated for its effects on rumen fermentation and methane production under controlled experimental conditions.
Trial Design:
Ingredients
g/kg DM
Chopped Tifton hay
500
Ground maize
325
Soybean meal
172
Chemical composition
% in DM
Organic matter
91.8
Crude protein
13.2
Neutral detergent fiber
59.4
The study was conducted at the CENA (University of São Paulo). Nine rumen-cannulated Santa Inês sheep (55 ± 3.7 kg of BW) were divided into three groups and randomly distributed in a 3×3 Latin square design for three consecutive periods of 37 days each.
At the beginning of each trial period, all sheep were fed ad libitum a basal diet without additives for 15 days. After this period, the animals were distributed to three different groups:
Group 1: Control (basal diet without additives) Group 2: Basal diet with 200 mg product/kg DM Group 3: Basal diet with 400 mg product/kg DM.
The sheep were fed experimental diets twice daily in equal portions and had free access to fresh water.
Results:
Experimental results showed a significant reduction in protozoa from day 7 after the first application and in methane production.
Figure 1: Decreasing levels of protozoa with increasing dosage of Activo Premium
Figure 2: Decreasing methane production due to the application of Activo Premium
Furthermore, propionate levels increased. The shift in SCFA towards propionic acid indicates that hydrogen, which methanogenic bacteria would have otherwise used for methane production, can now be used by rumen bacteria to produce bacterial protein, which then can serve as a nutrient for the sheep.
Figure 3: Shift of SCFA towards propionate with increasing dosage of Activo Premium
Phytomolecules are an optimal tool for methane reduction
Reducing greenhouse gas emissions has become a global responsibility to protect the future of our planet. Among agricultural sources, methane production from ruminants is considered one of the major contributors to greenhouse gas emissions. Therefore, effective nutritional strategies are increasingly important for sustainable livestock production. Phytomolecules-based products, such as Activo Premium, represent a promising approach to reducing methane formation by modulating rumen fermentation while maintaining animal productivity. This offers benefits for both farmers and the environment.
Rapetti, L., & Colombini, S. (n.d.). Evaluation of the effects of a blend of essential oils (named ACTIVO PREMIUM) on in vivo rumen microbiota and in vitro fermentation profile: Final report of the experimental trial. Università degli Studi di Milano, Department of Agricultural and Environmental Sciences.
Learning from AGP mechanisms to advance poultry nutrition
By Ilinca Anghelescu, Dr. Andreas Michels, Predrag Persak
Our understanding of how nutrition influences growth and resilience in poultry has greatly expanded in recent years. It is now clear that animal performance stems to a large extent from a balance between metabolism, immune function, and the gut microbiome. These systems interact continuously, and even small nutritional or environmental changes can shift the animals’ physiological response. This growing knowledge has encouraged the development of nutritional strategies and feed components that work through adaptive, non-antibiotic mechanisms. One recent proposed explanation for these responses has rapidly gained ground: hormetic modeling.
Hormetic modeling describes how small or moderate doses of nutritional components can activate beneficial adaptive responses (improved resilience or metabolic efficiency), while excessive doses become harmful. This idea parallels, largely speaking, Paracelsus’s famous principle: “The dose makes the poison.” In poultry nutrition, such hormetic patterns are well recognized in nutrients like trace elements (selenium, zinc) and specific amino acids (for example, arginine). At optimal levels, these nutrients support antioxidant defense, growth, and immune balance, whereas excessive intake may cause oxidative or metabolic stress
This review examines the hormetic principle and its application to modern poultry/swine feeding concepts, exploring how balanced nutrient design and controlled inclusion of bioactive compounds can strengthen cellular adaptation, improve stress tolerance, and enhance production efficiency.
How do AGPs actually work?
Despite AGP’s widespread historical use, the precise mechanisms by which subtherapeutic doses of antibiotics enhance animal productivity remained poorly understood. Recent advances in systems biology and mitochondrial research propose new answers, much needed to develop future advanced nutritional systems.
The traditional explanations for AGP efficacy have focused primarily on antimicrobial effects:
reducing nutrient competition from microorganisms
decreasing harmful bacterial metabolites
improving gut wall morphology (thinner gut wall ➡ better nutrient absorption)
preventing subclinical infections
However, these mechanisms alone could not fully explain why different classes of antibiotics with diverse mechanisms of action produce similar growth-promoting effects (Gutierrez-Chavez et al., 2025).
Niewold (2007) hypothesized that the primary mechanism of AGPs is non-antibiotic anti-inflammatory activity, reducing the energetic costs of chronic low-grade inflammation. Inflammation diverts nutrients from growth toward immune responses, with cytokine production (particularly IL-1β, IL-6, and TNF-α) suppressing anabolic pathways (Kogut et al., 2018). AGPs appear to selectively inhibit pro-inflammatory cytokine production without completely suppressing immune function.
A paper published in 2024 by Fernandez Miyakawa et al. proposes that antibiotics at subtherapeutic levels act primarily through mitochondrial hormesis and adaptive stress responses, and not simply through antimicrobial activity. In this model, mitochondria act as bioenergetic hubs and signaling centers. Low-dose antibiotics trigger mild mitochondrial stress, which triggers the activation of adaptive protective pathways.This in turn induces mitokine release, leading to systemic adaptive responses improving growth, feed efficiency, and disease tolerance.
Mechanism of action in the hormetic model of AGP efficiency
Hormesis is a biphasic mechanism whereby high doses are toxic, but low doses stimulate adaptive responses and are beneficial. In the case of AGPs, Fernandez Miyakawa et al. propose that low doses stimulate growth, stress resistance, and cellular repair.
Key signaling pathways
As Bottje et al. (2006, 2009) shows, efficient animals often have mitochondrial inner membranes that are less permeable to protons and other ions, allowing for more effective coupling between electron transport and ATP synthesis, which reduces energy loss through proton leak and maximizes the production of ATP per oxygen molecule consumed. Lower membrane permeability is influenced by factors like decreased membrane surface area per protein mass, specific membrane protein content (such as adenine nucleotide translocase), and fatty acid composition in the membrane phospholipids, all contributing to a tighter barrier that prevents unregulated electron or proton flow and supports higher energetic efficiency. Such features make mitochondria in efficient species more capable of maintaining membrane integrity and ATP generation, especially when facing environmental stress, as seen in freeze-tolerant animals whose mitochondria do not undergo damaging permeability transitions under extreme conditions.
Nrf2
Many AGPs interfere with mitochondrial protein synthesis and electron transport chain. At subtherapeutic levels, they cause a mild ROS increase, which triggers the activation of redox-sensitive transcription factor Nrf2. Since Nrf2 regulates over 250 antioxidant, detoxification, and anti-inflammatory genes, the result is improved cell survival, redox balance, and tolerance to stress.
Figure 1 From Zhang et al., 2024
Mitokine production
Mitokines are “signaling molecules that enable communication of local mitochondrial stress to other mitochondria in distant cells and tissues” (Burtscher 2023). Through fibroblast growth factor 21 (FGF21), growth differentiation factor 15 (GDF15), adrenomedullin2 (ADM2) etc, these stress signals are released systemically and coordinate tissue-wide responses, leading to improved growth and resilience.
Inflammation and disease defense
While the negative side of antibiotic growth promoters is well researched and understood (Rahman et al., 2022), science can advance by isolating the positive effects and attempting to offer different pathways to the same benefits. One such lesson can be derived from understanding inflammation pathways and responses.
Chronic low-grade intestinal inflammation is common in modern poultry production, due to diet, microbiota shifts, high metabolic demands etc. This inflammation diverts energy from growth to immune responses.
AGPs reduce the energy costs of this inflammation in three main ways:
Reduces inflammation through adaptive stress response
Raising the threshold to trigger inflammation
Promoting overall resilience, rather than simply killing pathogens
Fernandez Miyakawa et al. suggest, in this emerging model, that disease defense can operate two different actions: resistance to health challenges through reduction of the pathogen load (which is driven by the immune system and is energy costly); and overall resilience by reducing host damage without reducing the pathogen load. AGPs, the authors claim, mainly promote resilience by enhancing mitochondrial stress responses and tissue repair, i.e. more precisely:
Direct mitochondrial stimulation in intestinal epithelial cells
Metabolic optimization supporting growth and feed efficiency
Figure 2 From Fernandez Miyakawa et al., 2024.
In this context, “metabolic optimization” refers to the enhancement of metabolic processes within livestock or poultry to support efficient growth, feed conversion, and physiological resilience, without relying on immune-mediated pathways that are energetically costly. Scientific evidence shows that metabolic optimization involves improving nutrient assimilation, promoting more efficient energy production in tissues (such as mitochondrial ATP synthesis), and minimizing wasteful metabolic byproducts, resulting in reduced feed intake per unit of growth and better utilization of dietary nutrients (Rauw 2025, El-Hack 2025).
Function of feed additives and feed components
Feed additives and feed components in many ways represent the complete other side of the spectrum from antibiotics, but are there some features where antibiotics and feed additives come close in their functions? There is a good case to be made for certain feed additives ultimately working in the animal to achieve similar benefits to the desirable, non-medicinal usage of AGP´s. Especially with the emergent model of AGP mechanism described above, it is worth discussing how certain feed additives can support the same end goal: promoting animal resilience.
Lillejhoj et al (2018), Gutierrez-Chavez et al. (2025) and others outline the end-results such products must achieve:
Growth performance & feed conversion efficiency
Promotion of animal productivity under real-world conditions
Support gut homeostasis
Non-adverse effect on the immune system
Reduction of oxidative stress
Support organism in mitigation of enteric inflammatory consequences
Within the hormetic model, possibly the most important systemic benefit is, in one phrase, promoting resilience. Phytomolecules have long been used, in human and animal medicine, for the same end goal. The mechanisms described below should naturally be seen with caution, as phytomolecule microbiome effects can be subtler and context-dependent. However, the substantiating literature has been increasingly accumulating on these specific topics.
1. Immunometabolic regulation
Phytomolecules demonstrate remarkably similar anti-inflammatory effects to what Niewold (2007) suggested was a primary mechanism of AGPs: non-antibiotic anti-inflammatory activity, reducing the energetic costs of chronic low-grade inflammation. Inflammation diverts nutrients from growth toward immune responses, with cytokine production (particularly IL-1β, IL-6, and TNF-α) suppressing anabolic pathways (Kogut et al., 2018). AGPs appear to selectively inhibit pro-inflammatory cytokine production without completely suppressing immune function. A similar effect can be observed with various types of phytomolecules, which significantly reduced pro-inflammatory and/or increased anti-inflammatory cytokine expression in animals challenged with several pathogens. The anti-inflammatory mechanism appears to involve inhibition of NF-κB activation and modulation of MAPK signaling pathways (Kim et al., 2010; Long et al., 2021).
2. Mitochondrial hormesis and energy metabolism
Fernández Miyakawa et al. (2024, see above) proposed that AGPs exert growth-promoting effects through mitochondrial hormesis – subtherapeutic antibiotic doses induce mild mitochondrial stress, triggering adaptive responses that enhance mitochondrial function, energy metabolism, and cellular resilience. This mechanism, while requiring further validation, explains why different antibiotics with diverse targets produce similar growth outcomes.
The mitochondrial stress response involves activation of the IL-6 receptor family signaling cascade, which regulates metabolism, growth, regeneration, and homeostasis in liver and other tissues (Perry et al., 2024). Subtherapeutic antibiotic exposure activates proteins involved in growth and proliferation through IL-6R gp130 subunit signaling, including JAK, STAT, mTOR, and MAPK pathways.
Phytomolecules demonstrate similar mitochondrial effects. Perry et al. (2024) showed that increased activity of AMPK, mTOR, PGC-1α, PTEN, HIF, and S6K can also be available via phytomolecule activity, suggesting enhanced anabolic metabolism.
Capsicum oleoresin supplementation in broilers increased jejunal lipase and trypsin activity, enhanced ileal amylase activity, improved jejunal morphology, and modulated immune organ development, indicating enhanced digestive efficiency and nutrient utilization (Li et al., 2022).
Compounds such as vanillin, thymol, eugenol have been shown to improve glucose and lipid metabolism through TRPV1 activation and mitochondrial function enhancement (Gupta et al., 2022; Zhang et al., 2017).
3. Gut microbiota modulation
AGPs selectively reduce specific microbial populations, particularly Lactobacillus species that produce bile salt hydrolase (BSH). Since BSH reduces fat digestibility and thus weight gain, AGP-mediated reduction of BSH-producing bacteria enhances energy extraction and growth (Lin, 2014; Bourgin et al., 2021).
Recent research by Zhan et al. (2025) using single-molecule real-time 16S rRNA sequencing demonstrated that therapeutic antibiotic doses (lincomycin, gentamicin, florfenicol, benzylpenicillin, ceftiofur, enrofloxacin) significantly altered chicken gut microbiota composition, with Pseudomonadota and Bacillota becoming dominant phyla after exposure. Different antibiotics produced distinct temporal effects on microbial diversity and community structure.
Phytomolecules exert targeted antimicrobial effects while promoting beneficial bacteria. Dietary supplementation with 800 mg/kg Capsicum extract in Japanese quails reduced cecal counts of pathogenic bacteria (Salmonella spp., E. coli, coliforms) while modulating Lactobacilli populations (Reda et al., 2020).
In pigs, 80 mg/kg natural capsicum extract increased cecal propionic acid and total volatile fatty acid concentrations, with increased butyric acid in the colon – indicating enhanced fermentation by beneficial bacteria (Long et al., 2021).
Capsicum and Curcuma oleoresins altered intestinal microbiota composition in commercial broilers challenged with necrotic enteritis, reducing disease severity through microbiome modulation (Kim et al., 2015).
Capsaicin demonstrates selective antimicrobial activity, inhibiting pathogenic Gram-negative bacteria while favoring development of certain Gram-positive bacteria. The antibacterial mechanism involves induction of osmotic stress and membrane structure damage (Adaszek et al., 2019; Rosca et al., 2020).
4. Intestinal barrier function and gut health
AGPs have been associated with improved intestinal morphology, including increased villus height and reduced crypt depth, which enhance absorptive capacity (Gaskins et al., 2002).
Phytomolecules produce similar or superior effects. Capsicum extract (80 mg/kg) in pigs increased ileal villus height and upregulated MUC-2 gene expression, indicating enhanced gut barrier function and integrity. The improved barrier function correlated with reduced diarrhea incidence (Liu et al., 2013; Long et al., 2021).
Allium hookeri extract increased expression of tight junction proteins (claudins, occludins, ZO-1) in LPS-challenged broiler chickens, demonstrating direct enhancement of barrier integrity (Lee et al., 2017).
5. Oxidative stress mitigation
Oxidative stress impairs growth by damaging cellular components and triggering inflammatory responses. AGPs reduce oxidative stress indirectly through anti-inflammatory effects and microbiota modulation (Bortoluzzi et al., 2021).
Phytomolecules possess direct antioxidant properties. Capsicum extract (50 mg/kg) in heat-stressed quails reduced serum and ovarian malondialdehyde (MDA) while increasing superoxide dismutase (SOD) and catalase (CAT) activities. Ovarian transcription factors showed decreased NF-κB and increased Nrf2 and HO-1 expression (Sahin et al., 2016).
A mixture of herbal extracts including pepper reduced thiobarbituric acid reactive substances and MDA in broiler liver and muscle, while increasing glutathione peroxidase (GSH-Px) activity and improving antioxidant enzyme expression (Saleh et al., 2018).
Capsicum extract (80 mg/kg) in pigs increased total antioxidant capacity, SOD, and CAT while reducing MDA levels, demonstrating robust antioxidant effects (Long et al., 2021).
Standardization and controlled release: Critical success factors
A major criticism of phytomolecules has been inconsistent efficacy across studies. However, this variability largely reflects differences in:
Active compound concentrations
Bioavailability and stability
Dosing precision
Product quality and standardization
Microencapsulation is one of the technologies that address the standardization and bioavailability challenges. It protects volatile compounds from degradation during feed processing and storage, with encapsulated essential oils showing significantly higher retention compared to unprotected forms (Stevanović et al., 2018). By creating a protective barrier around active ingredients, microencapsulation enables controlled release in specific regions of the gastrointestinal tract, improving absorption efficiency and reducing dose variability (Bringas-Lantigua et al., 2011). The technology also masks unpalatable flavors that can reduce feed intake while standardizing active ingredient concentrations through precise manufacturing processes (Gharsallaoui et al., 2007). Studies demonstrate that spray-dried microencapsulated essential oils achieve encapsulation efficiencies exceeding 93% with minimal loss during storage (Hu et al., 2020), and can be engineered for enzyme-mediated release to ensure bioactive delivery at optimal intestinal sites (Elolimy et al., 2025).
Mechanistic synthesis: An integrated model
The evidence indicates that both AGPs and phytomolecules operate through an integrated network of effects:
Primary Level: Selective antimicrobial effects modify gut microbiota composition
This integrative model explains why multiple antibiotics with different mechanisms produce similar growth outcomes: they converge on common pathways regulating immunometabolism and mitochondrial function (Fernández Miyakawa et al., 2024).
Phytomolecules operate through the same mechanistic framework but with potential advantages:
Safety and antimicrobial resistance considerations
Antibiotic exposure significantly disrupts gut microbiota diversity and stability, with effects persisting beyond withdrawal periods. The study by Zhan et al. (2025) demonstrated that different antibiotics produce varying degrees of microbiota disruption, with florfenicol and gentamicin showing the strongest and most persistent effects.
In contrast, phytomolecules generally do not generate resistance through the same mechanisms as antibiotics. Some phytochemicals may actually enhance antibiotic efficacy and resensitize resistant bacteria through structural modifications of bacterial membranes (Khameneh et al., 2021; Suganya et al., 2022).
However, one study reported increased correlation between antibiotic resistance genes (ARGs) and mobile genetic elements in pig feces after mushroom powder supplementation, suggesting that certain phytogenic compounds may increase ARG mobility (Muurinen et al., 2021). This emphasizes the need for continued surveillance of phytomolecule effects on resistance gene dynamics.
Capsaicinoids and capsinoids have well-established safety profiles. Capsiate, a non-pungent analogue of capsaicin, exhibits substantially lower toxicity while maintaining similar metabolic and growth-promoting effects (Gupta et al., 2022). No adverse effects on animal health or product quality have been reported at recommended dosages in reviewed studies.
Future directions and research needs
Despite substantial progress, several areas require further investigation:
Mechanistic refinement: Detailed characterization of signaling pathways, particularly the IL-6R/gp130 cascade and mitochondrial stress responses
Precision formulation: Development of combinations optimized for specific production stages, environmental conditions, and disease pressures
Bioavailability optimization: Advanced delivery systems ensuring consistent active compound release and absorption
Microbiome-host interaction mapping: High-resolution characterization of microbial community shifts and their functional consequences
Economic validation: Large-scale production trials assessing cost-effectiveness compared to AGPs and disease management costs
Conclusions
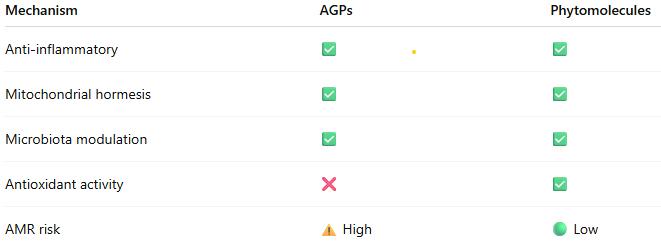
The scientific evidence demonstrates that standardized phytomolecules operate through well-characterized biological mechanisms that substantially replicate those of AGPs:
Anti-inflammatory effects reducing energetic costs of immune activation
Mitochondrial hormesis enhancing energy metabolism and cellular resilience
Selective microbiota modulation supporting beneficial bacteria while controlling pathogens
Intestinal barrier enhancement improving nutrient absorption and reducing translocation
Antioxidant activity mitigating oxidative stress and supporting immune function
When properly standardized and formulated for controlled release, phytomolecules deliver growth promotion, feed efficiency improvements, and disease resistance comparable to AGPs, while potentially offering advantages in AMR risk profile, stress resilience, and consumer acceptance.
The mechanistic convergence between AGPs and phytomolecules, coupled with demonstrated efficacy in controlled trials, provides producers with confidence that science-based phytomolecular interventions represent legitimate alternatives to AGPs. Success depends on product standardization, appropriate dosing, and understanding that phytomolecules work through fundamental biological pathways rather than undefined or mystical mechanisms.
As the livestock industry continues to navigate the post-AGP era, standardized phytomolecules offer a scientifically sound, mechanistically validated approach to maintaining animal performance, health, and welfare while addressing antimicrobial resistance concerns.
References
Adaszek, Ł., et al. “Properties of Capsaicin and Its Utility in Veterinary and Human Medicine.” Research in Veterinary Science, vol. 123, 2019, pp. 14 – 19.
Bottje, W., et al. “Mitochondrial proton leak kinetics and relationship with feed efficiency within a single genetic line of male broilers”. Poultry Science, Volume 88, Issue 8, 1 August 2009, p. 1683-1693.
Bortoluzzi, C., et al. “A Protected Complex of Biofactors and Antioxidants Improved Growth Performance and Modulated the Immunometabolic Phenotype of Broiler Chickens Undergoing Early Life Stress.” Poultry Science, vol. 100, 2021, p. 101176.
Bourgin, M., et al. “Bile Salt Hydrolases: At the Crossroads of Microbiota and Human Health.” Microorganisms, vol. 9, no. 1122, 2021.
Bravo, D., et al. “A Mixture of Carvacrol, Cinnamaldehyde, and Capsicum Oleoresin Improves Energy Utilization and Growth Performance of Broiler Chickens Fed Maize-Based Diet.” Journal of Animal Science, vol. 92, 2014, pp. 1531 – 1536.
Bringas-Lantigua, M., et al. “Influence of Spray-Dryer Air Temperatures on Encapsulated Mandarin Oil.” Drying Technology, vol. 29, 2011, pp. 520–526.
Burtscher, J., et al. “Mitochondrial Stress and Mitokines in Aging.” Aging Cell, vol. 22, no. 2, 2023, e13770.
El-Hack, M. et al. “Integrating metabolomics for precision nutrition in poultry: optimizing growth, feed efficiency, and health”. Frontiers in Veterinary Science, Sec. Animal Nutrition and Metabolism, Volume 12 – 2025. https://doi.org/10.3389/fvets.2025.1594749
Elolimy, Ahmed A., et al. “Effects of Microencapsulated Essential Oils and Seaweed Meal on Growth Performance, Digestive Enzymes, Intestinal Morphology, Liver Functions, and Plasma Biomarkers in Broiler Chickens.” Journal of Animal Science, vol. 103, 2025, p. skaf092, https://doi.org/10.1093/jas/skaf092.
Fernández Miyakawa, Mariano E., et al. “How Did Antibiotic Growth Promoters Increase Growth and Feed Efficiency in Poultry?” Poultry Science, vol. 103, no. 2, 2024, article 103136. https://doi.org/10.1016/j.psj.2023.103136
Gaskins, H. Rex, C. T. Collier, and D. B. Anderson. “Antibiotics as Growth Promotants: Mode of Action.” Animal Biotechnology, vol. 13, no. 1, 2002, pp. 29 – 42.
Gharsallaoui, A., et al. “Applications of Spray-Drying in Microencapsulation of Food Ingredients: An Overview.” Food Research International, vol. 40, no. 9, 2007, pp. 1107-21.
Gutiérrez-Chávez, Vanesa, et al. “Capsaicinoids and Capsinoids of Chilli Pepper as Feed Additives in Livestock Production: Current and Future Trends.” Animal Nutrition, vol. 22, 2025, pp. 483 – 501. https://doi.org/10.1016/j.aninu.2025.03.014.
Gupta, A., et al. “Capsaicin and Capsinoids: Recent Updates on Their Health Benefits and Mechanisms of Action.” Phytotherapy Research, vol. 36, no. 5, 2022, pp. 1898 – 1912.
Hu, Q., Li, X., Chen, F., Wan, R., Yu, C.-W., Li, J., McClements, D. J., & Deng, Z. (2020). “Microencapsulation of an essential oil (cinnamon oil) by spray drying: Effects of wall materials and storage conditions on microcapsule properties“. Journal of Food Processing and Preservation, 44(11). https://doi.org/10.1111/jfpp.14805
Khameneh, B., et al. “Mechanisms of Antibiotic Resistance Resensitization by Phytochemicals: Review.” Phytomedicine, vol. 85, 2021, p. 153529.
Kim, D. K., et al. “Effects of Capsicum and Curcuma on Necrotic Enteritis in Broilers.” Poultry Science, vol. 94, 2015, pp. 2314 – 2321.
Kim, J. S., et al. “Anti-inflammatory Effects of Plant-Derived Molecules via NF-κB and MAPK Pathways.” International Immunopharmacology, vol. 10, no. 3, 2010, pp. 306 – 314.
Lee, S. H., et al. “Allium Hookeri Extract Enhances Tight Junction Proteins in Broilers.” Journal of Animal Physiology and Animal Nutrition, vol. 101, no. 1, 2017, pp. e48 – e56.
Li, X., et al. “Capsicum Oleoresin Supplementation Improves Digestive Enzyme Activity and Gut Morphology in Broilers.” Poultry Science, vol. 101, no. 7, 2022, p. 101844.
Lin, J. “Effect of Antibiotics on the Intestinal Microbiota and Their Role in Animal Growth.” Animal Biotechnology, vol. 25, no. 3, 2014, pp. 149 – 157.
Lillehoj, H., et al. “Phytochemicals as Antibiotic Alternatives to Promote Growth and Enhance Host Health.” Veterinary Research, vol. 49, no. 76, 2018.
Liu, Y., et al. “Dietary Capsicum Extract Enhances Intestinal Barrier Function and Growth in Pigs.” Journal of Animal Science, vol. 91, 2013, pp. 518 – 525.
Long, L., et al. “Phytogenic Feed Additives Modulate Intestinal Immunity and Antioxidant Status in Pigs and Poultry.” Frontiers in Veterinary Science, vol. 8, 2021, p. 620998.
Muurinen, J., et al. “Mushroom Powder Supplementation Increases Antibiotic Resistance Gene Mobility in Pig Feces.” Frontiers in Microbiology, vol. 12, 2021, p. 676678.
Niewold, T. A. “The Non-antibiotic Anti-inflammatory Effect of Antimicrobial Growth Promoters, the Real Mode of Action? A Hypothesis.” Poultry Science, vol. 86, 2007, pp. 605 – 609.
Perry, F., C. N. Johnson, L. Lahaye, E. Santin, D. R. Korver, M. H. Kogut, and R. J. Arsenault. “Protected Biofactors and Antioxidants Reduce the Negative Consequences of Virus and Cold Challenge by Modulating Immunometabolism via Changes in the Interleukin-6 Receptor Signaling Cascade in the Liver.” Poultry Science, vol. 103, no. 9, 2024, article 104044. https://doi.org/10.1016/j.psj.2024.104044
Rahman, Md, et al. “Insights in the Development and Uses of Alternatives to Antibiotic Growth Promoters in Poultry and Swine Production.” Antibiotics, vol. 11, no. 6, 2022, p. 766, https://doi.org/10.3390/antibiotics11060766.
Reda, F. M., et al. “Capsicum Extract Supplementation Modulates Gut Microbiota and Performance in Japanese Quails.” Animal Feed Science and Technology, vol. 265, 2020, p. 114507.
Rosca, I., et al. “Capsaicin Induces Osmotic Stress in Gram-negative Pathogens.” Veterinary Sciences, vol. 7, no. 4, 2020, p. 172.
Sahin, K., et al. “Dietary Capsicum Extract Reduces Oxidative Stress in Heat-stressed Japanese Quails.” Poultry Science, vol. 95, no. 2, 2016, pp. 231 – 240.
Saleh, A. A., et al. “Herbal Extract Mixtures Improve Antioxidant Status and Performance in Broilers.” Poultry Science, vol. 97, no. 11, 2018, pp. 3927 – 3936.
Stevanović, Z. D., et al. „Essential oils as feed additives—Future perspectives”. Molecules, 23(7), 2018, pp1717.
Suganya, R., et al. “Phytochemicals in Combination with Antibiotics: Antimicrobial Resistance Breakers.” Antibiotics, vol. 11, 2022, p. 123.
Zhang, Benyuan et al. “Mitochondrial Stress and Mitokines: Therapeutic Perspectives for the Treatment of Metabolic Diseases.” Diabetes & Metabolism Journal vol. 48,1, 2024, pp. 1-18.
Zhan, Ru, et al. “Effects of Antibiotics on Chicken Gut Microbiota: Community Alterations and Pathogen Identification.” Frontiers in Microbiology, vol. 16, 2025, article 1562510. https://doi.org/10.3389/fmicb.2025.1562510
Zhang, Y., et al. “Effects of Vanillin, Thymol, and Eugenol on Glucose and Lipid Metabolism via TRPV1 Activation.” Journal of Agricultural and Food Chemistry, vol. 65, no. 13, 2017, pp. 2719 – 2727.
Energy Metabolism in Pigs: Disease and stress impact efficiency
By Dr. Inge Heinzl, Editor, and Predrag Persak, Regional Technical Manager North Europe
For profitable pig production, efficient energy metabolism is essential. Every kilojoule consumed must be wisely spent – on maintenance, growth, reproduction, or defense. An impacted energy metabolism due to disease or stress impacts animal performance and farm profitability.
Different faces of energy
Energy metabolism determines how efficiently pigs convert feed into body mass. The Gross energy (GE) of the diet, which the use of a calorimeter can determine, is progressively reduced by losses in feces (→digestible energy – DE), urine, gases (→metabolizable energy – ME), and heat, resulting in the →net energy (NE), which is then available for maintenance and performance (growth, milk…).
The requirements for maintenance include the minimum energy that an organism needs to maintain essential functions under standardized conditions and at complete rest. This includes respiration, thermoregulation, tissue turnover, and immune system activity. Only energy in excess of these needs is available for performance. The ratio between additional retained energy and additional energy intake defines the incremental efficiency of nutrient utilization. Under normal conditions, healthy, fast-growing pigs display high incremental efficiencies for both protein and energy deposition by channeling energy efficiently into lean tissue and approximately 25-30% of the metabolizable energy from the feed is used for maintenance, 20-25% for lean gain, and the rest for fat deposition, driving daily gain and carcass quality (Patience, 2019).
However, disease, immune stress, and suboptimal environmental conditions can disrupt this delicate balance, diverting nutrients from growth to survival processes (Obled, 2003). The activation of the immune system leads to reduced feed efficiency, slower growth, and inferior meat quality.
Disease generates costs
The health challenge of disease causes energy loss through several key mechanisms (Patience, 2019).
The activation of the immune system becomes an energetic priority. It consumes significant amounts of energy and nutrients, such as glucose and specific amino acids, to produce immune cells and acute-phase proteins, such as haptoglobin and CRP, and to combat pathogens. The nutrients are redirected away from performance toward immune defense, i.e., less energy available for growth performance or even a mobilization of body reserves (fat deposits). A study conducted by Huntley et al. (2017) showed a 23.6% higher requirement for metabolizable energy to activate and maintain the immune system, resulting in a 26% lower ADG.
Physiological responses to disease, such as fever (heat production), shivering, or increased physical activity due to discomfort or listlessness, require energy.
Additional lower feed intake due to reduced appetite, leading to less energy consumption and intensifying the problem of energy repartitioning.
Environmental challenges are energy-consuming
Besides environmental conditions that cause disease due to high pathogenic pressure, environmental challenges are often related to thermoregulation.
1. Cold stress
In the case of cold stress, the ambient temperature falls below the pig’s lower critical temperature. The animal must spend extra energy to produce heat and maintain a constant body temperature. Alternatively, it can achieve this through shivering (muscle friction generates heat) and the release of thyroid hormones, which increase the metabolic rate and boost body temperature. Another possibility is huddling with other pigs. If the pigs eat more to gain extra energy for warmth, they increase production costs.
2. Heat stress
Excessive temperature leads to heat stress, and the animals attempt to cope through several mechanisms. Increased respiratory evaporation by panting is energy-intensive. Other possibilities are lying spread out on cool surfaces (conduction), seeking shade, and reducing physical activity to minimize heat production. To reduce metabolic heat production, pigs decrease their feed intake; however, this results in an energy deficit and likely mobilizes body reserves, especially in lactating sows.
3. Poor housing and management
High ventilation rates, draughts, wet floors, high stocking densities, and, too often, mixing of pigs are other stressors that require adequate energy-consuming responses. Also, an environment that facilitates excessive heat loss, e.g., through cold concrete floors, constrains the pigs to expend more ME to compensate. Poor-quality air with high levels of harmful gases, such as ammonia or hydrogen sulfide, or dust can lead to respiratory issues and energy expenditure for immune defense.
What are the detailed consequences?
Energy required for immune defense cannot be used for the production of meat, milk, or eggs. Several energy-consuming processes are triggered during an immunological challenge.
Glucose, an important energy source
Several scientists (Spurlock, 1997; Rigobelo and Ávila, 2011) have stated that glucose is primarily used to meet the increased energy demands of an activated immune system. According to Kvidera et al. (2017), the reason might be that stimulated leucocytes change their metabolism from oxidative phosphorylation to aerobic glycolysis (Palsson-McDermott and O’Neill, 2013). A trial conducted by Kvidera et al. (2017) confirmed the high need for glucose. In their trial with E. coli LPS-challenged crossbred gilts, they measured the amount of glucose required to maintain normal blood glucose levels (euglycemia). They calculated that an acutely and intensely activated immune system requires 1.1 g of glucose/kg body weight0.75/h. As they obtained similar results in ruminants (Kvidera et al., 2016 and 2017), they regard this glucose requirement as conserved across species and physiological states. In a confirming study, McGilvray and coworkers (2018) observed a significant (P<0.01) decrease in blood glucose in pigs after injection of E. coli LPS.
A further energy-consuming process is the increase in body temperature (fever): To increase body temperature by 1°C, the metabolic rate must be raised by 10-12.5% (Evans et al., 2015).
Influence on protein metabolism
Stimulation of the immune system in growing pigs may lead to a redistribution of amino acids from protein retention to immune defense. Amino acids are needed as a ‘substrate’ to synthesize immune system metabolites, such as acute-phase proteins (e.g., haptoglobin, a-fibrinogen, antitrypsin, lipopolysaccharide-binding protein, C-reactive protein, and others (Rakhshandeh and De Lange, 2011)), immunoglobulins, and glutathione (Reeds and Jahoor, 2001). This impacts the requirements for amino acids quantitatively but also qualitatively, i.e., the amino acid profile. Various studies indicated an increased need for Methionine, cysteine, branched-chain amino acids (BCAAs), aromatic amino acids, Threonine, and Glutamine during immune system stimulation (Reeds et al., 1994; Melchior et al., 2004; Calder et al., 2006; Rakhshandeh and de Lange, 2011; Rakhshandeh et al., 2014).
If the required amino acids are not available, they must be either synthesized or obtained from body protein. This costs energy, leads to muscle mass degradation, and causes an imbalance in amino acid levels. Excess amino acids are catabolized, resulting in an increase in blood urea nitrogen (BUN). McGilvray et al. (2018), e.g., observed a 25% increase in BUN in their study, in which they stimulated pigs’ immune systems with LPS.
Another possibility is using amino acids as energy sources. L-Glutamine, for example, is a crucial energy source for immune cells and the primary energy substrate for mucosal cells (Mantwill, 2025).
Carcass and meat quality
As already mentioned, immune stimulation or disease leads to protein degradation. Plank and Hill (2000) reported a loss of up to 20% of body protein (mainly skeletal muscle) in critically ill humans over 3 weeks. This protein degradation influences carcass yield and quality by reducing the amount of muscle meat.
Another effect is a decrease in the muscle cross-sectional area of fibers and a significant shift from the myosin heavy chain (MHC)-II towards the MHC-I type (Gilvray et al, 2019)
How can feed additives support pigs in health challenges?
Health challenges can occur due to infections by bacteria, viruses, fungi, or protozoa, as well as due to myco-, exo-, or endotoxins. Phytomolecules-based and toxin-binding can help animals cope with these health challenges.
Phytomolecules have several health-supporting effects
Phytomolecules can support animals in the case of a health challenge by directly fighting bacteria – antimicrobial effect (Burt, 2004; Rowaiye et al., 2025), scavenging free radicals – antioxidant effect (Saravanan et al., 2025; Dhir, 2022), or mitigating infection – anti-inflammatory effect (Saravanan et al., 2025).
A trial with the phytomolecules-based product Ventar D demonstrated its antimicrobial and microbiome-modulating effects (Heinzl, 2022). The product clearly reduced the populations of Salmonella enterica, E. coli, and Clostridium perfringens but spared the beneficial lactobacilli.
The anti-inflammatory effects of phytomolecules inhibit the activity of pro-inflammatory cytokines and chemokines from endotoxin-stimulated immune cells and epithelial cells (Lang et al., 2004; Lee et al., 2005; Liu et al., 2020), and there is an indication that the anti-inflammatory effects might be mediated by blocking the NF-κB activation pathway (Lee et al., 2005). A trial confirmed this thesis by showing a dose-dependent reduction of NFκB activity in LPS-stimulated mouse cells (-11% & -54% with 50 & 200 ppm Ventar D, respectively) (Figure 1).
Figure 1: NFκB activity in LPS-stimulated mouse cells with different inclusion rates of Ventar D (light color: no LPS; dark color: 0.25 µg LPS/mL)
Additionally, Ventar D increases interleukin-10, a cytokine with anti-inflammatory properties, and decreases interleukin-6, a pro-inflammatory cytokine. The result is a dose-dependent decline in the ratio of IL-6 to IL-10 (Figure 2), indicating the effectiveness of the product.
Figure 2: IL-6/IL-10 ratio
The effects of Ventar D, which support the immune system and redirect energy to enhance growth performance, result in higher daily gains and improved feed conversion. This was observed in a trial conducted on a commercial farm in Germany, using, on average, 26-day-old weaned piglets with a mean body weight of approximately 8 kg. Just after weaning, young animals experience stress (new feed, new groups, and separation from the dam) and are more susceptible to disease.
Two groups of piglets were fed either the regular feed of the farm (Control) or the regular feed + 100 g Ventar per MT of feed. The results for final weight and FCR are shown in Figures 3 and 4
Figure 3: Final weight in weaned piglets with and without Ventar D
Figure 4: FCR in weaned piglets with and without Ventar D
Toxin-binding products support animals against health challenges caused by toxins
As mentioned, various toxins, including myco-, endo-, and exotoxins, can harm animals. The danger of mycotoxins lurks in many feeds, and exo- and endotoxins derive from bacteria. Toxin-binding products, possibly supplemented with phytomolecules that support health (e.g., liver protection), can help animals cope with these challenges.
Solis Max 2.0, a toxin solution containing bentonite and phytomolecules, showed excellent binding performance for myco- and endotoxins (Figures 5 and 6).
Trial with endotoxins
Two samples were prepared: one with only 25 EU (1 EU equivalent to approximately 100 pg or 10,000 cells) of LPS of E. coli O55:B5 LPS/mL solution, and one with the same concentration of LPS but also containing 700 mg Solis Max 2.0/mL.
Solis Max 2.0 bound about 80% of endotoxin.
Figure 5: Endotoxin-binding capacity of Solis Max
Trial with mycotoxins
In another in vitro trial, the binding capacity of Solis Max 2.0 for six different kinds of mycotoxins was evaluated. For that purpose, samples with 800 ppb AFB1, 400 ppb OTA, 800 ppb DON, 300 ppb T2, 2,000 ppb FB1, or 1,200 ppb ZEN were prepared, and Solis max was added at two inclusion rates, one corresponding to 1 kg/t, the other to 2 kg/t. The binding capacities ranged from 40.7% for OTA to 96% for AFB1, with the lower inclusion rate, and from 61.5% for OTA to 99% for AFB1, with the higher inclusion rate.
Figure 6: Mycotoxin-binding capacity of Solis Max
Health support by toxin-binding solutions improves performance
The mitigating effects of Solis Max concerning the negative impact of toxins are also reflected in performance. A trial involving 24 female weaned piglets was conducted to evaluate the mitigating effects of Solis Max in the event of a challenge with a naturally contaminated diet (3,400 ppb of DON and 700 ppb of ZEA). Solis Max was added to one half of the challenged piglets. The addition of Solis Max to the contaminated diet not only compensates for growth performance parameters, such as weight gain and feed conversion, but also for Vulva and tail necrosis scores. The results are shown in Figures 7-11.
Figure 7: Feed intake (g)
Figure 8: Body weight gain (g)
Figure 9: FCR
Figure 10: Vulva score
Figure 11: Tail necrosis score
Tools are available to prevent the unnecessary expenditure of energy for immune protection
As the various references in the article demonstrate, health challenges such as pathogens or toxins not only spoil the appetite of animals but also require energy due to the activation of the immune system. Products based on phytomolecules, as well as toxin solutions, can help animals cope with these challenges and conserve energy for improved performance.
References:
Balli, Swetha, Karlie R. Shumway, and Shweta Sharan. “Physiology, Fever.” StatPearls [Internet]., September 4, 2023. https://www.ncbi.nlm.nih.gov/books/NBK562334/.
Burt, Sara. “Essential Oils: Their Antibacterial Properties and Potential Applications in Foods—a Review.” International Journal of Food Microbiology 94, no. 3 (August 2004): 223–53. https://doi.org/10.1016/j.ijfoodmicro.2004.03.022.
Calder, Phillip C. “Branched-Chain Amino Acids and Immunity ,.” The Journal of Nutrition 136, no. 1 (January 2006). https://doi.org/10.1093/jn/136.1.288s.
Dhir, Vivek. “Emerging Prospective of Phytomolecules as Antioxidants against Chronic Diseases.” ECS Transactions 107, no. 1 (April 24, 2022): 9571–80. https://doi.org/10.1149/10701.9571ecst.
Evans, Sharon S., Elizabeth A. Repasky, and Daniel T. Fisher. “Fever and the Thermal Regulation of Immunity: The Immune System Feels the Heat.” Nature Reviews Immunology 15, no. 6 (May 15, 2015): 335–49. https://doi.org/10.1038/nri3843.
Heinzl, Inge. “Efficient Microbiome Modulation with Phytomolecules.” EW Nutrition, June 9, 2023. https://ew-nutrition.com/pushing-microbiome-in-right-direction-phytomolecules/.
Huntley, Nichole F., John F. Patience, and C. Martin Nyachoti. “Immune Stimulation UPS Maintenance Energy Requirements.” National Hog Farmer.com, September 28, 2017. https://www.nationalhogfarmer.com/hog-health/immune-stimulation-ups-maintenance-energy-requirements.
Kvidera, S. K., E. A. Horst, M. Abuajamieh, E. J. Mayorga, M. V. Sanz Fernandez, and L. H. Baumgard. “Technical Note: A Procedure to Estimate Glucose Requirements of an Activated Immune System in Steers.” Journal of Animal Science 94, no. 11 (November 1, 2016): 4591–99. https://doi.org/10.2527/jas.2016-0765.
Kvidera, S.K., E.A. Horst, M. Abuajamieh, E.J. Mayorga, M.V. Sanz Fernandez, and L.H. Baumgard. “Glucose Requirements of an Activated Immune System in Lactating Holstein Cows.” Journal of Dairy Science 100, no. 3 (March 2017): 2360–74. https://doi.org/10.3168/jds.2016-12001.
LANG, A. “Allicin Inhibits Spontaneous and Tnf-$alpha; Induced Secretion of Proinflammatory Cytokines and Chemokines from Intestinal Epithelial Cells.” Clinical Nutrition, May 2004. https://doi.org/10.1016/s0261-5614(04)00058-5.
Lee, Seung Ho, Sun Young Lee, Dong Ju Son, Heesoon Lee, Hwan Soo Yoo, Sukgil Song, Ki Wan Oh, Dong Cho Han, Byoung Mog Kwon, and Jin Tae Hong. “Inhibitory Effect of 2′-Hydroxycinnamaldehyde on Nitric Oxide Production through Inhibition of NF-ΚB Activation in RAW 264.7 Cells.” Biochemical Pharmacology 69, no. 5 (March 2005): 791–99. https://doi.org/10.1016/j.bcp.2004.11.013.
Liu, S. D., M. H. Song, W. Yun, J. H. Lee, H. B. Kim, and J. H. Cho. “Effect of Carvacrol Essential Oils on Growth Performance and Intestinal Barrier Function in Broilers with Lipopolysaccharide Challenge.” Animal Production Science 60, no. 4 (January 22, 2020): 545–52. https://doi.org/10.1071/an18326.
Liu, S. D., M. H. Song, W. Yun, J. H. Lee, H. B. Kim, and J. H. Cho. “Effect of Carvacrol Essential Oils on Growth Performance and Intestinal Barrier Function in Broilers with Lipopolysaccharide Challenge.” Animal Production Science 60, no. 4 (January 22, 2020): 545–52. https://doi.org/10.1071/an18326.
Mantwill, Elke. “Eiweiß & Immunsystem.” sportärztezeitung, April 10, 2025. https://sportaerztezeitung.com/rubriken/ernaehrung/9197/eiweiss-immunsystem/.
McGilvray, Whitney D, David Klein, Hailey Wooten, John A Dawson, Deltora Hewitt, Amanda R Rakhshandeh, Cornelius F de Lange, and Anoosh Rakhshandeh. “Immune System Stimulation Induced byEscherichia ColiLipopolysaccharide Alters Plasma Free Amino Acid Flux and Dietary Nitrogen Utilization in Growing Pigs1.” Journal of Animal Science 97, no. 1 (October 11, 2018): 315–26. https://doi.org/10.1093/jas/sky401.
Melchior, D., B. Sève, and N. Le Floc’h. “Chronic Lung Inflammation Affects Plasma Amino Acid Concentrations in Pigs.” Journal of Animal Science 82, no. 4 (April 1, 2004): 1091–99. https://doi.org/10.2527/2004.8241091x.
Obled, C. “Amino Acid Requirements in Inflammatory States.” Canadian Journal of Animal Science 83, no. 3 (September 1, 2003): 365–73. https://doi.org/10.4141/a03-021.
Palsson‐McDermott, Eva M., and Luke A. O’Neill. “The Warburg Effect Then and Now: From Cancer to Inflammatory Diseases.” BioEssays 35, no. 11 (September 20, 2013): 965–73. https://doi.org/10.1002/bies.201300084.
Pastorelli, H., J. van Milgen, P. Lovatto, and L. Montagne. “Meta-Analysis of Feed Intake and Growth Responses of Growing Pigs after a Sanitary Challenge.” Animal 6, no. 6 (2012): 952–61. https://doi.org/10.1017/s175173111100228x.
Patience, John. “One of the Most Important Decisions in Swine Production: Dietary Energy Level – Dr. John Patience by The Swine It Podcast Show.” Spotify for Creators, December 2, 2019. https://anchor.fm/swineitpodcast/episodes/One-of-the-most-important-decisions-in-swine-production-dietary-energy-level—Dr–John-Patience-e99j9u.
Plank, Lindsay D., and Graham L. Hill. “Sequential Metabolic Changes Following Induction of Systemic Inflammatory Response in Patients with Severe Sepsis or Major Blunt Trauma.” World Journal of Surgery 24, no. 6 (June 2000): 630–38. https://doi.org/10.1007/s002689910104.
Rakhshandeh, A., and C.F.M. de Lange. “Evaluation of Chronic Immune System Stimulation Models in Growing Pigs.” Animal 6, no. 2 (2012): 305–10. https://doi.org/10.1017/s1751731111001522.
Rakhshandeh, A., and C.F.M. De Lange. “Immune System Stimulation in the Pig: Effect on Performance and Implications for Amino Acid Nutrition.” Essay. In Manipulating Pig Production XIII, 31–46. Werribee, Victoria, Australia: Australasian Pig Science Association Incorporation, 2011.
Rakhshandeh, Anoosh, John K. Htoo, Neil Karrow, Stephen P. Miller, and Cornelis F. de Lange. “Impact of Immune System Stimulation on the Ileal Nutrient Digestibility and Utilisation of Methionine plus Cysteine Intake for Whole-Body Protein Deposition in Growing Pigs.” British Journal of Nutrition 111, no. 1 (January 14, 2014): 101–10. https://doi.org/10.1017/s0007114513001955.
Reeds, P., and F. Jahoor. “The Amino Acid Requirements of Disease.” Clinical Nutrition 20 (June 2001): 15–22. https://doi.org/10.1054/clnu.2001.0402.
Reeds, Peter J, Carla R Fjeld, and Farook Jahoor. “Do the Differences between the Amino Acid Compositions of Acute-Phase and Muscle Proteins Have a Bearing on Nitrogen Loss in Traumatic States?” The Journal of Nutrition 124, no. 6 (June 1994): 906–10. https://doi.org/10.1093/jn/124.6.906.
Rigobelo, E. Cid, and F. A. De Ávila. “Hypoglycemia Caused by Septicemia in Pigs.” Essay. In Hypoglycemia – Causes and Occurrences., 221–38. London, UK: InTechOpen, 2011.
Rowaiye, Adekunle, Gordon C. Ibeanu, Doofan Bur, Sandra Nnadi, Ugonna Morikwe, Akwoba Joseph Ogugua, and Chinwe Uzoma Chukwudi. “Phyto-Molecules Show Potentials to Combat Drug-Resistance in Bacterial Cell Membranes.” Microbial Pathogenesis 205 (August 2025): 107723. https://doi.org/10.1016/j.micpath.2025.107723.
Saravanan, Haribabu, Maida Engels SE, and Muthiah Ramanathan. “Phytomolecules Are Multi Targeted: Understanding the Interlinking Pathway of Antioxidant, Anti Inflammatory and Anti Cancer Response.” In Silico Research in Biomedicine 1 (2025): 100002. https://doi.org/10.1016/j.insi.2025.100002.
Spurlock, M E. “Regulation of Metabolism and Growth during Immune Challenge: An Overview of Cytokine Function.” Journal of Animal Science 75, no. 7 (1997): 1773–83. https://doi.org/10.2527/1997.7571773x.
Suchner, U., K. S. Kuhn, and P. Fürst. “The Scientific Basis of Immunonutrition.” Proceedings of the Nutrition Society 59, no. 4 (November 2000): 553–63. https://doi.org/10.1017/s0029665100000793.
Phytomolecules: Sustainability and Efficiency in Pig Production
Conference Report
By M. Rosenthal, Global Application Manager Swine, EW Nutrition GmbH
Sustainability is essential for the long-term survival of our planet. In pig production, sustainability involves maintaining economically viable outputs while simultaneously safeguarding animal health and welfare and minimizing environmental impact. The goal is to produce pork that is profitable, ethical, and has a minimal ecological footprint.
Phytomolecules, the bioactive constituents of plant-derived essential oils, play a promising role in advancing this goal. With multifunctional gut health benefits including antimicrobial, anti-inflammatory, antioxidant, and digestive-supportive properties, phytomolecules help maintain gut health and reduce the need for antibiotics. By improving feed efficiency, enhancing resilience, and supporting intestinal integrity, phytomolecules contribute to both sustainability and efficiency in pig production systems.
Targeting sustainability in pig production
Achieving sustainability in pig production requires a balanced approach that considers three key perspectives: those of the producer, the pig, and the environment.
For the producer, sustainable pig production must be profitable to ensure the long-term viability of the industry. This includes factors such as efficient feed conversion, optimized production practices, and fair market prices.
Another aspect is the maintenance of animal health and well-being, which is essential for optimal pig performance and can be achieved by providing appropriate housing, nutrition, and veterinary care, as well as minimizing stress and disease.
From an environmental perspective, minimizing negative impacts, such as greenhouse gas emissions, water pollution, and land degradation, is a key objective. Various strategies, such as improved manure management, efficient nutrient utilization, reuse of farm resources like manure and water, and the use of by-products from other industries as feed ingredients, can be applied.
Strategy for efficient pig production
Historically, pig production has relied heavily on the use of antibiotics to control enteric pathogens, promote gut health, and enhance growth. While effective in the short term, this practice led to unintended consequences, including the emergence of antimicrobial resistance (amr), disruption of microbiota across multiple organ systems, difficulties in manure management, and environmental contamination.
These outcomes triggered societal concern, regulatory interventions, and economic pressure, prompting a shift away from routine antibiotic use. The industry now faces increasing expectations for environmentally responsible practices, reduced dependence on antibiotics, and cost-effective, sustainable solutions.
Achieving both efficiency and sustainability in pig production requires a holistic, system-wide approach that includes an innovative, solution-oriented mindset, optimized management practices, and the adoption of effective gut health antibiotic alternatives.
The foundation of efficiency – the gut
The pigs gastrointestinal tract is the largest and most vulnerable interface between the pig and its external environment. It is a highly organized ecosystem comprised of epithelial cells, the mucosal immune system, and a diverse microbiome consisting of both beneficial commensal microbes and potentially harmful pathogens.
The functions of the gut include nutrient absorption, chemosensing of nutrients and other compounds, immune defence, and balancing the highly diverse microbiome within this complex environment (Furness et al. , 2013). Disruption of this ecosystems homeostasis can impair not only gut function and health but also negatively affect the overall well-being and growth efficiency of the pig.
When evaluating antibiotic alternatives to support this ecosystems homeostasis in the face of challenges, considerations include safety for humans, animals, and the environment, cost-effectiveness, antimicrobial efficacy, the ability to increase nutrient availability, and to modulate immune activation and inflammation.
Functional feed additives commonly utilized in pig nutrition, alone or in combination, include organic acids, probiotics, immunoglobulins, medium-chain fatty acids, and phytomolecules.
Phytomolecules: supporting gut health and performance
Phytomolecules are the bioactive components of plant-derived essential oils. Due to the variability in phytomolecule content and the presence of volatile and astringent components in essential oil extracts, utilizing commercial phytomolecule products is recommended. Proprietary formulations utilize encapsulation or matrix technology to protect the phytomolecules from damage or loss during storage, processing, and passage through the stomach.
Extensive research in humans and animals has identified phytomolecules as having antimicrobial, anti-inflammatory, antioxidative, and coccidiostatic properties. They enhance digestibility and immunity, promote gut health through differential modulation of bacterial populations, and reduce inflammation and oxidative stress (Brenes et al., 2010; Puvaca et al. , 2013; Chitprasert et al., 2014). Phytomolecules most researched and utilized in pig feed additives to date include terpenes (e. G., carvacrol and thymol) and phenylpropenes (e.g., cinnamaldehyde and eugenol).
1. Direct antimicrobial activity of phytomolecules
Phytomolecules such as carvacrol and thymol provide broad-spectrum antimicrobial activities against Gram- and Gram+ bacteria, fungi, and yeast and are regarded as promising alternatives to antibiotics in swine production systems (Lambert et al., 2001; Delaquis et al., 2002; Abbaszadeh et al., 2014).
Phytomolecules directly target bacterial cells through multiple mechanisms, with the cell wall and membrane being major sites of action. The lipophilic structure of phytomolecules enables their entry through bacterial membranes among the fatty acid chains, causing the cell wall and membranes to expand and become more fluid. This damage collapses the cell wall and cytoplasmic membrane, resulting in the destruction of membrane proteins, the coagulation of the cytoplasm, and a reduction in proton motive force. The result is leakage of vital intracellular contents and death of the bacterial cell (Cox et al., 1998; Faleiro, 2011; Nazzaro et al., 2013; Yap et al., 2014). For example, thymol and carvacrol can damage the outer membrane of Salmonellatyphimurium and Escherichia coli o157: h7 (Helander et al., 1998).
A further direct antimicrobial action involves phytomolecules acting as trans-membrane carriers, exchanging a hydroxyl proton for a potassium ion, resulting in dissipation of the ph gradient and electrical potential over the bacterial cytoplasmic membrane. The result is a reduced proton motive force and the depletion of the intracellular adenosine triphosphate (APT) pools. Loss of potassium further inhibits bacterial function as it is needed for the activation of cytoplasmic enzymes to maintain osmotic pressure and regulate intracellular pH. (Wendakoon et al., 1995).
In summary, the primary direct antimicrobial mechanism of action for terpene and phenylpropene phytomolecules is related to their effects on cell walls and cytoplasmic membranes, and energy metabolism of pathogenic bacteria.
2. Indirect antimicrobial activity of phytomolecules
Phytomolecules indirectly impact the physiological functioning and virulence capability of pathogenic bacteria through the interference of quorum-sensing (QS). QS involves pathogenic bacteria producing signaling molecules that are released based on cell numbers. The detection of these molecules regulates pathogen population behavior such as attachment, biofilm formation, and motility, i. e. , virulence (Greenberg, 2003; Joshi et al., 2016).
QS mechanisms require signal synthesis, signal accumulation, and signal detection, providing three opportunities for QS inhibitors to disrupt pathogenic bacteria from causing disease (Czajkowski and Jafra, 2009; Lasarre and Federle, 2013). Eugenol and carvacrol have been extensively studied for their QS inhibition activities (Zhou et al., 2013; Burt et al., 2014).
3. Combinations increase efficacy
Additional antimicrobial effects can be seen when different phytomolecules are combined, and/or applied with other functional additives such as organic acids (Souza et al., 2009; Hulankova and Borilova, 2011). Zhou et al. (2007) reported that carvacrol or thymol in combination with acetic or citric acid had a better efficacy against S. typhimurium when compared to the individual phytomolecule or organic acid. In recent studies, results have shown in vivo efficacy of such synergistic dietary strategies in pigs (Diao et al., 2015; Balasubramanian et al., 2016). The combined inclusion of phytomolecules and organic acids in pig diets before slaughter may hinder Salmonella shedding and seroprevalence (Walia et al., 2017; Noirrit et al., 2016).
4. Phytomolecules are more than antimicrobials
In addition to acting as antimicrobials, phytomolecules enhance production efficiency through multiple complementary mechanisms, including direct anti-inflammatory, antioxidative, digestive, and gut barrier-supportive effects.
Anti-inflammatory effects: Gut inflammation in pigs not only compromises intestinal function and barrier integrity but also has a direct negative impact on growth performance and overall health. Chronic or excessive immune activation diverts energy away from productive processes such as growth and feed efficiency.
Phytomolecules have demonstrated the ability to modulate immune responses by influencing key cell-signalling pathways involved in inflammation. For example, compounds such as cinnamaldehyde and carvacrol can modulate the activity of critical transcription factors, including nuclear factor erythroid 2 2-related factor 2 (Nrf2) and nuclear factor kappa B (NF-κB). Through this dual action, phytomolecules can simultaneously activate antioxidant defences and suppress pro-inflammatory signalling, thereby reducing intestinal inflammation and supporting improved performance outcomes (Krois-mayr et al., 2008; Wondrak et al., 2010; Zou et al., 2016).
Antioxidant effects: oxidative stress is a major biological challenge in modern swine production systems, where high-performance animals are frequently exposed to stressors such as weaning, disease challenges, heat stress, mycotoxin exposure, transport, and overcrowding. These stressors promote the generation of reactive oxygen species (ROS), and when ROS production exceeds the capacity of the pig’s antioxidant defence systems, oxidative stress occurs.
This imbalance can negatively affect growth, immunity, muscle integrity, feed intake, milk yield, and reproductive performance, including increased abortion rates in gestating sows (Zhou et al., 2013; Burt et al., 2014). As a result, there is growing interest in the use of natural antioxidant compounds, particularly phytomolecules, to counteract these detrimental effects. For example, carvacrol and thymol (1:1 ratio) at 100 mg/kg dietary supplementation reduced weaning-associated oxidative stress by decreasing TNF-α mRNA expression in the intestinal mucosa (Wei et al., 2017).
Additionally, carvacrol supplementation in the diets of late gestation and lactating sows under oxidative stress conditions significantly improved piglet performance (Tan et al., 2015).
Digestive function: The gastrointestinal tract functions not only as a site for nutrient absorption but also as a sensory organ. Specialized chemosensors in the gut monitor the concentration and composition of nutrients, playing a crucial role in the regulation of digestive enzyme secretion, gut peptide release, feed intake, and nutrient absorption and metabolism.
Studies in weaner piglets have shown that certain phytomolecules can stimulate the secretion of digestive enzymes and enhance gastrointestinal function (Maenner et al., 2011; Li et al., 2012).
Tight junctions and gut barrier integrity: The intestinal epithelium functions as a highly dynamic and selective barrier, facilitating the absorption of fluids and solutes while preventing the translocation of pathogens and toxins into underlying tissues. This barrier function occurs through intercellular tight junctions. During episodes of mucosal inflammation, the integrity of these junctions can be compromised, leading to increased intestinal permeability, reduced nutrient absorption, and systemic immune activation and inflammation.
Research has shown that phytomolecules can enhance transepithelial electrical resistance and upregulate the expression of tight junction proteins, reducing epithelial permeability and maintaining a functional barrier, even under inflammatory conditions (Yu et al., 2020; Kim and Kim, 2019).
Sustainable efficiency in pig production supported by in-feed phytomolecules
As the pig industry moves away from reliance on in-feed antibiotics, the need for sustainable, efficient, and health-focused production strategies has never been greater. Modern pig production systems must respond to societal expectations, regulatory mandates, and environmental pressures, while still maintaining profitability and high animal welfare standards.
Central to this transformation is a holistic approach-one that includes a shift in mindset among stakeholders, optimized management across all production domains, and the strategic use of effective antibiotic alternatives. The gastrointestinal tract, as the core of nutrient absorption and immune defence, is a critical control point for supporting health and performance.
Phytomolecules and other functional feed additives have demonstrated potential to enhance gut integrity, reduce inflammation, combat oxidative stress, and improve nutrient utilization. While no single solution can fully replace antibiotics, targeted combinations of these compounds have shown the most consistent success in promoting gut health and sustainable performance.
With continued innovation, collaboration, and science-based application of these alternatives, the industry is well-positioned to achieve its goals of profitable, ethical, and ecologically responsible pork production for the future.
References
Abbaszadeh, S., A. Sharifzadeh, H. Shokri, A. Khosravi, and A. Abbaszadeh. 2014. “Antifungal Efficacy of Thymol, Carvacrol, Eugenol and Menthol as Alternative Agents to Control the Growth of Food-Relevant Fungi.” Journal de Mycologie Médicale 24 (2): 51–56. Balasubramanian, B., J. W. Park, and I. H. Kim. 2016. “Evaluation of the Effectiveness of Supplementing Micro-Encapsulated Organic Acids and Essential Oils in Diets for Sows and Suckling Piglets.” Italian Journal of Animal Science 15 (4): 626–33. Baschieri, A., M. D. Ajvazi, J. L. F. Tonfack, L. Valgimigli, and R. Amorati. 2017. “Explaining the Antioxidant Activity of Some Common Non-Phenolic Components of Essential Oils.” Food Chemistry 232: 656–63. Berchieri-Ronchi, C., S. Kim, Y. Zhao, C. Correa, K.-J. Yeum, and A. Ferreira. 2011. “Oxidative Stress Status of Highly Prolific Sows During Gestation and Lactation.” Animal 5 (11): 1774–79. Brenes, A., and E. Roura. 2010. “Essential Oils in Poultry Nutrition: Main Effects and Modes of Action.” Animal Feed Science and Technology 158 (1): 1–14. Burt, S. A., V. T. Ojo-Fakunle, J. Woertman, and E. J. Veldhuizen. 2014. “The Natural Antimicrobial Carvacrol Inhibits Quorum Sensing in Chromobacterium violaceum and Reduces Bacterial Biofilm Formation at Sub-Lethal Concentrations.” PLoS One 9 (4): e93414. Chitprasert, P., and P. Sutaphanit. 2014. “Holy Basil (Ocimum sanctum Linn.) Essential Oil Delivery to Swine Gastrointestinal Tract Using Gelatine Microcapsules Coated with Aluminium Carboxymethyl Cellulose and Beeswax.” Journal of Agricultural and Food Chemistry 62 (52): 12641–48. Cox, S., J. Gustafson, C. Mann, J. Markham, Y. Liew, and R. Hartland, et al. 1998. “Tea Tree Oil Causes K⁺ Leakage and Inhibits Respiration in Escherichia coli.” Letters in Applied Microbiology 26 (5): 355–58. Czajkowski, R., and S. Jafra. 2009. “Quenching of Acyl-Homoserine Lactone-Dependent Quorum Sensing by Enzymatic Disruption of Signal Molecules.” Acta Biochimica Polonica 56 (1): 1–16. Delaquis, P. J., K. Stanich, B. Girard, and G. Mazza. 2002. “Antimicrobial Activity of Individual and Mixed Fractions of Dill, Cilantro, Coriander and Eucalyptus Essential Oils.” International Journal of Food Microbiology 74 (1): 101–9. Diao, H., P. Zheng, B. Yu, J. He, X. Mao, J. Yu, et al. 2015. “Effects of Benzoic Acid and Thymol on Growth Performance and Gut Characteristics of Weaned Piglets.” Asian-Australasian Journal of Animal Sciences 28 (6): 827–35. Faleiro, M. 2011. “The Mode of Antibacterial Action of Essential Oils.” In Science Against Microbial Pathogens: Communicating Current Research and Technological Advances, vol. 2, 1143–56. Badajoz, Spain: Formatex Research Center. Furness, J., L. Rivera, and H. J. Cho, et al. 2013. “The Gut as a Sensory Organ.” Nature Reviews Gastroenterology & Hepatology 10: 729–40. Greenberg, E. P. 2003. “Bacterial Communication and Group Behavior.” Journal of Clinical Investigation 112 (9): 1288–90. Helander, I. M., H.-L. Alakomi, K. Latva-Kala, T. Mattila-Sandholm, I. Pol, E. J. Smid, et al. 1998. “Characterization of the Action of Selected Essential Oil Components on Gram-Negative Bacteria.” Journal of Agricultural and Food Chemistry 46 (9): 3590–95. Hulankova, R., and G. Borilova. 2011. “In Vitro Combined Effect of Oregano Essential Oil and Caprylic Acid Against Salmonella Serovars, Escherichia coli O157:H7, Staphylococcus aureus and Listeria monocytogenes.” Acta Veterinaria Brno 80 (4): 343–48. Joshi, J. R., N. Khazanov, H. Senderowitz, S. Burdman, A. Lipsky, and I. Yedidia. 2016. “Plant Phenolic Volatiles Inhibit Quorum Sensing in Pectobacteria and Reduce Their Virulence by Potential Binding to ExpI and ExpR Proteins.” Scientific Reports 6: 38126. Kim, M. S., and J. Y. Kim. 2019. “Cinnamon Subcritical Water Extract Attenuates Intestinal Inflammation and Enhances Intestinal Tight Junction in a Caco-2 and RAW264.8 Co-Culture Model.” Food & Function 20: 4350–60. Kroismayr, A., J. Sehm, M. Pfaffl, K. Schedle, C. Plitzner, and W. Windisch.2008. “Effects of Avilamycin and Essential Oils on mRNA Expression of Apoptotic and Inflammatory Markers and Gut Morphology of Piglets.” Czech Journal of Animal Science 53: 377–87. Lambert, R., P. N. Skandamis, P. J. Coote, and G. J. Nychas. 2001. “A Study of the Minimum Inhibitory Concentration and Mode of Action of Oregano Essential Oil, Thymol and Carvacrol.” Journal of Applied Microbiology 91 (3): 453–62. LaSarre, B., and M. J. Federle. 2013. “Exploiting Quorum Sensing to Confuse Bacterial Pathogens.” Microbiology and Molecular Biology Reviews 77 (1): 73–111. Li, P., X. Piao, Y. Ru, X. Han, L. Xue, and H. Zhang. 2012. “Effects of Adding Essential Oil to the Diet of Weaned Pigs on Performance, Nutrient Utilization, Immune Response and Intestinal Health.” Asian-Australasian Journal of Animal Sciences 25 (11): 1617–26. Maenner, K., W. Vahjen, and O. Simon. 2011. “Studies on the Effects of Essential-Oil-Based Feed Additives on Performance, Ileal Nutrient Digestibility, and Selected Bacterial Groups in the Gastrointestinal Tract of Piglets.” Journal of Animal Science 89 (7): 2106–12. Nazzaro, F., F. Fratianni, L. De Martino, R. Coppola, and V. De Feo. 2013. “Effect of Essential Oils on Pathogenic Bacteria.” Pharmaceuticals 6 (12): 1451–74. Noirrit, M., and F. Philippe. 2016. “Reduction of Salmonella Prevalence on Sows and Finishing Pigs by Use of a Protected Mix of Organic Acids and Essential Oils in the Feed of Lactating Sows and Weaned Piglets.” Journées Recherche Porcine 48: 351–52. Puvaca, N., V. Stanacev, D. Glamocic, J. Levic, L. Peric, and D. Milic. 2013. “Beneficial Effects of Phytoadditives in Broiler Nutrition.” World’s Poultry Science Journal 69 (1): 27–34. Souza, E. L., J. C. Barros, M. L. Conceiçao, N. J. Gomes Neto, and A. C. V. Costa.2009. “Combined Application of Origanum vulgare L. Essential Oil and Acetic Acid for Controlling the Growth of Staphylococcus aureus in Foods.” Brazilian Journal of Microbiology 40 (2): 387–93. Tan, C., H. Wei, H. Sun, J. Ao, G. Long, S. Jiang, et al. 2015. “Effects of Dietary Supplementation of Oregano Essential Oil to Sows on Oxidative Stress Status, Lactation Feed Intake of Sows, and Piglet Performance.” BioMed Research International 2015: Article ID 941754. Walia, K., H. Argüello, H. Lynch, F. C. Leonard, J. Grant, D. Yearsley, et al. 2017. “Effect of Strategic Administration of an Encapsulated Blend of Formic Acid, Citric Acid, and Essential Oils on Salmonella Carriage, Seroprevalence, and Growth of Finishing Pigs.” Preventive Veterinary Medicine 137: 28–35. Wei, H. K., H. X. Xue, Z. Zhou, and J. Peng. 2017. “A Carvacrol-Thymol Blend Decreased Intestinal Oxidative Stress and Influenced Selected Microbes Without Changing the Messenger RNA Levels of Tight Junction Proteins in Jejunal Mucosa of Weaning Piglets.” Animal 11 (2): 193–201. Wendakoon, C. N., and M. Sakaguchi. 1995. “Inhibition of Amino Acid Decarboxylase Activity of Enterobacter aerogenes by Active Components in Spices.” Journal of Food Protection 58 (3): 280–83. Wondrak, G. T., N. F. Villeneuve, S. D. Lamore, A. S. Bause, T. Jiang, and D. D. Zhang. 2010. “The Cinnamon-Derived Dietary Factor Cinnamic Aldehyde Activates the Nrf2-Dependent Antioxidant Response in Human Epithelial Colon Cells.” Molecules 15 (5): 3338–55. Yap, P. S. X., B. C. Yiap, H. C. Ping, and S. H. E. Lim. 2014. “Essential Oils, a New Horizon in Combating Bacterial Antibiotic Resistance.” Open Microbiology Journal 8 (1). Yu, J., Y. Song, B. Yu, J. He, P. Zheng, X. Mao, Z. Huang, Y. Luo, J. Luo, H. Yan, Q. Wang, H. Wang, and D. Chen. 2020. “Tannic Acid Prevents Post-Weaning Diarrhea by Improving Intestinal Barrier Integrity and Function in Weaned Piglets.” Journal of Animal Science and Biotechnology 11: 87. Zhou, F., B. Ji, H. Zhang, H. Jiang, Z. Yang, J. Li, et al. 2007. “Synergistic Effect of Thymol and Carvacrol Combined with Chelators and Organic Acids Against Salmonella Typhimurium.” Journal of Food Protection 70 (7): 1704–9. Zhou, L., H. Zheng, Y. Tang, W. Yu, and Q. Gong. 2013. “Eugenol Inhibits Quorum Sensing at Subinhibitory Concentrations.” Biotechnology Letters 35 (4): 631–37. Zou, Y., J. Wang, J. Peng, and H. Wei. 2016. “Oregano Essential Oil Induces SOD1 and GSH Expression Through Nrf2 Activation and Alleviates Hydrogen Peroxide-Induced Oxidative Damage in IPEC-J2 Cells.” Oxidative Medicine and Cellular Longevity 2016: Article ID 5987183.
The big challenge: Keeping sows healthy and productive – Part 2 Nutritional interventions – Phytomolecules
Dr. Inge Heinzl – Editor of EW Nutrition, and Dr. Merideth Parke – Global Application Manager for Swine, EW Nutrition
The first of the two articles focused on general aspects to be observed to achieve a particular stock of healthy and well-performing sows, as well as high productivity on the farm. In addition to general measures, feed supplements can be used to further support the sows. Phytomolecules with characteristics supporting gut and overall health are effective for this purpose.
Phytomolecules – how can they help?
Phytogenics, also known as phytomolecules, are plant-derived, natural bioactive compounds that promote livestock health and well-being, as well as improve growth performance and production efficiency. Phytomolecules encompass a diverse range of compounds, including terpenes, phenols, glycosides, saccharides, aldehydes, esters, and alcohols.
The literature describes some of their effects, including stimulation of digestive secretions, immune stimulation and anti-inflammatory activity, intestinal microflora modulation, and antioxidant effects (Durmic and Blanche, 2012; Ehrlinger, 2007; Zhao et al., 2023), as well as estrogenic and hyperprolactinemic properties (Farmer, 2018) and effects on colostrum and milk porcine sensory profiles (Val-Laillet et al., 2018). They represent exciting antibiotic alternatives in swine production (Omonijo et al., 2018).
1. Phytomolecules modulate intestinal microbiota
Phytomolecules are microbiome modulators through different mechanisms. They can directly impact pathogenic bacteria by damaging the cell membrane, cell wall, or cytoplasm, interrupting the anion exchange, resulting in changes to cellular pH, and inhibiting the cell’s energy production system. Additionally, phytomolecules interfere with the virulence capacity of pathogenic bacteria through the indirect quorum quenching mechanism. (Rutherford and Bassler, 2012).
The favorable consequence of this differential microbial modulation is maintaining gut microbiome diversity, shifting it to a bacterial population with reduced pathogenic and increased beneficial microbes.
Proof of Ventar D’s pathogen-inhibiting effect
An in vitro study evaluated the effect of Ventar D on pathogenic Clostridium perfringens and beneficial Lactobacillus spp.
Process
To test the effect of Ventar D on four different beneficial Lactobacillus spp., and pathogenic Clostridium perfringens, the phytogenic formulation (Ventar D) was added to the respective culture medium in the following concentrations: 0 µg/mL – control, 500 µg/mL (only C. perfr.), 750 µg/mL, 1000 mg/mL (only C. perfr.), and 1250 µg/mL.
After cultivating the bacteria in the culture medium, the colony-forming units (CFU) were counted.
Results and discussion
The study demonstrated a dose-dependent decrease in the Clostridium perfringens population. At the lowest tested concentration (500 µg/mL), Ventar D’s antimicrobial effect was already detectable; at 750 µg/mL, scarce colonies were observed; and at 1000 µg/mL, C. perfringens could no longer grow.
Figure 1: Effect of Ventar D on Clostridium perfringens
In contrast, even at higher concentrations of Ventar D, the beneficial L. agilis S73 and L. agilis S1 populations were only mildly affected, and L. casei and L. plantarum were unaffected.
Figure 2: Effect of Ventar D on Lactobacillus spp.
These findings confirm the differential antimicrobial activity of Ventar D’s formulation, specifically a bactericidal effect on pathogenic C. perfringens populations and a mild to no inhibition of beneficial Lactobacillus spp.
2. Phytomolecules improve intestinal integrity
The gut barrier is semipermeable and is responsible for immune sensing and regulating the movement of nutrients and undesirable microbes and substances.
The “gatekeepers” are tight junctions (TJ), adherent junctions (AJ), and desmosomes situated between the intestinal enteric cells (IEC). The tight junctions regulate the transport of small molecules and ions. The adherent junctions and desmosomes maintain the integrity of the intestinal barrier by keeping the IECs together through cell-adhesion bonds.
Oxidative stress resulting from factors such as heat stress or fat oxidation in the feed, as well as dysbacteriosis caused by changes in diet, out-of-feed events, poor dietary formulation, or bacterial contamination, can compromise the integrity of these critical adhesions and junctions between enterocytes.
The support of these tight junctions prevents bacteria and toxins from passing into the organism. Besides reducing disease occurrence, it also reduces the activation of the immune system and inflammatory processes. Ingested nutrients can be used for growth and need not be spent for the defense of the organism.
Proof of Ventar D’s gut barrier-stabilizing effect
An experiment was conducted to determine the level of tight junction gene expression biomarkers closely related to gut integrity.
Process
The experiment was conducted in broilers. They were fed 100 g of Ventar D/ t of feed, and the gene expression of Claudin and Occludin was measured (the higher the gene expression, the higher the level of gut barrier damage).
Figure 3: Effect of Ventar D on gut barrier function
Results
The lower levels of both gut tight junction gene expression biomarkers, Claudin and Occludin, in Ventar D-supplemented birds support a lower level of damage and a more robust gut barrier function (Figure 3).
3. Phytomolecules act as antioxidants
As mentioned, oxidative stress can disrupt gut barrier function and negatively impact the health of sows and piglets. Therefore, it is vital to scavenge reactive oxygen species (ROS) to reduce the damage these free radicals can cause to enterocytes and tight junctions.
Proof of Ventar D’s antioxidant effect in vitro
In this case, an in vitro trial was conducted to show Ventar D’s antioxidant effects.
Process
Ventar D’s antioxidant activity was tested in vitro using the ORAC (Oxygen Radical Absorbent Capacity) test. The ORAC test measures the antioxidant activity of a compound compared to that of the Vitamin E analog Trolox.
Result
The components in Ventar D demonstrated its capacity as an antioxidant, with a more substantial effect than the Vitamin E analog Trolox (see Figure 4).
Figure 4: Antioxidant capacity of Ventar D compared to Vit. E analogue (AUC – Area under curve)
4. Phytomolecules decrease inflammation
In intensive production, animals face daily inflammation associated with various stressors, including gut incidents and intestinal dysbiosis, social hierarchy-associated fighting resulting in musculoskeletal or skin injuries, farrowing and lactation trauma to reproductive organs, and diseases affecting any system in the pig.
Animals with high-performance expectations, such as gestating, farrowing, and lactating sows, are particularly susceptible to high nutrient diversion, which can lead to inflammation and activation of the immune system. To mitigate the excessive continuation of inflammatory processes, phytomolecules with anti-inflammatory effects can be utilized.
Proof of Ventar D’s anti-inflammatory effect in vitro
The anti-inflammatory effect of Ventar D was shown in an in vitro trial conducted in the Netherlands.
Process
For the trial, cells from mice (Murine macrophages, RAW264.7) were stressed with lipopolysaccharides (LPS, Endotoxin) from E. coli O111:B4 (0.25 µg/ml) to provoke an immune response. To evaluate the effects of Ventar D, two different concentrations (50 and 200 ppm) were tested, and the levels of NF-κB, IL-6, and IL-10 were determined. IL-6 and IL-10 could be measured directly using specific ELISA kits, whereas, in the case of NF-κB activity, an enzyme induced by NF-κB (secreted embryonic alkaline phosphatase – SEAP) was used for measurement. The trial design was as follows (Figure 5):
Figure 5: Trial design
Results
The trial results showed a dose-dependent reduction of NF-κB activity in LPS-stimulated mouse cells, with 11% and 54% reductions at 50 and 200 ppm Ventar D, respectively. The pro-inflammatory cytokine IL-6 was downregulated, and the anti-inflammatory cytokine IL-10 was upregulated by 84% and 20%, respectively, resulting in a decrease in the IL-6 to IL-10 ratio. This ratio is essential in balancing the pro- and anti-inflammatory outcomes of cellular signaling.
Figure 6: Activity of NFκB
Figure 7: IL-6/IL-10 ratio
5. Phytomolecules improve production performance and efficiency
The intensive production systems of today encompass many factors that create stress in the animals. Phytomolecules exhibiting the positive characteristics mentioned in points 1 to 4 result in better performance in animals.
In pigs in suboptimal conditions, the antimicrobial effect of phytomolecules is the most important. However, in pigs held under optimal conditions and with extraordinary growth, the antioxidant and anti-inflammatory effects are most relevant. Anabolic processes, driven by strong growth, increase oxidative stress, while non-infectious inflammations burden the immune system.
Proof of Ventar D’s performance-promoting effect in pigs
To evaluate growth-promoting effects in pigs, a study was conducted on a commercial farm in the United States.
Process
A total of 532 approx. 24-day-old weaned piglets were housed in 28 pens, each containing 19 non-castrated males or gilts. Piglets were blocked by body weight and fed a three-phase feeding program (Table 1). Phase 1 and 2 diets were pellets, and phase 3 was mash. Diets were based on corn and soybeans, and a concentrate including soy protein concentrate, whey permeate, and fish meal was added in phases 1 and 2, at a ratio of 50% of the total feed in phase 1 and 25% in phase 2. No feed medication was used in this trial.
Table 1: Feeding scheme and product application
Trial groups
Feeding Phase 1 (day1 – day 14)
Feeding Phase 2 (day 15 – day 24)
Feeding Phase 3
Control
No additive
No additive
No additive
Ventar D
Ventar D 200 g/MT
Ventar D 200 g/MT
Ventar D 200 g/MT
Results
Adding Ventar D increased final body weight and improved FCR (see Figures 8 to 10). Furthermore, the addition of Ventar D to the feed reduced mortality.
Figures 8-10: Performance of piglets fed Ventar D in comparison to a negative control
Phytomolecules can help to keep sows healthy and productive
Intensive animal production places a significant strain on animal organisms. High stocking density often accompanies high pathogenic pressure and stress, and high growth performance can lead to increased oxidative stress and inflammation. It isn’t easy to keep all challenges under control. However, phytomolecules can be a solution as their modes of action cover different relevant topics.
References
Durmic, Z., and D. Blache. “Bioactive Plants and Plant Products: Effects on Animal Function, Health and Welfare.” Animal Feed Science and Technology 176, no. 1–4 (September 2012): 150–62. https://doi.org/10.1016/j.anifeedsci.2012.07.018.
Farmer, Chantal. “Nutritional Impact on Mammary Development in Pigs: A Review.” Journal of Animal Science 96, no. 9 (June 15, 2018): 3748–56. https://doi.org/10.1093/jas/sky243.
Omonijo, Faith A., Liju Ni, Joshua Gong, Qi Wang, Ludovic Lahaye, and Chengbo Yang. “Essential Oils as Alternatives to Antibiotics in Swine Production.” Animal Nutrition 4, no. 2 (June 2018): 126–36. https://doi.org/10.1016/j.aninu.2017.09.001.
Rutherford, S. T., and B. L. Bassler. “Bacterial Quorum Sensing: Its Role in Virulence and Possibilities for Its Control.” Cold Spring Harbor Perspectives in Medicine 2, no. 11 (November 1, 2012). https://doi.org/10.1101/cshperspect.a012427.
Val-Laillet, David, J Stephen Elmore, David Baines, Peter Naylor, and Robert Naylor. “Long-Term Exposure to Sensory Feed Additives during the Gestational and Postnatal Periods Affects Sows’ Colostrum and Milk Sensory Profiles, Piglets’ Growth, and Feed Intake1.” Journal of Animal Science, June 29, 2018. https://doi.org/10.1093/jas/sky171.
Zhao, Bi-Chen, Tian-Hao Wang, Jian Chen, Bai-Hao Qiu, Ya-Ru Xu, Qing Zhang, Jian-Jie Li, Chun-Jiang Wang, Qiu-Feng Nie, and Jin-Long Li. “Effects of Dietary Supplementation with a Carvacrol–Cinnamaldehyde–Thymol Blend on Growth Performance and Intestinal Health of Nursery Pigs.” Porcine Health Management 9, no. 24 (May 23, 2023). https://doi.org/10.1186/s40813-023-00317-x.
The Science Behind Phytogenics
Conference Report
Essential oils, secondary plant compounds, phytogenics – all these expressions can be found in the context of animal feed. In the following, Dr. Sabiha Kadari, Regional Technical Director Southeast Asia/Pacific at EW Nutrition, will show the difference between essential oils and phytomolecules and the science behind phytogenics.
Essential oils and phytomolecules– not the same
Let us first show what are essential oils using the example of oregano oil. Essential oils are extracted from plants and unpurified mixes of different phytomolecules. The raw oregano oil extract contains carvacrol, thymol, P-cymene, and several other phytomolecules. The concentration and composition of these phytomolecules can vary significantly, depending on factors such as geographical origin, seasonal variations, plant part, plant growth stage and harvest time, extraction methods, and post-harvest processing. As a result, there can be significant batch-to-batch variations, resulting in differences in animal performance. Furthermore, there is the potential for the presence of undesirable contaminants.
In contrast, phytomolecules are the active ingredients in essential oils or other plant materials. They are clearly defined as one active compound (IUPAC name/CAS number) by their unique chemical structures, such as carvacrol. By focusing on specific active compounds, standardized products don’t have batch-to-batch variation, enhancing consistent animal performance.
Stringent screening processes
To yield the best phytogenic formulations for animal production, a rigorous screening process is required:
The initial screening process consists of ensuring the bioactives are generally recognized as safe (GRAS) by the US Department of Agriculture and approved by the European Food Safety Authority (EFSA). This step is crucial to ensure that any compounds used in formulations do not pose health risks to animals or humans.
In addition to being selected for their chemical-physical properties, which play a significant role in determining how well the phytogenics will perform in various applications, and a thorough cost-benefit analysis, the phytogenics are mapped for their following biological activities.
Antioxidant
Phytomolecules exert their antioxidant effects through various mechanisms, including scavenging free radicals. The ORAC (Oxygen Radical Absorbance Capacity) test is widely regarded as a gold standard for measuring the antioxidant potential of phytomolecules. It quantitatively assesses the ability of compounds to scavenge free radicals, providing a reliable comparison against a known standard, specifically Trolox, a vitamin E analog. Trolox has well-documented antioxidant properties, making it a reliable benchmark for evaluating the effectiveness of other antioxidants.
Antimicrobial
Incorporating a comprehensive approach to testing the antibacterial properties of phytogenics is essential for developing effective feed additives. The antibacterial properties should not only be tested against harmful enteropathogenic bacteria, such as Clostridium perfringens, E. coli, and Salmonella. It should also be evaluated if beneficial species such as Lactobacilli, the proliferation of which is wanted, are preserved.
By evaluating both pathogenic and beneficial bacteria, researchers can ensure that phytogenic formulations support optimal gut health and reduce the reliance on antibiotics.
Anti-inflammatory
Anti-inflammatory properties also help to modulate the gut-associated immune system and mitigate excessive immune response so that animals can allocate more energy towards growth and production. This shift is vital for optimizing feed conversion ratios and overall performance.
Dr. Kadari noted that “EW Nutrition uses nuclear factor kappa beta (NFkß), which regulates the expression of various pro-inflammatory cytokines, and interleukin 6 (pro-inflammatory) and 10 (anti-inflammatory) cytokines as biomarkers, for measuring anti-inflammatory activity. A reduction in NFkß and the ratio of IL-6/ IL-10 indicates a decrease in inflammatory response.”
Anti-conjugation
Conjugation is a common mechanism of horizontal gene transfer that is instrumental in spreading antibiotic resistance between bacteria. “Most resistance genes are found on mobile genetic elements named plasmids and primarily spread by conjugation,” explained Dr. Kadari.
Cell stress of bacteria modulates the conjugation frequency. Among these stressors are antimicrobial phytogenics. The goal is to keep the conjugation frequency below the one that could occur under unchallenged conditions.
Figure 1: High throughput screening allows EW Nutrition researchers to quickly conduct millions of chemical, genetic, or pharmacological tests
Delivery mechanism
Lastly, to optimize the benefit of the selected phytogenics and deliver consistent results, the substances must be protected by, e.g., encapsulation to ensure homogenous distribution in feed and thermostability in pelleted feed. A special delivery system provides for the targeted release of the active ingredients within the organism, specifically ensuring that these compounds are effectively utilized within the body rather than eliminated through the feces. This is crucial for optimizing their benefits in animal production.
Phytomolecules are an essential support in antibiotic reduction
“Phytogenics are increasingly recognized as effective alternatives in antimicrobial reduction programs. The combination of stringent screening processes alongside rigorous in vitro and in vivo testing is essential for ensuring that phytogenics deliver optimal and consistent performance in animal production,” noted Dr. Kadari.
EW Nutrition’s Swine Academies took place in Ho Chi Minh City and Bangkok in October 2024. Dr. Sabiha Kadari, Regional Technical Director at EW Nutrition SEAP, was one of the highly experienced speakers of EW Nutrition. With expertise in feed cost optimization, feed additive management, audits, and lab support, she provides customized technical solutions and troubleshooting challenges for customers.
Optimising Weaner Performance
Conference report
To optimize weaner performance, it is helpful to understand the stressful situation the piglets are facing. In contrast to weaning in nature, which occurs gradually until completion at approximately 4-5 months, weaning in intensive pig operations is an acute process, typically occurring at 3-4 weeks of age. This critical phase subjects piglets to multiple stressors, which can have cumulative effects on their health and development.
Furthermore, the weaning process usually coincides with a decline in the levels of maternally derived antibodies. As these antibody levels decrease, piglets become increasingly susceptible to infections, particularly during the stressful transition to solid food and movement from the sow to the new nursery environment. Managing the weaning process carefully is crucial to minimize stress and support immune function.
Weaning factors that influence a successful weaning
Several aspects must be considered to provide the weaning piglets with the best conditions, and diverse measures must be taken. These measures range from the social environment to nutrition, hygiene, and the people dealing with the pigs.
Social dynamics
When forming nursery groups, aim to keep pigs in these groups as long as possible. Moving all pigs to their new environment at the same time can promote a more rapid establishment of social stability. If possible, once weaning groups are selected and placed in the nursery, keep these groups together to harvest. Any change in the pig group will again result in the need for a new hierarchy to be established, along with fighting and disrupting the group. “Allow newly selected nursery groups to establish their hierarchy by avoiding interventions during the first 48 hours, except to treat sick or injured pigs”, recommends Dr. Parke. “A well-enriched environment, such as chewable ropes and toys, can help reduce stress levels and may reduce the frequency of abnormal behaviors such as tail biting and aggression.”
Environmental management
The piglets should be kept at an optimal temperature between 27-30°C – depending on floor type, weight, and age of piglets. Adding a heat lamp/floor mat warm area for just-weaned piglets will further assist thermoregulation and minimize stress through the weaning transition.
Proper ventilation is crucial for maintaining air quality and preventing the buildup of harmful gases like ammonia. Good airflow helps regulate temperature and humidity, reducing stress on the pigs. However, care must be taken to avoid drafts that can chill young pigs. For example, a draft of 0.5 m/second can ‘feel’ like an 8°C drop for the piglet.
Targets for gas, dust, and bacteria levels
Risk factor
Gas
Total dust
Respirable dust
Bacteria
Ammonia
Hydrogen sulphide
Carbon dioxide
Target levels
<10ppm (20ppm max.)
<5ppm
<3,000ppm (aim for <1,500ppm)
2.4mg/m3
0.23mg/m3
100,000 CFU/m3
Flooring and pen materials should be robust, in good condition, and easily cleaned to reduce the risk of skin abrasions and subsequent infections.
Provide sufficient space (recommended 0.19 m2/8 kg pig on slat/solid floor) in pens to minimize competition for feed and water and to reduce social stress among piglets.
Weaner pigs benefit from using the same type of feeder in the nursery as in the farrowing room. This consistency can help to reduce stress and anxiety during the transition to the nursery and increase the feed intake during the first few days post-weaning.
Nutritional support
Weaning stress and poor feed intake post-weaning commonly result in dysbiosis and a decrease in villus height in the small intestine of piglets. Associated digestive impairment and altered gut morphology can lead to decreased nutrient absorption, as well as enteric and systemic health issues. A palatable transition diet, from 7 days pre- to 7 days post-weaning, is recommended to keep piglets eating. The composition or form of the transition diet should remain the same during this period. Consider using functional feed additives, such as phytomolecules or egg immunoglobulins, to support microbial modulation and gut integrity.
Ensure piglets have access to fresh, cool, and clean water (minimum water flow of 0.5-0.7L/minute), with enough drinking space (maximum of ten piglets per drinker). Consider providing additional water supply points (e.g., bowls) in the first week.
Hygiene and biosecurity
All-in, all-out management avoids the mixing of different age groups. It is particularly beneficial for weaner pigs, as it helps minimize disease transmission. After removing each batch of weaners, the nursery must be thoroughly cleaned, disinfected, and dried. This includes not just the floors but also feeders, waterers, and any equipment used in the room.
There should be strict rules for everything that comes through the external perimeter fence. Internal biosecurity is also essential, e.g., changing into clean, disinfected boots and thoroughly washing hands when moving between rooms/buildings.
Routine monitoring
Regular and proactive monitoring of weaner pigs, including carefully observing their behavior, is essential for ensuring their health and optimizing growth performance. By implementing effective monitoring strategies, producers can identify potential challenges early and take timely interventions to minimize negative impacts.
Pig positive people
Dr. Parke emphasized that the attitude and skills of stockpersons play a significant role in reducing stress during this vulnerable weaning transition period. Positive handling can improve piglet welfare and their future response to human contact, which is crucial for their short and long-term production performance.
Piglets that receive positive handling are likelier to demonstrate affiliative behaviors towards humans, facilitating smoother transitions during weaning and enhancing their overall development. Stockpersons should be trained to recognize signs of stress or discomfort in pigs.
Collaborative approach
“Collaboration is critical for successful weaning; we can’t have silos in pig production unless it’s to store feed,” joked Dr. Parke. “By adopting a proactive approach that emphasizes collaboration and comprehensive management strategies across the production system, pig welfare and long-term productivity of the herd will be enhanced,” she concluded.
EW Nutrition’s Swine Academy took place in Ho Chi Minh City and Bangkok in October 2024. Dr. Merideth Parke, Global Application Manager, Swine, was one of the highly experienced speakers of EW Nutrition. She is a veterinarian who strongly focuses on swine health and preventive medicine.
Piglet rearing – there is still room for improvement!
By I. Heinzl, Editor, and Predrag Persak, Regional Technical Manager North Europe
Optimal rearing conditions for piglets are crucial for ensuring their healthy growth, reducing mortality, and enhancing productivity. These conditions include proper temperature, nutrition, housing, hygiene, and care. Here are the key aspects:
1. Temperature and ventilation
Piglets are sensitive to cold because they cannot regulate their body temperature effectively in the first few days after birth. Proper temperature control is essential to prevent chilling, possibly leading to illness and death. Additionally, regulating the temperature would cost energy, which otherwise could be spent for growth.
Signs of a too-cold environmental temperature are piling on top of one another, tucking the legs under the body, being unable to get up, laying near a corner or wall, or shivering, which may stop if the conditions worsen. Measuring the body temperature shows less than 35°C in the case of chilling.
The following temperatures are recommended for successful piglet rearing:
Farrowing unit (for newborns)
32 – 35°C (90–95°F) during the first few days
After the first week
The temperature can gradually decrease by about 1.5-2.0°C per week until it reaches 25°C (77°F)
For supplemental heating, heat lamps, heated floors, or creep areas (a designated warm spot) can be used to maintain the ideal temperature, especially in cooler climates.
Temperature is often closely related to ventilation. Ventilation is essential to reduce dust, humidity, ammonia, and other harmful substances occurring in the air. However, if fresh/cold air enters the pigsty, the temperature decreases, which can get dangerous for the piglets. Suitable ventilation means finding a good balance between providing fresh air and maintaining temperature to prevent energy losses and chilling of the piglets.
Comfort zones can be a solution. They are an effective way to keep the piglets warm and ventilation rates where needed to maintain proper air exchange and humidity levels.
2. Nutrition
Nutrition is critical for piglet growth and immune system development. Most important after birth is the access to colostrum. Piglets are born with an immature immune system, and the maternal antibodies ingested with the colostrum are vital for their survival. They should consume colostrum within the first 6 hours after birth.
It will take 5 to 7 days for piglets to stabilize and get regular on suckling schedule.
At around seven days of age, it is recommended to introduce a highly digestible, nutrient-dense creep feed that helps transition piglets from milk to solid food. Fresh and clean water of the best quality must always be available.
Never forget most important nutrient, beside sow´s love and care – water. Allow piglets free access to the excellent quality water.
3. Housing and Space
A well-designed, clean, and dry environment is critical for reducing stress and promoting health. Farrowing crates help prevent sows from accidentally crushing the piglets during the first few weeks. However, these farrowing crates should provide enough space for the sow to nurse the piglets while allowing piglets to move freely.
Separate warm and clean areas (creep spaces) for the piglets within the farrowing pen are helpful to help the piglets escape from cooler or potentially dangerous parts of the crate. Straw, sawdust, or rubber mats should be provided to keep the piglets warm and comfortable, and good drainage is essential to maintain dryness.
4. Hygiene and Health
Hygiene is crucial to prevent disease and promote the health of piglets. For this purpose, pens and farrowing units should be thoroughly cleaned. Regular removal of waste and keeping bedding dry helps control pathogens. It is essential to clean and disinfect the farrowing unit from one farrowing to the other to reduce disease risks.
Health: After birth, the piglets’ umbilical cord stump should be disinfected to prevent infections. A further essential precautionary measure to prevent anemia is an oral supplementation or an iron injection within the first three days of life, as piglets are born with low iron levels.
For further health monitoring and management, it should be ensured that the piglets are vaccinated against common diseases, such as E. coli, Mycoplasma, and Porcine Circovirus. Additionally, deworming protocols and monitoring for signs of parasites should be implemented for parasite control.
5. Weaning Practices
Piglets are typically weaned between 3 and 4 weeks of age, but early weaning (around 21 days) can be practiced in intensive systems. Optimal weaning requires gradual adaptation to solid feed and a stress-free environment.
If the piglets are weaned at 21 to 28 days, a high-quality starter diet after weaning is essential to maintain growth rates and minimize post-weaning stress.
6. Minimizing Stress
Stress management is essential to prevent disease and poor growth. For this purpose, minimize handling to the minimum during the first few days and, if necessary, handle the piglets gently to reduce stress.
A new environment also means strain for the piglets, so keep the litter groups together during weaning to reduce fighting and social stress.
7. Supportive functional feed ingredients
Depending on veterinary and managing practices, the availability of feed, and the possible use of antimicrobials or other medicals as prophylactics, there can be high variability in rearing conditions in diverse areas of the world. In the following, two functional feed ingredients with entirely different modes of action are presented that support piglets at different rearing conditions.
7.1 Egg immunoglobulins (IgY) support piglets under poor rearing conditions
Egg immunoglobulins are beneficial if piglets are not raised under the best conditions, meaning lower hygienic standards and higher pathogenic pressure. With egg immunoglobulins coming from hens having been in contact with pathogens relevant to piglets, it is possible to support the young animals. What is the background? Hens are able to transfer maternal antibodies against diseases that they are confronted with to the egg. With this mechanism, they can provide their progeny with a starter kit for the first time after hatching. However, the best thing is that these antibodies are also helpful for mammals.
A trial conducted on a commercial farm in Spain shows the weight development of piglets fed an IgY-containing egg powder product (EP) compared to a negative control. The weaned piglets were fed a two-phase feeding (15 days prestarter, 22 days starter). The control (n=51) received no additional functional feed ingredient, whereas the EP group was fed 2 kg of the product/t of feed during the prestarter phase. The animals were weighed individually on days 16 and 37.
The results are shown in Figures 1 and 2.
Figure 1: Weight development of piglets receiving an IgY-containing egg powder product compared to a negative control
Figure 2: Daily gain of piglets receiving an IgY-containing egg powder product compared to a negative control
Explanation of the results: Under poor hygienic conditions, the pathogenic pressure is relatively high, and everything lowering this pressure helps to improve gut health, the utilization of nutrients, and performance. Egg immunoglobulins positively influence the gut microbiome, thus helping reduce diarrhea. By lowering the pathogenic pressure, the organism’s energy can be used for growth and must not be employed for the body’s defense.
7.2 Phytomolecules can even show improvement under optimum conditions
Phytomolecules generally show diverse gut health-promoting effects, from driving the intestinal microbiome in the right direction and strengthening the intestinal barrier to acting as antioxidants or anti-inflammatories or increasing the secretion of digestive juices and, therefore, improving digestion. Which mode of action is relevant if the piglets are raised under already optimal conditions (best hygiene, no prophylactic antibiotics or zinc oxide) and show the highest growth? Is there still room for improvement? Yes, it is. A trial conducted in Germany adduces evidence.
In this trial, 220 piglets weaned on average at 26 days and weighing around 8 kg were housed in 20 pens of 11 castrated males or gilts each. Piglets were blocked by body weight and fed a two-phase feeding program (phase 1 from day 1 to day 13 and phase 2 from day 17 to day 40; pelleted diet). Neither feed or water medication nor therapeutic levels of ZnO were used.
The results of this piglet trial can be seen in Figures 3 and 4.
Figure 3: Weight development of piglets fed Ventar D compared to a negative control
Figure 4: Feed conversion rate in piglets fed Ventar D compared to a negative control
Explanation of the results: The figures show that the piglets in the control already have an extremely high weight compared to those of a similar age in the previous trial, indicating the best rearing conditions in this trial. But, even here, Ventar D has the capacity to improve performance. Why? High-performing animals stress their body more than low-performing ones. Anabolic processes increase oxidative stress and non-infectious inflammation and burden the immune system. The relevant mode of action of Ventar D is not the gut health-promoting or the antimicrobial one because there is no issue. The relevant modes of action in this case are antioxidant and anti-inflammatory. With these two characteristics, Ventar D still has the capacity to improve the performance of piglets that are already at the top level.
8. Conclusion
For high piglet performance, providing the best possible rearing conditions is essential. However, there are differences concerning these conditions in different areas of the world. Depending on them, different feed strategies can be used. Egg immunoglobulins show the best effects if there is a certain pathogenic pressure. Phytomolecules, however, due to their various modes of action, can be beneficial under different levels in rearing conditions. In a low standard, the antimicrobial and gut health-promoting effect is more relevant; in the case of best conditions, the anti-oxidant and anti-inflammatory effects are decisive.
In summary, it could be said that functional feed ingredients have significant advantages in piglet rearing, but the right choice must be made depending on the prevailing conditions.
The crucial role of short-chain fatty acids and how phytomolecules influence them
by Dr. Inge Heinzl, Editor EW Nutrition
For optimum health, the content of short-chain fatty acids (SCFAs) is decisive. On the one hand, they act locally in the gut, on the other hand, they are absorbed via the intestinal mucosa into the organism and can affect the whole body. Newer studies in humans show a connection between the deficiency of SCFAs and the occurrence of chronic diseases such as diabetes type 2 or chronic inflammatory gut diseases.
SCFAs – what are they, and where do they come from?
SCFAs consist of a chain of one to six carbon atoms. They are crucial metabolites primarily generated through the bacterial fermentation of dietary fiber (DF) in the hindgut. However, SCFAs and branched SCFAs can also arise during protein fermentation. Short-chain fatty acids predominantly include acetate, propionate, and butyrate, which together account for over 95% of the total SCFAs, typically in a 60:20:20 ratio.
Acetate is produced in two different ways, via the acetyl-CoA and the Wood-Ljungdahl pathways where Bacteroides spp., Bifidobacterium spp., Ruminococcus spp., Blautia hydrogenotrophica, Clostridium spp. are involved. Additionally, acetogenic bacteria can synthesize acetate from carbon dioxide and formate through the Wood-Ljungdahl pathway (Ragsdale and Pierce, 2021). Acetate counts for more than 50% of the total SCFAs in the colon and is the most abundant one.
Propionate can also be produced in two ways. If it is produced via the succinate pathway involving the decarboxylation of methyl malonyl-CoA, the essential bacteria are Firmicutes and Bacteroides. In the acrylate pathway, lactate is converted to propionate. Here, only some bacteria, such as Veillonellaceae or Lachnospiraceae, participate.
Butyrate is produced from acetyl-CoA via the classical pathway by several Firmicutes. However, also other gut microbiota such as Actinobacteria, Proteobacteria, and Thermotogae, which contain essential enzymes (e.g., butyryl coenzyme A dehydrogenase, butyryl-CoA transferase, and butyrate kinase) can be involved. Butyrate can also be produced via the lysine pathway from proteins.
Besides the production of SCFAs from dietary fiber, there is another possibility for the synthesis of SCFAs as well as branched SCFAs – the fermentation of protein in the hindgut. This is something we want to avoid, since it´s clear signal of incorrect animal nutrition. It tells us that there is either oversupply of protein or decrease in protein digestion and absorption.
Which roles do SCFAs play?
SCFAs play a crucial role in the maintenance of gut health. Some benefits originate from these substances’ general character, while others are specific to one acid. If we talk about the benefits of all SCFAs, we can mention the following:
Primarily, SCFAs are absorbed by the intestine and serve enterocytes as an essential substrate for energy production.
By lowering the pH in the intestine, SCFAs inhibit the invasion and colonization of pathogens.
SCFAs can cross bacterial membranes in their undissociated form. Inside the bacterial cell, they dissociate, resulting in a higher anion concentration and bactericidal effect (Van der Wielen et al., 2000)
SCFAs repair the intestinal mucosa
They mitigate intestinal inflammation by G protein-coupled receptors (GPRs).
They enhance immune response by producing cytokines such as IL-2, IL-6, IL-10, and TNF-α in the immune cells. Furthermore, they enhance the differentiation of T-cells into T regulatory cells (Tregs) and bind to receptors (Toll-like receptor, G protein-coupled receptors) on immune cells (Liu et al., 2021).
SCFAs are involved in the modulation of some processes in the gastrointestinal tract, such as electrolyte and water absorption (Vinolo et al., 2011)
After seeing the general characteristics of short-chain fatty acids, let us take a closer look at the specialties of the single SCFAs.
Acetate might play a crucial role in the competitive process between enteropathogens and bifidobacteria and help to build a balanced gut microbial environment (Liu et al., 2021). Additionally, acetate promotes lipogenesis in adipocytes (Liu et al., 2022).
Concerning general health, acetate inhibits, e.g., lung inflammatory response and the reduced air-blood permeability induced by avian pathogenic E. coli-caused chicken colibacillosis (Peng et al., 2021).
Propionate is thought to be involved in controlling intestinal inflammation by regulating the immune cells assisting and, consequently, in maintaining the gut barrier. Furthermore, propionate regulates appetite, controls blood glucose, and inhibits fat deposition in broiler chickens (Li et al., 2021).
In a trial conducted by Elsherif et al. (2022), birds fed a diet with 1.5 g sodium propionate/kg showed considerably (P<0.05) longer and wider guts, higher counts of lactobacillus(P<0.05) and no colonization of Clostridium perfringens. The immunological state improved significantly (P<0.05), which could be seen by the higher antibody titers when the birds were vaccinated against Newcastle disease or avian influenza.
Butyrate additionally improves the function of the intestinal barrier by regulating the assembly of tight junctions (Peng et al., 2009) and stimulating cell renewal and differentiation of the enterocytes. Butyrate-producing microbes on their side prevent the dysbiotic expansion of potentially pathogenic E. coli and Salmonella (Byndloss et al., 2017; Cevallos et al., 2021) by stimulating PPAR-γ signaling. This leads to the suppression of iNOS synthesis and a significant reduction of iNOS and nitrate in the colonic lumen. Furthermore, the microbiota-induced PPAR-γ-signaling inhibits dysbiotic Enterobacteriaceae expansion by limiting the bioavailability of oxygen and, therefore, respiratory electron acceptors to Enterobacteriaceae in the colon.
In a trial conducted by Xiao et al. (2023), sodium butyrate enhanced broiler breeders’ reproductive performance and egg quality due to the regulation of the maternal intestinal barrier and gut microbiota. Additionally, it improved the antioxidant capacity and immune function of the breeder hens and their offspring.
SCFAs’ production can be managed
The extent of production depends on the diet and the composition of the intestinal flora. Nutritional strategies can be taken to regulate the production of short-chain fatty acids by providing dietary fiber and prebiotics, the respective bacteria but also additives in the diet or, on the other, negative way, use of antibiotics.
One example of SCFA-promoting additives is phytomolecules. Ventar D, a blend of diverse gut health-promoting phytomolecules, shows its SCFAs-increasing effect in a trial with Ross 308 broilers.
Trial design: The 41-day research study was conducted at an R&D farm in Turkey, with 3200 Ross 308 broilers in total. The day-old broiler chicks were randomly divided into two groups with 8 replicates in 16-floor pens (6.5×2 m each), each of 200 chicks (100 males and 100 females). One group was managed as a control group with regular feed formulation, and the other group was supplemented with Ventar D. All the birds were provided feeds and water ad libitum. Temperature, lighting, and ventilation were managed as per Ross 308 recommendation.
Groups
Application dose
Starter (crumbles)
Grower & Finisher – 1 & 2 (pellet)
Control
No additive
Ventar D
100 gm/MT
100 gm/MT
All the birds and feed were weighed on days 0, 11, 23, and 41. Dead birds were also weighed, and the feed consumption was corrected accordingly. At the end of the experiment, one male and one female chicken close to the average weight of each pen were separated, weighed, and slaughtered. Short-chain fatty acid (SCFA) concentration in the caecum was measured by gas chromatography (Zhang et al. 2003). Statistical analysis of the data obtained in this study was carried out in the Minitab 18 program using the T-test following the randomized block trial design (P ≤ 0.05). The research results were subjected to statistical analysis on a pen basis. Mortality results were evaluated with the Chi-square test.
Results: Ventar D significantly increased the levels of acetate, butyrate, and total SCFAs. The level of propionate was numerically higher. Additionally, higher final body weights (on average 160 g), improved feed efficiency (6 points), a higher EPEF (33 points), and lower mortality (0.5%) could be asserted in this experiment.
One explanation could be the microbiota-balancing effect of Ventar D. Meimandipour et al. (2010), for example, saw in their study that increased colonization of Lactobacillus salivarius and Lactobacillus agilis in cecum significantly increased propionate and butyrate formation in caeca.
Phytomolecules: Balancing intestinal microbiome and increasing healthy SCFAs
By promoting beneficial intestinal bacteria and fighting the harmful ones, phytomolecules drive the microbiome in the right direction and promote the production of short-chain fatty acids. Their gut health-protecting effect, in turn, provides for adequate digestion and absorption of nutrients, leading to optimal feed conversion and growth rates. The support of the immune system and the promotion of the antioxidant capacity additionally enhance the health of the animals. Healthy animals grow better, which ultimately leads to a higher profit for the farm.
References:
Byndloss, Mariana X., Erin E. Olsan, Fabian Rivera-Chávez, Connor R. Tiffany, Stephanie A. Cevallos, Kristen L. Lokken, Teresa P. Torres, et al. “Microbiota-Activated PPAR-γ Signaling Inhibits Dysbiotic Enterobacteriaceae Expansion.” Science 357, no. 6351 (August 11, 2017): 570–75. https://doi.org/10.1126/science.aam9949.
Cevallos, Stephanie A., Jee-Yon Lee, Eric M. Velazquez, Nora J. Foegeding, Catherine D. Shelton, Connor R. Tiffany, Beau H. Parry, et al. “5-Aminosalicylic Acid Ameliorates Colitis and Checks Dysbiotic Escherichia Coli Expansion by Activating PPAR-γ Signaling in the Intestinal Epithelium.” mBio 12, no. 1 (February 23, 2021). https://doi.org/10.1128/mbio.03227-20.
Elsherif, Hany M.R., Ahmed Orabi, Hussein M.A. Hassan, and Ahmed Samy. “Sodium Formate, Acetate, and Propionate as Effective Feed Additives in Broiler Diets to Enhance Productive Performance, Blood Biochemical, Immunological Status, and Gut Integrity.” Advances in Animal and Veterinary Sciences 10, no. 6 (June 2022): 1414–22.
Li, Haifang, Liqin Zhao, Shuang Liu, Zhihao Zhang, Xiaojuan Wang, and Hai Lin. “Propionate Inhibits Fat Deposition via Affecting Feed Intake and Modulating Gut Microbiota in Broilers.” Poultry Science 100, no. 1 (January 2021): 235–45. https://doi.org/10.1016/j.psj.2020.10.009.
Liu, Lixuan, Qingqing Li, Yajin Yang, and Aiwei Guo. “Biological Function of Short-Chain Fatty Acids and Its Regulation on Intestinal Health of Poultry.” Frontiers in Veterinary Science 8 (October 18, 2021). https://doi.org/10.3389/fvets.2021.736739.
Liu, Lixuan, Qingqing Li, Yajin Yang, and Aiwei Guo. “Biological Function of Short-Chain Fatty Acids and Its Regulation on Intestinal Health of Poultry.” Frontiers in Veterinary Science 8 (October 18, 2021). https://doi.org/10.3389/fvets.2021.736739.
Meimandipour, A., M. Shuhaimi, A.F. Soleimani, K. Azhar, M. Hair-Bejo, B.M. Kabeir, A. Javanmard, O. Muhammad Anas, and A.M. Yazid. “Selected Microbial Groups and Short-Chain Fatty Acids Profile in a Simulated Chicken Cecum Supplemented with Two Strains of Lactobacillus.” Poultry Science 89, no. 3 (March 2010): 470–76. https://doi.org/10.3382/ps.2009-00495.
Peng, Lu-Yuan, Hai-Tao Shi, Zi-Xuan Gong, Peng-Fei Yi, Bo Tang, Hai-Qing Shen, and Ben-Dong Fu. “Protective Effects of Gut Microbiota and Gut Microbiota-Derived Acetate on Chicken Colibacillosis Induced by Avian Pathogenic Escherichia Coli.” Veterinary Microbiology 261 (October 2021): 109187. https://doi.org/10.1016/j.vetmic.2021.109187.
Peng, Luying, Zhong-Rong Li, Robert S. Green, Ian R. Holzmanr, and Jing Lin. “Butyrate Enhances the Intestinal Barrier by Facilitating Tight Junction Assembly via Activation of AMP-Activated Protein Kinase in Caco-2 Cell Monolayers.” The Journal of Nutrition 139, no. 9 (September 2009): 1619–25. https://doi.org/10.3945/jn.109.104638.
Ragsdale, Stephen W., and Elizabeth Pierce. “Acetogenesis and the Wood–Ljungdahl Pathway of CO2 Fixation.” Biochimica et Biophysica Acta (BBA) – Proteins and Proteomics 1784, no. 12 (December 2008): 1873–98. https://doi.org/10.1016/j.bbapap.2008.08.012.
Vinolo, Marco A.R., Hosana G. Rodrigues, Renato T. Nachbar, and Rui Curi. “Regulation of Inflammation by Short Chain Fatty Acids.” Nutrients 3, no. 10 (October 14, 2011): 858–76. https://doi.org/10.3390/nu3100858.
Wielen, Paul W. van der, Steef Biesterveld, Servé Notermans, Harm Hofstra, Bert A. Urlings, and Frans van Knapen. “Role of Volatile Fatty Acids in Development of the Cecal Microflora in Broiler Chickens during Growth.” Applied and Environmental Microbiology 66, no. 6 (June 2000): 2536–40. https://doi.org/10.1128/aem.66.6.2536-2540.2000.
Xiao, Chuanpi, Li Zhang, Bo Zhang, Linglian Kong, Xue Pan, Tim Goossens, and Zhigang Song. “Dietary Sodium Butyrate Improves Female Broiler Breeder Performance and Offspring Immune Function by Enhancing Maternal Intestinal Barrier and Microbiota.” Poultry Science 102, no. 6 (June 2023): 102658. https://doi.org/10.1016/j.psj.2023.102658.
Oxidative & Inflammatory stress in reproductive Sows
By Dr. Inge Heinzl, Editor and Technical Team, EW Nutrition
One of the biggest challenges in swine production is keeping the modern, hyperprolific sow healthy and in good shape so that she can wean large, healthy litters and maintain her high reproductive performance.
Unfortunately, sows often suffer from stress and increased systemic inflammation around farrowing and during lactation. This leads to impaired feed intake and disturbed endocrine homeostasis, negatively affecting reproductive and litter performance.
The key to increasing the efficiency of pig production is to reduce the metabolic burden of sows while maintaining the reproductive performance of high-yield sows. A deep understanding of the complex interplay between environmental factors, sow well-being, health, and productivity is necessary to implement enhanced nutritional regimens and meticulous management practices.
Why does oxidative stress occur in today’s sows?
Nowadays, hyperprolific sows produce between 30 and 40 weaned piglets per year and are at a higher risk of suffering from stress. What are the reasons?
A high number of piglets causes oxidative stress
Oxidative stress occurs when reactive oxygen species (ROS) are produced faster than the body’s antioxidant mechanisms can neutralize them and cause damage to lipids, proteins, and DNA. During gestation, the sow needs high amounts of energy to provide for the fetuses. This energy is produced in the placental mitochondria. The placenta, therefore, is a place of active oxygen metabolism during gestation and a source of oxidative stress. In hyperprolific sows, a higher number of fetuses need even more energy to grow. Consequently, ROS production and the risk for intrauterine growth retardation (IUGR) increases (Figure 1). Moreover, evidence shows that the body’s antioxidant potential is reduced in late gestation and after parturition (Szczubial, 2010), resulting in increased oxidative stress biomarkers (Yang, 2023). Increased milk production for large litters demands a substantial amount of energy, risking similar oxidative distress. Therefore, both the final phase of gestation and the subsequent lactation period are predestined for oxidative stress, which has been demonstrated by reduced TEAC (Trolox equivalent antioxidant capacity) levels during these phases (Lee et al., 2023).
Figure 1. Illustration of the effect of oxidative stress on the fetus: intrauterine growth retardation (IUGR) (adapted from Yang et al., 2023)
Heat and ambient stress also contribute
The reproductive sow produces lots of heat. From the beginning of gestation, the sow’s thermoneutral zone decreases. This, however, does not always correspond with the ambient conditions. Especially during the last days of gestation, the discrepancy is exceptionally high as everything is prepared for the newborn piglets, which need a temperature of about 27-35°C. The sow, on the contrary, would be happy with 18-22°C. Additionally, changes around farrowing – moving to the farrowing unit, social stress, change of feed, and the preparation for parturition – exert additional stress for the sows.
Why does the inflammation level increase?
After parturition, systemic inflammation is a normal phenomenon: the reproductive organs have sustained injuries during the parturition process and require remodeling. Inflammation is a natural and desired process, to repair the tissues and return to a normal status. However, inflammation is increased in modern sows, adversely affecting their inflammatory balance. Some possible underlying reasons are:
The high numbers of piglets need a lot of space in the uterus, often leading to damage of the uterine tissue and an inflammatory response in the sows. Lee et al. (2023) found significantly (p<0.10) higher TNF-α concentrations in sows with litters of 15-20 piglets than in sows with 7-14 piglets. TNF-α is a biomarker of inflammation.
Pathogenic infections – particularly infections of the reproductive tract – can induce a prolonged or excessive inflammatory state. A further reason can be the need for more obstetric interventions in hyperprolific sows, which can injure the birth canal or the uterus.
Imbalanced nutrition: Excessive backfat is associated with a higher expression of proinflammatory cytokines, and feed contaminated with mycotoxins can impair the sow’s immunocompetence.
Biomarkers can inform us about the oxidative status
Biomarkers are naturally occurring molecules that help us identify diseases or physiological processes. They provide insights into the oxidative state and inflammatory processes.
Anti-oxidative biomarkers
To check the anti-oxidative capacity, the “beneficial” substances, or antioxidants, can be quantified. These substances can neutralize free radicals or be neutralized by them. Higher levels of antioxidants indicate better antioxidant capacity; when antioxidants are abundant, fewer oxidizable substances have undergone oxidation.
Examples of antioxidant biomarkers:
Total Antioxidant Capacity (T-AOC): represents the synergistic interaction effects of all antioxidants in a matrix (E.g., diet or body fluids). It’s a global measure of non-enzymatic antioxidant efficiency. Various assays, like Trolox Equivalent Antioxidant Capacity (TEAC), which measures a substance’s antioxidant capacity compared to Trolox, can measure T-AOC.
Glutathione Peroxidase (GSH-Px) belongs to the peroxidase family and converts hydrogen peroxide to water.
Catalase (CAT): scavenges ROS. Its activity can predict oxidative stress.
Superoxide Dismutase (SOD): catalyzes the dismutation of superoxide radicals to oxygen and hydrogen peroxide.
Oxidative biomarkers
Oxidative stress biomarkers, the ‘negative’ substances, can also serve as general biomarkers. These include free radicals with oxidant capacity or intermediate/final oxidation products. Ideally, their levels should be minimized.
Examples of oxidative stress biomarkers:
Thiobarbituric acid reactive substances (TBARS): to measure lipid peroxidation products in cells, tissues, and body fluids.
Reactive oxygen species(ROS) or free radicals: unstable, oxygen-containing molecules that react with other molecules in a cell. They might damage DNA, RNA, and proteins and cause cell death. Hydrogen Peroxide (H₂O₂) is a ROS produced during normal cellular metabolism, which causes oxidative damage at excessive levels.
Malondialdehyde (MDA): a final product of oxidative fat degradation and, therefore, a biomarker for lipid peroxidation.
Pro-inflammatory biomarkers
Like oxidative stress, the interplay between pro- and anti-inflammatory signals helps develop the proper immune response for the appropriate duration.
Examples of Pro-inflammatory biomarkers or molecules produced in the case of inflammation:
Plasma Adenosine Deaminase (ADA-1 and ADA-2): involved in immune regulation, with ADA-1 inhibiting pro-inflammatory responses and ADA-2 supporting immune cell functions.
Interleukins (IL-1α and IL-1β), IL-6: IL-1α and IL-1β are associated with inflammatory diseases, IL-6: is produced during inflammation and acute-phase response.
Tumor Necrosis Factor α (TNF-α): endogenous pyrogen that induces fever and promotes inflammation.
C-reactive Protein (CRP): liver-produced acute-phase protein responding to inflammation.
Procalcitonin (PCT) is produced by the liver during infections and helps detect bacterial infections.
Examples of anti-inflammatory substances – the “good ones”:
Interleukines – IL-4, IL-10: inhibit the function of the macrophages and act, therefore, anti-inflammatory
Cortisol: anti-inflammatory and immune-suppressive
ACTH: stimulates the production and release of cortisol
Higher stress or infection level lowers performance in sows and piglets
As mentioned, hyperprolific sows suffer from higher oxidative stress, especially during late gestation, parturition, and lactation. Additionally, systemic inflammation occurs to repair the injured tissues to facilitate the healing of the birth canal and remodeling of the uterus to establish the subsequent pregnancy. To this purpose, an inflammatory cascade, triggered by the injuries due to gestation and parturition, involves the release of critical (pro-inflammatory) mediators such as TNF-α and IL-6, leading to the activation of acute phase proteins.
After triggering inflammatory pathways, anti-inflammatory pathways must also be activated to reestablish homeostasis in the reproductive organs (Serhan & Chiang, 2008). Alterations at the onset of anti-inflammatory pathways and exacerbated activation and maintenance of inflammatory pathways can lead to uncontrolled inflammation and the onset of reproductive disease in sows (Kaiser et al., 2018), as well as reduced feed intake and insufficient milk production, resulting in poorly growing piglets and lower weaning weights or piglets suffering from clinical infectious diseases such as diarrhea. If possibly homeostasis cannot be restored, the sow is at risk of contracting diseases like post-partum dysgalactia syndrome (PPDS), lameness, and impaired fertility.
Targeted use of polyphenols can mitigate inflammation and improve the oxidative status of sows
There are several experiments showing the beneficial effects of natural compounds. Especially polyphenols, disposing of phenyl rings and two or more hydroxyl substituents, are perfect radical scavengers and proven antioxidants (Chen, 2023). Phytogenic substances that have anti-inflammatory effects can be found in the families of polyphenols as well as terpenoids, flavonoids, saponins, and tannins (Bunte et al., 2019; Ge et al., 2022; Ginwala et al., 2019; Santos Passos et al., 2022; Ambreen and Mirza, 2020).
Here are some examples showing the beneficial effects of phytochemicals:
Primiparous sows fed with Moringa oleifera leaf meal, rich in polyphenols, saponins, and tannins, illustrate the potential of phytomolecules: serum levels of T-AOC (total anti-oxidative capacity), were increased in late gestation and during lactation, while MDA was reduced. Additionally, piglets that received Moringa oleifera meal showed the highest serum CAT and SOD activities. The meal significantly decreased the farrowing length and number of stillbirths, while there was an increasing trend in the number of live‐born piglets (Sun et al., 2020).
The polyphenol Daidzein, a member of the class of compounds known as isoflavones (200 mg/kg during gestation), increased the total antioxidant capacity (T-AOC) and the activities of glutathione peroxidase and superoxide dismutase. Additionally, it elevated the level of immunoglobulin G and increased the number of piglets born and born alive per litter (Li et al., 2021).
Glycitein, a polyphenol occurring in the isoflavone fraction of soy products, applied during late gestation and lactation increased the total antioxidant capacity and SOD activity during the first 18 days of lactation and the CAT and GSH-Px activity in mid-lactation. Plasma MDA level was reduced from late gestation to the 18th day of lactation. The enhanced oxidative status of the sow resulted in a higher daily gain of the piglets and a higher weaning weight of the litter (Hu et al., 2015).
Meng et al. (2018) tested Resveratrol (300 mg/kg), a stilbenes polyphenol, in sows from day 20 of gestation until farrowing. They saw noticeably higher GSH-Px, SOD, and CAT activities, as well as lower contents of MDA and H2O2 in the placental tissue, improving the antioxidant status of sows and piglets.
Xu et al. (2022) fed silymarin to sows in late gestation. They observed that IL-1ß concentration in the blood sample on the 18th day of lactation was reduced in the supplemented group. The altered fecal microbiota was associated with variations in inflammatory factors, suggesting that silymarin modulates microbiota in the gut and may improve the health of lactation sow.
Phytochemicals support sows against oxidative and inflammatory stress
The above-presented examples show that phytochemicals, particularly those developed to have a potent anti-inflammatory and anti-oxidative capacity, have a high potential to alleviate oxidative stress in pregnant and lactating sows and reduce inflammation when applied in sow diets. Consequently, a broader use of these natural substances should be considered to reduce the metabolic burden of sows and increase the efficiency of pig production.
References:
Ambreen, Madieha, and Safdar Ali Mirza. “Evaluation of Anti-Inflammatory and Wound Healing Potential of Tannins Isolated from Leaf Callus Cultures of Achyranthes Aspera and Ocimum Basilicum.” Pak J Pharm Sci . 33, no. 1 (January 2020): 361–69.
Bunte, Kübra, Andreas Hensel, and Thomas Beikler. “Polyphenols in the Prevention and Treatment of Periodontal Disease: A Systematic Review of in Vivo, Ex Vivo and in Vitro Studies.” Fitoterapia 132 (January 2019): 30–39. https://doi.org/10.1016/j.fitote.2018.11.012.
Chen, Jun, Zhouyin Huang, Xuehai Cao, Tiande Zou, Jinming You, and Wutai Guan. “Plant-Derived Polyphenols in Sow Nutrition: An Update.” Animal Nutrition 12 (March 2023): 96–107. https://doi.org/10.1016/j.aninu.2022.08.015.
Ge, Jiamin, Zhen Liu, Zhichao Zhong, Liwei Wang, Xiaotao Zhuo, Junjie Li, Xiaoying Jiang, Xiang-Yang Ye, Tian Xie, and Renren Bai. “Natural Terpenoids with Anti-Inflammatory Activities: Potential Leads for Anti-Inflammatory Drug Discovery.” Bioorganic Chemistry 124 (July 2022): 105817. https://doi.org/10.1016/j.bioorg.2022.105817.
Ginwala, Rashida, Raina Bhavsar, De Gaulle Chigbu, Pooja Jain, and Zafar K. Khan. “Potential Role of Flavonoids in Treating Chronic Inflammatory Diseases with a Special Focus on the Anti-Inflammatory Activity of Apigenin.” Antioxidants 8, no. 2 (February 5, 2019): 35. https://doi.org/10.3390/antiox8020035.
Hu, Y. J., K. G. Gao, C. T. Zheng, Z. J. Wu, X. F. Yang, L. Wang, X. Y. Ma, A. G. Zhou, and Z. J. Jiang. “Effect of Dietary Supplementation with Glycitein during Late Pregnancy and Lactation on Antioxidative Indices and Performance of Primiparous Sows1.” Journal of Animal Science 93, no. 5 (May 1, 2015): 2246–54. https://doi.org/10.2527/jas.2014-7767.
Kaiser, Marianne, Stine Jacobsen, Pia Haubro Andersen, Poul Bækbo, José Joaquin Cerón, Jan Dahl, Damián Escribano, Peter Kappel Theil, and Magdalena Jacobson. “Hormonal and Metabolic Indicators before and after Farrowing in Sows affected with postpartum Dysgalactia Syndrome.” BMC Veterinary Research 14, no. 1 (November 7, 2018). https://doi.org/10.1186/s12917-018-1649-z.
Lee, Juho, Hyeonwook Shin, Janghee Jo, Geonil Lee, and Jinhyeon Yun. “Large Litter Size Increases Oxidative Stress and Adversely Affects Nest-Building Behavior and Litter Characteristics in Primiparous Sows.” Frontiers in Veterinary Science 10 (August 22, 2023). https://doi.org/10.3389/fvets.2023.1219572.
Li, Yan, Guoru He, Daiwen Chen, Bing Yu, Jie Yu, Ping Zheng, Zhiqing Huang, et al. “Supplementing Daidzein in Diets Improves the Reproductive Performance, Endocrine Hormones and Antioxidant Capacity of Multiparous Sows.” Animal Nutrition 7, no. 4 (December 2021): 1052–60. https://doi.org/10.1016/j.aninu.2021.09.002.
Meng, Qingwei, Tao Guo, Gaoqiang Li, Shishuai Sun, Shiqi He, Baojing Cheng, Baoming Shi, and Anshan Shan. “Dietary Resveratrol Improves Antioxidant Status of Sows and Piglets and Regulates Antioxidant Gene Expression in Placenta by Keap1-Nrf2 Pathway and SIRT1.” Journal of Animal Science and Biotechnology 9, no. 1 (April 20, 2018). https://doi.org/10.1186/s40104-018-0248-y.
Santos Passos, Fabiolla Rocha, Heitor Gomes Araújo-Filho, Brenda Souza Monteiro, Saravanan Shanmugam, Adriano Antunes Araújo, Jackson Roberto Almeida, Parimelazhagan Thangaraj, Lucindo José Júnior, and Jullyana de Quintans. “Anti-Inflammatory and Modulatory Effects of Steroidal Saponins and Sapogenins on Cytokines: A Review of Pre-Clinical Research.” Phytomedicine 96 (February 2022): 153842. https://doi.org/10.1016/j.phymed.2021.153842.
Serhan, C N, and N Chiang. “Endogenous Pro‐resolving and Anti‐inflammatory Lipid Mediators: A New Pharmacologic Genus.” British Journal of Pharmacology 153, no. S1 (March 2008). https://doi.org/10.1038/sj.bjp.0707489.
Sun, Jia‐Jie, Peng Wang, Guo‐Ping Chen, Jun‐Yi Luo, Qian‐Yun Xi, Geng‐Yuan Cai, Jia‐Han Wu, et al. “Effect of Moringa Oleifera Supplementation on Productive Performance, Colostrum Composition and Serum Biochemical Indexes of Sow.” Journal of Animal Physiology and Animal Nutrition 104, no. 1 (October 30, 2019): 291–99. https://doi.org/10.1111/jpn.13224.
Szczubiał, M. “Changes in Oxidative Stress Markers in Plasma of Sows during Periparturient Period.” Polish Journal of Veterinary Sciences, March 3, 2020, 185–90. https://doi.org/10.24425/pjvs.2020.132764.
Xu, Shengyu, Xiaojun Jiang, Xinlin Jia, Xuemei Jiang, Lianqiang Che, Yan Lin, Yong Zhuo, et al. “Silymarin Modulates Microbiota in the Gut to Improve the Health of Sow from Late Gestation to Lactation.” Animals 12, no. 17 (August 26, 2022): 2202. https://doi.org/10.3390/ani12172202.
Yang, Xizi, Ruizhi Hu, Mingkun Shi, Long Wang, Jiahao Yan, Jiatai Gong, Qianjin Zhang, Jianhua He, and Shusong Wu. “Placental Malfunction, Fetal Survival and Development Caused by Sow Metabolic Disorder: The Impact of Maternal Oxidative Stress.” Antioxidants 12, no. 2 (February 2, 2023): 360. https://doi.org/10.3390/antiox12020360.